分子标记技术的类型及其原理
- 格式:doc
- 大小:37.50 KB
- 文档页数:5

分子标记技术原理、方法及应用一、遗传标记的类型及发展遗传标记(genetic marker):指可追踪染色体、染色体某一节段、某个基因座在家系中传递的任何一种遗传特性.它具有两个基本特征,即可遗传性和可识别性;因此生物的任何有差异表型的基因突变型均可作为遗传标记。
包括形态学标记、细胞学标记、生化标记和分子标记四种类型。
形态学标记:主要包括肉眼可见的外部形态特征,如:矮秆、紫鞘、卷叶等;也包括色素、生理特性、生殖特性、抗病虫性等有关的一些特性。
优点:形态学标记简单直观、经济方便.缺点:(1)数量在多数植物中是很有限的; (2)多态性较差,表现易受环境影响; (3)有一些标记与不良性状连锁;(4)形态标记的获得需要通过诱变、分离纯合的过程,周期较长细胞学标记:植物细胞染色体的变异:包括染色体核型(染色体数目、结构、随体有无、着丝粒位置等)和带型(C带、N带、G带等)的变化。
优点:能进行一些重要基因的染色体或染色体区域定位。
缺点: (1)材料需要花费较大的人力和较长时间来培育,难度很大;(2) 有些变异难以用细胞学方法进行检测生化标记:主要包括同工酶和等位酶标记。
分析方法是从组织蛋白粗提物中通过电泳和组织化学染色法将酶的多种形式转变成肉眼可辩的酶谱带型.优点: 直接反映了基因产物差异,受环境影响较小.缺点:(1)目前可使用的生化标记数量还相当有限;(2)有些酶的染色方法和电泳技术有一定难度分子标记:主要指能反映生物个体或种群间基因组中某种差异特征的DNA片段,它直接反映基因组DNA间的差异,也叫DNA标记。
(1)数量多,高多态性,信息量大(2)与生长发育无关,取材不受限制(3)能明确辨别等位基因(4)均匀分布于整个基因组(5)选择中性,不影响目标性状的表达(6)检测手段简单、快速(7)成本低廉(8)稳定,重复性好(9)共显性遗传在遗传学研究中广泛应用的DNA分子标记已经发展了很多种,一般依其所用的分子生物学技术大致可以分为三大类:第一类是以分子杂交为核心的分子标记,包括RFLP、DNA指纹技术等,这类分子标记被称为第一代分子标记;第二类是以PCR为核心的分子标记,包括随机扩增多态性RAPD、简单序列重复SSR、扩增片段长度多态性AFLP、序列标签位点STS等,为第二代分子标记;第三类是一些新型的分子标记,如:SNP标记、表达序列标签EST 标记等,也以PCR技术为基础,为第三代分子标记。

分子标记辅助育种技术分子标记辅助育种技术第一节分子标记的类型和作用原理遗传标记是指可以明确反映遗传多态性的生物特征。
在经典遗传学中,遗传多态性是指等位基因的变异。
在现代遗传学中,遗传多态性是指基因组中任何座位上的相对差异。
在遗传学研究中,遗传标记主要应用于连锁分析、基因定位、遗传作图及基因转移等。
在作物育种中,通常将与育种目标性状紧密连锁的遗传标记用来对目标性状进行追踪选择。
在现代分子育种研究中,遗传标记主要用来进行基因定位和辅助选择。
1、形态标记形态标记是指那些能够明确显示遗传多态性的外观性状。
如、株高、穗型、粒色等的相对差异。
形态标记数量少,可鉴别标记基因有限,难以建立饱和的遗传图谱。
有些形态标记受环境的影响,使之在育种的应用中受到限制。
2、细胞学标记细胞学标记是指能够明确显示遗传多态性的细胞学特征。
如染色体的结构特征和数量特征。
核型:染色体的长度、着丝粒位置、随体有无。
可以反映染色体的缺失、重复、倒位、易位。
染色体结构特征带型:染色体经特殊染色显带后,带的颜色深浅、宽窄和位置顺序,可以反映染色体上常染色质和异染色质的分布差异。
染色体数量特征—是指细胞中染色体数目的多少。
染色体数量上的遗传多态性包括整倍体和非整倍体变异。
细胞学标记优点:克服了形态标记易受环境影响的缺点。
缺点:(1)培养这种标记材料需花费大量的人力物力;(2)有些物种对对染色体结构和数目变异的耐受性差,难以获得相应的标记材料;(3)这种标记常常伴有对生物有害的表型效应;(4)观察鉴定比较困难。
3、蛋白质标记用作遗传标记的蛋白质分为酶蛋白质和非酶蛋白质两种。
非酶蛋白质:用种子储藏蛋白质经一维或二维聚丙烯酰胺凝胶电泳,根据显示的蛋白质谱带或点,确定其分子结构和组成的差异。
酶蛋白质:利用非变性淀粉凝胶或聚丙烯酰胺凝胶电泳及特异性染色检测,根据电泳谱带的不同来显示酶蛋白在遗传上的多态性。
蛋白质标记的不足之处:(1)每一种同工酶标记都需特殊的显色方法和技术;(2)某些酶的活性具有发育和组织特异性;(3)标记的数量有限。


DNA分子标记技术概述1. 引言DNA分子标记技术是现代生物学和医学领域中非常重要的一项技术。
它可以通过特定的标记方法,在DNA分子上进行特异性地标记,从而实现对DNA序列的检测、定位和分析。
本文将对DNA分子标记技术进行全面、详细、完整和深入地探讨。
2. DNA分子标记技术的原理2.1 标记物选择在进行DNA分子标记之前,需要选择合适的标记物。
常用的DNA分子标记物包括荧光染料、辣根过氧化物酶标记物、生物素标记物等。
这些标记物具有不同的优势和适用范围,可以根据具体实验需求来选择合适的标记物。
2.2 标记方法DNA分子标记方法有多种,常用的包括直接标记法和间接标记法。
直接标记法是将标记物直接连接到DNA分子上,常用于荧光标记。
间接标记法是通过先引入标记物、再进行特定的反应来实现标记,常用于酶标记和生物素标记等。
2.3 标记效率和准确性DNA分子标记技术的效率和准确性是衡量其优劣的重要指标。
高效率和准确性可以保证实验结果的可靠性和准确性。
因此,在选择标记物和标记方法时,需要考虑到其标记效率和准确性,以及对实验结果的影响。
3. DNA分子标记技术的应用领域3.1 DNA测序和基因组学研究DNA分子标记技术在DNA测序和基因组学研究中有广泛的应用。
通过标记技术,可以对DNA序列进行检测和定位,从而实现对基因组的研究和分析。
3.2 分子诊断和疾病检测DNA分子标记技术在分子诊断和疾病检测中起到关键作用。
通过标记技术,可以检测和分析与疾病相关的基因或基因突变,从而实现早期诊断和治疗。
3.3 人类遗传学研究DNA分子标记技术对人类遗传学研究具有重要意义。
通过标记技术,可以进行人类遗传多样性和遗传变异的研究,为疾病发生机制和个体差异提供重要的参考和依据。
3.4 动植物遗传改良DNA分子标记技术在动植物遗传改良中有广泛应用。
通过标记技术,可以进行动植物基因分型和基因定位,为遗传改良工作提供重要的科学依据和技术支持。

分子标记1.分子标记技术及其定义1974年,Grozdicker等人在鉴定温度敏感表型的腺病毒DNA突变体时, 利用限制性内切酶酶解后得到的DNA片段的差异, 首创了DNA分子标记。
所谓分子标记是根据基因组DNA存在丰富的多态性而发展起来的可直接反映生物个体在DNA水平上的差异的一类新型的遗传标记,它是继形态学标记、细胞学标记、生化标记之后最为可靠的遗传标记技术。
广义的分子标记是指可遗传的并可检测的DNA序列或蛋白质分子。
通常所说的分子标记是指以DNA多态性为基础的遗传标记。
分子标记技术本质上都是以检测生物个体在基因或基因型上所产生的变异来反映基因组之间差异。
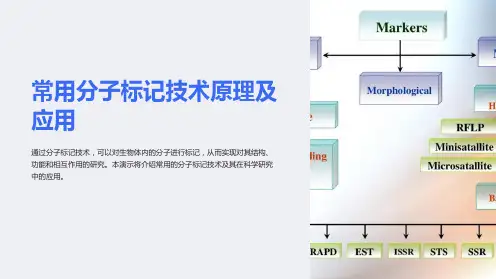
2.分子标记技术的类型分子标记从它诞生之日起, 就引起了生物科学家极大的兴趣,在经历了短短几十年的迅猛发展后, 分子标记技术日趋成熟, 现已出现的分子标记技术有几十种, 部分分子标记技术所属类型如下。
2.1 建立在Southern杂交基础上的分子标记技术(1) RFLP ( Rest rict ion Fragment Length Polymorphism)限制性内切酶片段长度多态性标记;(2) CISH ( Chromosome In Situ Hybridization) 染色体原位杂交。
2.2 以重复序列为基础的分子标记技术(1) ( Satellite DNA ) 卫星DNA;(2) ( Minisatellite DNA ) 小卫星DNA;(3) SSR( Simple Sequence Repeat ) 简单序列重复, 即微卫星DNA。
2.3 以PCR为基础的分子标记技术(1) RAPD ( Randomly Amplif ied Polymorphic DNA ) 随机扩增多态性DNA;(2) AFLP( Amplif ied Fragment Length Polymorphism) 扩增片段长度多态性;(3) SSCP( Single Strand Conformation Polymorphism) 单链构象多态性;(4) cDNA-AFLP( cDNA- AmplifiedFragment Length Polymorphism) cDNA -扩增片段长度多态性;(5) TRAP( Target Region Amplified Polymorphism) 靶位区域扩增多态性;(6) SCAR ( Sequence Char acterized Amplified Region) 序列特征化扩增区域;(7) SRAP ( Sequencerelated Amplified Polymorphism) 相关序列扩增多态性。

分子标记种类及概述分子标记是一种在生物学、生物化学和药理学研究中广泛应用的技术。
它主要通过将分子或化合物与特定的标记物相结合,以便于对其进行检测、跟踪和定量分析。
分子标记的种类非常多样,包括荧光标记、放射性标记、酶标记和生物素标记等。
每种标记方法都有其特定的优势和适用范围,下面将详细介绍这些分子标记的类型及其概述。
1.荧光标记:荧光标记是最常用且广泛应用的一种分子标记方法。
它通过将目标分子与荧光染料结合,利用目标分子与激发光源相互作用后发出荧光信号来进行检测和定量分析。
荧光标记具有灵敏度高、非破坏性、实时监测能力强等特点,适用于细胞生物学、分子遗传学和生物化学等研究领域。
2.放射性标记:放射性标记是利用放射性同位素来标记目标分子的一种方法。
通过将放射性同位素(如3H、14C、32P等)与目标分子结合,可以通过放射性衰变的特性来检测和定量分析目标分子。
放射性标记具有极高的敏感性和特异性,适用于分子生物学、药理学和临床药理学等研究领域。
3.酶标记:酶标记是利用酶来标记目标分子的一种方法。
通过将酶与目标分子结合,然后加入适当的底物来触发酶的催化反应,可以产生可见色素或荧光信号,从而实现对目标分子的检测和定量分析。
酶标记具有高度特异性和灵敏度,适用于生物化学、免疫学和临床检验等研究领域。
4.生物素标记:生物素标记是利用生物素(一种小分子)与目标分子结合,然后利用亲和性层析或荧光染料来检测和定量分析目标分子的一种方法。
生物素标记具有快速、简单和高效的特点,适用于生化学、药理学和分子生物学等研究领域。
除了以上几种常见的分子标记方法外,还有许多其他的分子标记方法,比如金纳米颗粒标记、蛋白质标记和DNA标记等。
这些标记方法可以根据研究的具体需求来选择和应用。
标记方法的选择应考虑到目标分子的性质、研究目的和实验条件等因素。
分子标记在生物学研究中有着广泛的应用,如细胞成像、蛋白质定位、基因表达研究等。
它们在分子和细胞水平上为我们提供了许多有关生物学过程和分子机制的信息。

遗传学中的分子标记技术遗传学是研究遗传现象的一门学科,而分子标记技术则是其中的一个重要领域。
它不仅可以帮助我们研究物种间的遗传联系,还可以应用于医学和农业领域,为人们的生活带来更多便利和进步。
本文将介绍遗传学中的分子标记技术,探讨其在实践中的应用以及未来的发展方向。
一、分子标记技术简介分子标记技术是利用分子水平的遗传标记对个体、品系或群体进行鉴别、分类、分子配对等分析的一种技术。
目前常用的几种分子标记技术包括限制性片段长度多态性(RFLP)、随机扩增多态性(RAPD)、序列标记位点(SSR)和单核苷酸多态性(SNP)等。
RFLP技术是一种基于DNA序列限制性切割位点的分析方法。
通过将基因组DNA切成不同的长度片段,然后对这些片段进行电泳分离,最后通过DNA探针的帮助确定特定位点的DNA序列。
RAPD技术则是一种无需事先知道DNA序列的技术,通过使用随机序列的寡核苷酸为引物进行PCR扩增,经过电泳分离后可以得到特定长度的DNA条带。
SSR技术则是利用序列中重复核苷酸序列的多态性,选取特定的序列扩增后进行电泳分离,得到条带后可以确定所研究物种基因组的遗传变异情况。
SNP技术则是一种最新的分子标记技术,它是基于单核苷酸变异位点的方法,通过测量单个碱基的点突变来分析遗传多样性。
二、分子标记技术的应用1.遗传分析分子标记技术在遗传学研究中可以用于基因型鉴定、亲缘关系分析、遗传多样性评估等方面。
例如,利用SSR技术可以分析豆科作物的遗传多样性,帮助育种学家定位有用的基因,并加速豆科作物的育种进程。
另外,RFLP技术还可以用于协助医学领域的DNA指纹分析,对于识别罪犯身份、证明亲子关系等方面都有巨大贡献。
2.病理学研究在病理学研究中,分子标记技术可以用于检测各种疾病的基因突变、表达谱的差异、重要调节基因的变化等。
例如,SNP技术可以用于筛查患有代谢性疾病的患者,SSR技术可以用于评价肿瘤的恶性程度。
3.农业领域分子标记技术在农业领域中的应用越来越普遍,可以用于作物品种鉴别、繁殖方式分析、作物改良等方面。

分子标记技术的种类根据不同的核心技术基础,DNA分子标记技术大致可分为三类: 第一类以Southern杂交为核心, 其代表性技术为RFLP;第二类以PCR技术为核心,如RAPD、SSR、AFLP、STS、SRAP、TRAP等;第三类以DNA序列(mRNA或单核苷酸多态性)为核心,其代表性技术为EST标记、SNP标记等。
理想的分子标记应达到以下的要求:①具有高的多态性;②共显性遗传;③能够明确辨别等位基因;④分布于整个基因组中;⑤选择中性(即无基因多效性);⑥检测手段简单、快速;⑦开发成本和使用成本尽量低廉;⑧在实验室内和实验室间重复性好。
目前,没有任何一种分子标记均满足以上的要求,它们均具有各自的优点和不足。
其特点比较见表一。
1限制性内切酶片段长度多态性标记(Restriction Fragment Length Polymorphism,RFLP)1974年,Grozdicker 等人鉴定温度敏感表型的腺病毒DNA突变体时,发现了经限制性内切酶酶解后得到的DNA片段产生了差异,由此首创了第一代DNA分子标记技术——限制性内切酶片段长度多态性标记(RFLP)。
其原理是由于不同个体基因型中内切酶位点序列不同(可能由碱基插入、缺失、重组或突变等造成),利用限制性内切酶酶解基因组DNA时,会产生长度不同的DNA酶切片段,通过凝胶电泳将DNA片段按各自的长度分开,通过Southern印迹法,将这些大小不同的DNA片段转移到硝酸纤维膜或尼龙膜上,再用经同位素或地高辛标记的探针与膜上的酶切片段分子杂交,最后通过放射性自显影显示杂交带,即检出限制性片段长度多态性。
进行RFLP时,酶切要彻底,注意内切酶的选择,对于亲缘关系很近的物种,可增加内切酶的使用种类。
目前RFLP的使用领域很广泛,其具有以下优点:①RFLP标记源于基因组DNA的自身变异,理论上可覆盖整个基因组,能提供丰富的遗传信息;②标记不受组织、环境和发育阶段的影响;③呈共显性,即杂交时等位DNA片段均呈现带,能区分纯合基因型和杂合基因型,F2表现出 1∶2∶1的孟德尔分离定律[3],提供标记座位完全的遗传信息;④由于限制性内切酶的专一性使结果稳定可靠,重复性好。

分子标记原理和技术分子标记原理和技术是一种用于研究和检测生物分子的方法。
分子标记是通过给生物分子附上一种特定的标记物,使其能够被观察和测量。
分子标记技术在生物医学研究、临床诊断、药物研发和环境监测等领域都有广泛的应用。
分子标记的原理是利用化学反应将标记物与待检测的生物分子结合起来,然后通过适当的方法观察或检测标记物。
常见的标记物有荧光染料、放射性同位素、酶和金纳米粒子等。
标记物的选择要考虑其化学性质、稳定性、检测灵敏度和特异性等因素。
分子标记技术有很多种,下面列举几种常见的技术:1.荧光标记:荧光标记是最常用的分子标记技术之一、通过给生物分子附加荧光染料,可以通过荧光显微镜观察其分布和表达水平。
荧光标记还可以用于流式细胞术、酶联免疫吸附实验等。
荧光标记可以选择多种不同的荧光染料,如草莓红、FITC和PE等。
2.放射性标记:放射性标记是利用放射性同位素将标记物与生物分子结合起来。
这种标记方法可以通过放射性计数器或放射影像技术来检测,具有极高的灵敏度。
常用的放射性同位素有3H(氚)、14C(碳14)和32P(磷32)等。
3.酶标记:酶标记是利用酶与底物之间的反应来检测生物分子。
常用的酶有辣根过氧化物酶(HRP)和碱性磷酸酶(AP)。
酶标记技术可以通过底物的颜色变化或荧光信号来观察酶的活性和分布。
4. 化学标记:化学标记是利用特定化学反应将标记物与生物分子结合起来。
常见的化学标记方法有SNAP标记、CLIP标记和Biotin-avidin 标记等。
化学标记的优点是反应选择性高,标记物的稳定性和特异性好。
5.金纳米粒子标记:金纳米粒子标记是一种新兴的分子标记技术。
金纳米粒子可以通过调节粒子大小和表面修饰来实现对生物分子的特异性识别。
金纳米粒子标记可以通过紫外-可见吸收光谱或扫描电镜观察。
分子标记技术在生物学研究中扮演着重要角色,能够帮助科学家观察和分析生物分子的功能和相互作用。
此外,分子标记技术还被广泛应用于临床诊断和药物研发领域,例如用于检测肿瘤标记物、鉴定药物靶点和筛选药物库。

分子标记技术的类型原理及应用分子标记技术是一种基于分子生物学的技术,在研究、诊断和治疗等领域具有广泛的应用价值。
这种技术利用染料、荧光物质、辐射标记物等来标记目标分子,从而实现对分子的检测、追踪和研究。
下面将介绍分子标记技术的几种类型、原理及应用。
一、荧光标记技术荧光标记技术是一种常见的分子标记技术,基于物质的荧光特性,通过在目标分子上标记荧光染料或荧光蛋白等物质,实现对目标分子的可见或可荧光检测。
该技术的原理是标记物被激发后会发出荧光,通过检测荧光信号的强度、波长或寿命等特征来获得关于目标分子的信息。
荧光标记技术在生物学研究、生命体内药物输送系统的研究和临床诊断等方面得到了广泛的应用。
在生物学研究中,荧光标记技术可以用于研究细胞结构和功能、蛋白质相互作用、细胞内信号传导等。
在药物输送系统的研究中,荧光标记技术可以用于研究药物在体内的分布和代谢情况等。
在临床诊断中,荧光标记技术可以用于检测血液中的病原体、肿瘤标志物以及其他疾病相关分子等。
二、辐射标记技术辐射标记技术是一种通过辐射标记物对目标分子进行标记的技术。
常用的辐射标记物包括放射性同位素和放射性荧光染料等。
该技术的原理是通过辐射标记物自身所放出的辐射(如α、β射线等)或荧光来检测目标分子。
辐射标记技术在医学、生物学和环境科学等领域都有广泛的应用。
在医学方面,辐射标记技术可以用于肿瘤的早期诊断和治疗、药物代谢和排泄的研究等。
在生物学方面,辐射标记技术可以用于研究生物体的代谢过程、病原体的传播途径等。
在环境科学方面,辐射标记技术可以用于了解污染物的迁移和转化、生态系统的功能及稳定性等。
三、化学标记技术化学标记技术是一种通过化学反应将标记物与目标分子结合的技术。
常见的化学标记物包括生物素、抗原抗体等。
该技术的原理是通过物质间的化学反应使两者结合,并通过检测化学标记物的特征来获得目标分子的信息。
化学标记技术在生物医学研究、食品安全检测和环境监测等领域有广泛应用。
分子标记的种类及其发展分子标记是指通过将特定的标记分子连接到目标分子上来实现对其进行检测和定位的技术。
随着生物学、医学和化学等领域的发展,分子标记逐渐得到了广泛应用。
目前,分子标记主要分为以下几类:荧光标记、放射性标记、酶标记和纳米颗粒标记。
以下是对每种标记的简要介绍以及其发展情况。
1.荧光标记:荧光标记是一种通过激发分子到高能态,再通过发射光子使其降至低能态的方式来实现分子标记的方法。
荧光标记的特点是对目标物质的检测灵敏、成本相对较低、操作相对简便等。
随着成像技术的发展,荧光标记已经广泛应用于细胞和分子生物学、医学影像等领域。
2.放射性标记:放射性标记是通过标记分子与放射性同位素结合来实现分子标记的方法。
放射性标记的特点是检测敏感度高、能够实现定量测量等。
然而,由于放射性同位素的辐射危害,放射性标记的应用受到了限制。
近年来,随着技术的发展,放射性标记已经逐渐向非放射性标记转变,例如利用放射性同位素替代品或荧光物质进行标记。
3.酶标记:酶标记是将酶与目标分子结合,然后利用酶的催化活性对目标分子进行检测的方法。
酶标记的特点是灵敏度高、选择性好、对目标分子的检测快速等。
常见的酶标记方法包括辣根过氧化物酶(HRP)标记、碱性磷酸酶(AP)标记等。
酶标记在生物学和生物化学研究中广泛应用,特别是在免疫学研究和生物医学检测中得到了广泛应用。
4.纳米颗粒标记:纳米颗粒标记是将纳米颗粒与目标分子结合,然后利用纳米颗粒的特殊性质对目标分子进行检测的方法。
纳米颗粒标记的特点是灵敏度高、标记稳定性好、可实现多重标记等。
常见的纳米颗粒包括金纳米颗粒、磁性纳米颗粒等。
近年来,纳米颗粒标记在生物医学检测、分子诊断等领域得到了广泛应用,并展现出了很大的潜力。
总体来说,分子标记的发展已经取得了很大的进展,不同种类的标记各具特点,适用于不同领域的研究和应用。
未来,随着技术的不断发展,分子标记将更加精准、灵敏和多样化,为科学研究、医学诊断和治疗等领域提供更多的可能。
分子标记原理和技术分子标记是一种用于追踪和分析生物分子的技术,在生命科学研究中得到广泛应用。
它通过在特定分子上加上标记物,如荧光染料、放射性同位素或酶等,来实现对这些分子的检测和定位。
分子标记技术的原理主要包括标记物的选择和绑定、信号的检测和分析等方面。
分子标记技术的核心是选择适合的标记物,并将其与目标分子进行特异性的结合。
常用的标记物包括荧光染料、放射性同位素和酶等。
荧光染料是一类可发光的化学物质,可以通过荧光显微镜来检测其存在和分布情况。
放射性同位素则利用放射性衰变的原理,通过放射性测量仪来检测其辐射信号。
酶则是一类能够催化特定化学反应的蛋白质,通过对酶反应进行检测来间接地确定目标分子的存在。
标记物与目标分子的结合方式多种多样,常用的方法包括共价结合、亲和结合和非共价结合等。
共价结合是指通过化学反应将标记物与目标分子共同连接起来,常用的反应有偶氮化反应、醛基化反应等。
亲和结合则是利用亲和分子对目标分子进行特异性结合,常用的亲和分子包括抗体、配体等。
非共价结合则是通过分子间的非共价相互作用来实现标记物与目标分子的结合,如疏水相互作用、静电相互作用等。
分子标记技术中信号的检测和分析是至关重要的一步。
对于荧光标记物,需要使用荧光显微镜来观察目标分子的荧光信号,并通过图像处理和分析来定量分析目标分子的数量和分布情况。
对于放射性同位素标记物,需要使用放射性测量仪来测量目标分子的放射性信号,并通过计数方法来确定目标分子的数量。
对于酶标记物,需要使用酶反应底物来触发酶催化反应,产生可测量的信号,如颜色变化、发光等,通过光谱仪或分光光度计来检测和分析。
分子标记技术具有许多优点,如高灵敏度、高特异性、高分辨率等。
它可以用于检测和定位生物分子,如蛋白质、核酸等,也可以用于研究生物分子的相互作用、代谢途径等。
分子标记技术在生命科学研究中的应用非常广泛,如免疫组织化学、原位杂交、蛋白质定位、细胞追踪等。
2. Saha K, Agasti SS, Kim C, Li X, Rotello VM. Gold nanoparticles in chemical and biological sensing. Chem Rev. 2024;112(5):2739-2779.。
分子标记技术的类型及其原理08农生1班陈耀光 200830010403所谓分子标记就是基于基因组DNA 存在极其丰富的多态性而发展的一类可以直接反映生物个体间DNA 水平上差异的新型的遗传标记方法。
在遗传学发展过程中,先后出现了形态学标记、细胞学标记、生化标记和分子标记,其中以分子标记最为理想、可靠,因为DNA分子中碱基的缺失、插入、易位、倒位或是长短与排列不一的重复序列等产生的差异,都可以通过分子标记进行检测。
DNA 分子标记较以往的形态标记其优越性表现在:(1)以核酸为研究对象,不受季节、环境限制,不存在基因表达与否的问题,也没有组织或器官特异性;(2)数量的丰富性,遍及整个基因组,标记的数量几乎是无限的;(3)多态性高,自然存在丰富的等位变异;(4)许多标记表现为共显性,能很好地鉴别纯合基因型与杂合基因型;(5)检测手段简便、快速,并且重复性好;(6)既不对目标形状的表达造成影响,也不会与不良性状之间产生必然的关联。
1 分子标记的类型及其原理分子标记技术自诞生以来,短短的几十年时间中得到突飞猛进的发展,至今被发展和利用的分子标记技术已有二十余种,为不同研究领域提供了有效的技术手段,同时也发挥着至关重要的作用。
目前,根据对DNA 多态性检测手段和所应用序列范围的不同,对部分分子标记技术分类如下。
1.1 基于全基因序列的分子标记RFLP (restriction fragment length polymorphism,限制性片段长度多态性):RFLP 作为最早发展的分子标记技术由Grozdicker 等于1974 年创建,并由Bostein 等再次提出。
RFLP 技术的出现开创了直接在DNA 水平上进行遗传研究的新时代。
其基本原理是:基因组DNA中限制性内切酶所识别的序列由于出现碱基变化而致使酶切位点的数量也变化,从而使酶切片段长短发生差异产生长度多态性。
利用特定的限制性内切酶切割不同个体的基因组DNA,由于不同个体中酶切位点的差别就得到了长短相异的片段DNA,电泳分离后,借助Southern 杂交将DNA 片段转移至硝酸纤维素膜上,将具有放射性标记的探针与膜上的片段杂交,通过放射自显影技术就可以获得显示物种特异性的多态性图谱。
常用分子标记技术原理及应用分子标记技术是现代分子生物学、生物化学和生物医学研究中常用的重要方法之一,其原理是利用特定的物质(分子标记)与待检测分子结合,从而实现对待检测分子的定位、测定和分析。
常用的分子标记技术包括荧光标记、酶联免疫法(ELISA)、放射性同位素标记和生物素标记等,下面将详细介绍其中的原理及应用。
1.荧光标记技术荧光标记技术是一种基于物质固有性质的分子标记方法,其原理是将待检测物质与荧光染料结合,通过荧光信号的激发和发射实现对物质的定位和检测。
荧光标记技术具有高灵敏度、多重标记、高分辨率和实时监测等优点,在生物学研究和临床诊断中得到广泛应用。
例如,荧光标记技术可应用于细胞内分子定位、蛋白质相互作用研究和病原体检测等领域。
2.酶联免疫法(ELISA)酶联免疫法是一种常用的免疫学实验方法,其原理是将待检测物质与特异性抗体结合,然后再用酶标记的二抗对抗体进行反应,通过酶底物的转化反应实现对待检测物质的定性和定量分析。
酶联免疫法具有高灵敏度、高特异性和简单易行等特点,在医学诊断和生物分析中被广泛应用。
例如,酶联免疫法可用于检测临床血清中的肿瘤标志物、抗体和炎症因子等,对于早期疾病诊断、药物研发和治疗效果评估具有重要意义。
3.放射性同位素标记技术放射性同位素标记技术是一种基于放射性元素的分子标记方法,其原理是将待检测物质与放射性同位素结合,通过放射性同位素的放射衰变实现对物质的定位和追踪。
放射性同位素标记技术具有极高的灵敏度和追踪性,广泛应用于核医学、分子显像和生物研究等领域。
例如,放射性同位素标记技术可用于肿瘤显像、药物代谢研究和放射免疫测定等,对于肿瘤早期诊断、药物研发和治疗效果评估有着重要的作用。
4.生物素标记技术生物素标记技术是一种基于生物素-亲和素相互作用的分子标记方法,其原理是将待检测物质与生物素结合,通过生物素和亲和素之间的特异性结合实现对物质的定位和检测。
生物素标记技术具有高特异性、高亲和力和多重标记等优势,在生物学研究和生物医学中得到广泛应用。
分子标记的原理特点及应用一、分子标记的原理分子标记是一种用于确定和定位特定分子的方法。
它基于分子中的特定结构或性质进行标记,并利用这些标记来进行分子的定位和识别。
常见的分子标记方法包括荧光标记、抗体标记和DNA标记等。
1. 荧光标记荧光标记是通过给分子引入荧光物质,使其发出特定波长的荧光信号来标记分子。
荧光标记的原理是将某种荧光物质或染料与目标分子结合,形成带有荧光信号的复合物。
荧光标记具有灵敏度高、实时性好等优点,广泛应用于药物研发、细胞生物学等领域。
2. 抗体标记抗体标记是通过给目标分子结合抗体,利用抗体的特异性和亲和性来对目标分子进行标记。
抗体标记的原理是将目标分子与特定的抗体结合,形成结合复合物。
抗体标记具有高度特异性和灵敏性,常用于生命科学研究和临床诊断等领域。
3. DNA标记DNA标记是利用DNA分子的特性对分子进行标记和检测。
DNA标记的原理是将目标分子与特定的DNA序列结合,形成带有DNA标记的复合物。
DNA标记常用于基因测序、分子诊断和基因工程等领域。
二、分子标记的特点分子标记具有以下几个特点:1. 高选择性分子标记的方法通常具有高度的选择性,可以根据目标分子的特定结构或性质进行标记。
这使得分子标记可以精确地定位和识别目标分子,减少误判和混淆。
2. 高灵敏度分子标记方法通常具有高度的灵敏度,可以检测到非常低浓度的目标分子。
这使得分子标记成为很多科学研究和临床诊断中的重要工具,例如用于检测罕见疾病的基因突变。
3. 实时性分子标记方法通常具有较快的响应速度和实时性,可以实时监测和观察目标分子的变化。
这使得分子标记在动态过程观察和实时监测中具有重要意义,例如用于研究细胞信号转导的过程。
4. 多样性分子标记具有多样性,可以根据具体需求选择不同的标记方法和标记物质。
不同的标记方法可以针对不同的分子结构和目标分子进行标记,满足不同领域和研究的需求。
三、分子标记的应用分子标记在多个领域中得到广泛应用,包括生命科学研究、临床诊断、药物研发等。
分子标记详细教程分子标记是一种广泛应用于生物学和生物化学研究中的技术,它能够帮助科学家们定位、观察和分析分子的位置和功能。
在这个教程中,我们将详细介绍分子标记的原理和方法,并讲解如何正确地进行分子标记实验。
一、分子标记的原理分子标记是利用特定的化学物质(标记物)对目标分子进行标记,从而使其在实验中能够被观察和检测到。
常用的分子标记方法包括荧光标记、放射性标记和酶标记等。
这些方法都基于不同的原理,但最终目的都是通过标记物的特性来实现对目标分子的检测和定位。
二、分子标记的方法1.荧光标记:荧光标记是最常用的分子标记方法之一。
它利用具有荧光特性的染料或荧光蛋白对目标分子进行标记,然后利用荧光显微镜等设备观察荧光信号。
这种方法可以实现对分子位置、形态和数量的检测。
2.放射性标记:放射性标记是利用放射性同位素对目标分子进行标记。
通过测量目标分子放射出的射线,可以得到目标分子的定量和定位信息。
这种方法在一些需要高灵敏度和高分辨率的实验中被广泛应用。
3.酶标记:酶标记是利用酶对目标分子进行标记。
酶标记的原理是将酶与目标分子结合,然后通过酶的催化作用来检测目标分子的存在和活性。
常用的酶标记方法包括辣根过氧化物酶(HRP)标记和碱性磷酸酶(AP)标记等。
三、分子标记实验步骤1.选择适合的标记物:根据实验需求和目标分子的特性,选择合适的标记物进行标记。
常用的标记物包括荧光染料、放射性同位素和酶等。
2.标记物与目标分子的结合:将标记物与目标分子进行反应,使它们发生特异性结合。
这一步骤需要注意反应条件的控制,以确保标记物与目标分子的结合效率和特异性。
3.纯化标记产物:通过一系列的纯化步骤,将标记物和未反应的物质进行分离,得到纯净的标记产物。
纯化的方法可以根据标记物的特性和实验需求选择。
4.标记物的检测和定位:利用相应的检测方法对标记物进行检测和定位。
具体的方法可以根据标记物的特性和实验需求选择,如荧光显微镜、放射性计数器和酶标仪等。
分子标记技术的类型及其原理08农生1班陈耀光 200830010403所谓分子标记就是基于基因组DNA 存在极其丰富的多态性而发展的一类可以直接反映生物个体间DNA 水平上差异的新型的遗传标记方法。
在遗传学发展过程中,先后出现了形态学标记、细胞学标记、生化标记和分子标记,其中以分子标记最为理想、可靠,因为DNA分子中碱基的缺失、插入、易位、倒位或是长短与排列不一的重复序列等产生的差异,都可以通过分子标记进行检测。
DNA 分子标记较以往的形态标记其优越性表现在:(1)以核酸为研究对象,不受季节、环境限制,不存在基因表达与否的问题,也没有组织或器官特异性;(2)数量的丰富性,遍及整个基因组,标记的数量几乎是无限的;(3)多态性高,自然存在丰富的等位变异;(4)许多标记表现为共显性,能很好地鉴别纯合基因型与杂合基因型;(5)检测手段简便、快速,并且重复性好;(6)既不对目标形状的表达造成影响,也不会与不良性状之间产生必然的关联。
1 分子标记的类型及其原理分子标记技术自诞生以来,短短的几十年时间中得到突飞猛进的发展,至今被发展和利用的分子标记技术已有二十余种,为不同研究领域提供了有效的技术手段,同时也发挥着至关重要的作用。
目前,根据对DNA 多态性检测手段和所应用序列范围的不同,对部分分子标记技术分类如下。
1.1 基于全基因序列的分子标记RFLP (restriction fragment length polymorphism,限制性片段长度多态性):RFLP 作为最早发展的分子标记技术由Grozdicker 等于1974 年创建,并由Bostein 等再次提出。
RFLP 技术的出现开创了直接在DNA 水平上进行遗传研究的新时代。
其基本原理是:基因组DNA中限制性内切酶所识别的序列由于出现碱基变化而致使酶切位点的数量也变化,从而使酶切片段长短发生差异产生长度多态性。
利用特定的限制性内切酶切割不同个体的基因组DNA,由于不同个体中酶切位点的差别就得到了长短相异的片段DNA,电泳分离后,借助Southern 杂交将DNA 片段转移至硝酸纤维素膜上,将具有放射性标记的探针与膜上的片段杂交,通过放射自显影技术就可以获得显示物种特异性的多态性图谱。
RFLP 标记的等位基因是共显性的,能区分纯合基因型和杂合基因型,结果稳定可靠,重复性好,适合于连锁图谱的建立。
但是对DNA 要求较高,检测步骤多,操作复杂,耗时费资,而且需要放射性同位素等,这都在很大程度上限制了RFLP 技术的应用。
RAPD (randomly amplified polymorphic DNA,随机扩增多态性DNA):1990 年Williams 等(1990)与Welsh 和McClelland (1990)两个研究小组分别提出RAPD 标记技术,该技术是建立在PCR 基础上,对未知序列的全基因组进行多态性分析的一种新型分子标记技术。
其基本原理是利用一个人工合成的随机寡核苷酸(一般为8~10 bp)为引物,通过PCR对基因组DNA 进行体外扩增,产生大小不同的DNA 片段,再经凝胶电泳分析扩增产物呈现DNA 片段的多态性。
与RFLP 相比,RAPD 技术的优点是技术简单,检测迅速,安全性好,DNA 用量少,设计引物不需知道序列信息等,但RAPD 标记为显性遗传,不能区分纯合基因型与杂合基因型,且易受多种因素影响,结果的稳定性和重复性难以保证。
AFLP (amplification fragment length polymorphism,扩增片段长度多态性;又称选择性限制片段扩增:AFLP 技术是1993 年由荷兰科学家Zabeau等创建的一种将限制性酶切和PCR 技术相结合的检测DNA 多态性的分子标记方法。
其实质是对基因组DNA 限制性酶切片段进行选择性扩增,具体过程为基因组DNA 经两种或两种以上的限制性内切酶酶切,产生大小不等的随机片段,将人工双链接头连接到这些片段的末端,从而作为扩增反应的模板,再根据接头的序列和酶切位点设计引物实现选择性PCR 扩增。
AFLP 技术可以说是集RFLP 和RAPD 技术与一身,兼备两者的优点,既有RFLP 的可靠性,又具有RAPD 的高效性,且重复性好,是一种十分理想的遗传标记,但与此同时也给试验带来了昂贵的费用和复杂的操作过程,尽管如此,该技术依然在遗传多样性研究、遗传图谱的构建、基因定位和品质鉴定等方面已得到广泛的应用,经过人们近年来不断地改进和完善,AFPL 被认为是迄今为止最有效的分子标记。
1.2 基于简单重复序列的分子标记SSR (simple sequence repeat, 简单序列重复, 又称微卫星DNA):一般是指基因组中存在的由2~6 个核苷酸为重复单位组成的长达几十个核苷酸的串联重复序列。
这种序列广泛分布于真核生物基因组,含量丰富,且随机均匀分布,其重复次数的不同产生了等位基因之间的多态性。
微卫星由核心序列和两侧的保守侧翼序列构成,可以根据SSR 两侧的保守序列设计引物进行PCR 扩增,由于不同品种SSR 基序的重复次数不同,导致PCR 扩增条带差异性,即简单重复序列长度多态性(simple sequencelength polymorphism, SSLP)。
SSR 标记具有以下优点:表现多为共显性遗传,遵循孟德尔遗传法则;数量丰富,广泛分布于整个基因组;对DNA 数量及纯度要求不高,易于利用PCR 检测;实验操作简便,结果稳定,重复性强。
然而,SSR 引物具有高度的种属特异性,开发和合成新的SSR 引物费用高、难度大,但由于其优点众多,在遗传图谱构建、基因定位、品种鉴定、种质资源保存和利用等方面均有十分广泛的应用。
ISSR (inter-simple sequence repeat, 内部简单序列重复):ISSR 技术是对SSR 的进一步发展,利用SSR 本身设计引物,其基本原理就是在SSR 的3' 或5' 端锚定1~4 个随机选择的核苷酸,从而保证引物对两个相距较近、方向相反的SSR 之间的DNA 区段进行PCR 扩增,然后进行电泳、染色,观察谱带情况分析不同样品间的多态性,因此这类标记又被称为锚定简单序列重复(anchored simple sequence repeat,ASSR)或锚定微卫星引物PCR (anchored microsatellite-primed PCR)。
该技术具有引物设计简单,多态性较RFLP、SSR、RAPD 丰富,可重复性和稳定性好于RAPD 等特点。
SPAR (single primer amplification reaction, 单引物扩增反应):是与RAPD 技术相似的一种分子标记技术,SPAR 也只用一条引物,不同在于所用引物不是随机的,而是根据SSR 序列设计,通过PCR 扩增SSR 之间的序列,然后电泳分析其多态性。
另外,还有一种与SPAR 技术十分相近的标记技术,即ISTR(inverse sequence-tagged repeat, 反向序列标签重复技术),在反向重复序列基础上设计引物,进而扩增反向重复序列之间的DNA 序列。
1.3 基于已知的特定序列的分子标记STS (sequence tagged site, 序列标签位点):是指基因组上一段已知序列的、作为位置标志的单拷贝短DNA 片段,长度多在200~500 bp 之间。
这种序列通常在染色体上只出现一次,从而可以作为基因组中的特异位点,以此判断DNA 的方向和其它特定序列的相对位置。
理论上说,任何单拷贝序列,只要知道其在基因组中的位置,都可以转变为STS 标记。
依据单拷贝DNA 片段两端序列设计一对长度为20 bp左右的特异引物,进行PCR 扩增以显示STS 特异片段。
STS 标记的特点是操作简单,信息量大,多态性好,呈共显性标记,在人类基因组物理图谱作图中发挥着重要的作用。
EST (expressed sequence tag, 表达序列标签):EST 技术于1991 年由美国国立卫生研究院(NationalInstitutes of Health, NIH) 的生物学家Venter 首次提出。
EST 是指对cDNA 文库中随机挑选克隆,对其5'端或3' 端进行一步法测序后获得的长约150~500 bp的表达序列片段。
一个EST 可代表生物体某种组织某一时期的一个表达基因。
以EST 为基础的分子标记除具有一般分子标记的特点外,还有开发简便、信息量高和通用性好等优势。
EST 标记根据开发的方法不同可以分为4 类:(1) EST-PCR 和EST-SSR (微卫星)标记,是以PCR技术为核心,操作简便、经济,是目前研究和应用最多的一类;(2) EST-SNP (单核苷酸多态性)标记,是以特定EST 区段内单个核苷酸差异为基础的标记,可依托杂交、PCR 等多种手段进行检测;(3) EST-AFLP标记,是以限制性内切酶技术和PCR 相结合为基础的标记;(4) EST-RFLP 标记,是以限制性内切酶和分子杂交为依托,以EST 本身作为探针,与经过不同限制性内切酶消化后的基因组DNA 杂交而产生的。
1.4 基于反转录转座子的分子标记SSAP (sequence-specific amplification polymorphism,特异性序列扩增多态性):SSAP 是一种基于反转录转座子插入位点与邻近限制性内切酶位点之间的DNA 多态性,其基本步骤是将基因组DNA 酶切后,连接与酶相对应的接头,预扩增后利用基于长末端重复序列的特异性引物和加选择性碱基的酶切引物进行PCR 扩增。
SSAP 技术是目前应用最广泛的一种基于转座元件的分子标记,具有标记数量多、稳定性高等优点,但其结果受多种因素影响,难以保证结果的有效性和可靠性。
IRAP(inter-retrotransposon amplified polymorphism,逆转录转座子插入位点间扩增多态性):Kalendar等(1999)提出的一种能够检测逆转录转座子插入位点间多态性的分子标记。
原理是根据逆转录转座子的长末端重复序列中的保守序列设计引物,检测基因组中相邻的同家族逆转录转座子成员之间的片段,还可以根据逆转录酶基因中相对保守区域设计引物进行扩增。
REMAP(retrotransposon-microsatellite amplifiedpolymorphism, 逆转录转座子微卫星扩增多态性):REMAP 技术是一种基于反转录转座子的标记技术,基本原理是利用逆转录转座子的长末端重复序列中的保守序列与SSR 序列分别设计引物,以便扩增出逆转录转座子与相邻的SSR 间的片段,进而可以检测逆转录转座子插入位点上长末端重复序列与简单重复序列之间的多态性。