关于动物冷应激
- 格式:doc
- 大小:40.00 KB
- 文档页数:7


什么是动物的应激应激是动物在外界和内在环境中,一些具有损伤性的生物、物理、化学以及特种心理上的强烈刺激(应激因素)作用于机体后,随即产生的一系列非特异性全身性反应,或曰非特异性反应的总和。
应激对动物有不利的一面,但是低度应激下平缓进入适应阶后,甚至可以提高动物的生产力和抵抗力应激反应的机理应激反应的发生过程,其机制十分复杂,动物在应激状态下通过神经—内分泌系统几乎动员了所有的器官和组织应对激源刺激,机体通过极复杂的神经体液调节,保持体内生理生化过程协调与平衡,建立新的稳恒态。
中枢神经系统尤是大脑皮层起整合作用,交感—肾上腺髓质系统、下丘脑—垂体—肾上皮质轴、下丘脑—垂体—甲状腺轴、下丘脑—垂体—性腺轴等起着执行作用。
应激因素(1)物理性应激因素:过热、过冷、强幅射、贼风、强噪声等;(2)化学性应激因素:畜禽舍中的氨、硫化氢、二氧化碳等有毒有害气体;(3)饲养性应激因素,饥饿或过饱、日粮营养不均衡、急剧变更日粮与饲养水平、饮水不足和水质不卫生或水温过低、饲料投饲时间过长等(4)生产性应激因素,饲养规程变更、饲养员更换、断奶、称重、转群、饲养密度过大、组群过大等;(5)外伤性应激因素,去势、打耳号、断尾等;(6)心理性应激因素,争斗、在畜群中的等级地位、惊吓、饲养员的粗暴对待等;(7)运输应激因素,装卸和运输行程中的不良条件及刺激等;(8)兽医预防或治疗应激因素,疫苗注射、消毒、兽医治疗等。
应激的发生过程惊恐反应阶段,亦称动员阶段或紧急反应阶段(总持续时间一般6~48小时),亦称肾上腺反应阶段,动物受到应激源的刺激后机体的早期反应。
从生理生化角度,可将该阶段划分为休克相(Shock phase)和反休克相(Countershock phase), 前者表现为体温和血压下降,血液浓稠,神经系统受到抑制,肌肉紧张度降低,进而发生组织降解,低血氯、高血钾、胃肠急性溃疡,机体抵抗力低于正常水平。
体克相持续几小时至一天,随后进入反休克相,机体防御系统动员、重组而加强,血压上升,血钠和血氯增加,血钾减少,血糖升高、血液粘度增加,分解代谢加强,胸腺,脾脏和淋巴系统萎缩,嗜酸性白细胞和淋巴细胞减少,肾上腺皮质肥大、肾上腺皮质激素分泌增加,机体总抵抗力提高,甚或高于正常水平。

12类用于水产养殖的抗应激饲料添加剂鱼虾类水生动物的集约化人工养殖过程中,难免面临着越来越多应激因素的刺激影响,如水质污染、气候冷热突变、惊吓、饵料营养不良、有害物的侵袭、捕捞、运输、喂养管理不当等等,这些不同程度地应激反应.不但能直接影响动物体的正常生长发育,而且严重者可导致鱼虾的机体免疫力下降.诱发患病及至死亡。
因此.减少或消除养殖鱼虾的应激因素,除了实行科学饲养管理外.还应根据具体情况.在饵料中添加有关抗应激饲料添加剂。
1 维生素C(Vc)Vc是动物机体不可缺少的营养物质。
Vc具有提高鱼虾生产性能.增强机体免疫力和抗病力,同时可增强鱼虾抗应激能力。
王伟庆(1996)报道,加适量的Vc能有效提高中国对虾的缺氧耐受力。
Merchine(1995)采用Vc单磷酸酯(AP)培养的artemia nauptii饲喂组对盐度应激抵抗力强。
Dabrowska(1991)也报道。
适量的Vc会减轻鲤鱼的应激性损伤。
王昆伦等(2000)报道,在受到工业废水污染的水环境中。
在试验组的尼罗罗非鱼的饲料添加Vc一磷酸酯按130mg/g 干饲料.对照组喂基础饵料.试验结果.试验组鱼的发病率比对照组下降47.72%,死亡率下降64.17%。
唐古文等(1997)、罗金萜(2000)均报道.Vc对鱼虾有很强的抗热应激作用。
2 核苷酸核苷酸已被证实为一种免疫增强剂.可减轻对虾应激反应。
据Koppel(1998)报道,以始重6.97 g的中成对虾为试验对象.通过经常改变水的咸度为应激因素.试验结果表明,食含核苷酸饵料对虾的试验组存活率达93.4%.而对照组存活率最低仅53.3%。
蔡明娃(1999)试验.用白色斑点病毒(WSSA)人工感染对虾,饲喂含0.2%核苷酸饵料的对虾存活率达90%,饲喂含0.1%核苷酸的存活率达60% ,而对照组存活率仅27%。
3 腐植酸腐植酸是天然的“绿色”动物饲料添加剂。
其在动物体内具有免疫功能,促进动物生长、抗炎、抗溃疡,促进肠道微生态平衡,抗环境污染感染应激功效。

小寒动物世界中的冬眠启示随着气温逐渐降低,越来越多的动物开始进入冬眠状态。
冬眠是指某些动物在寒冷季节中进入低代谢状态的现象,以保证它们在无食物、水源或营养物的情况下生存。
然而,冬眠的启示并不仅仅是给动物们在寒冷季节的生存提供了保障,它们也为我们人类带来了有益的启示和教训。
一、关于生物应激冬眠是一种应对环境压力的方式,即通过降低代谢速度来减少动物对食物、水和氧气的需求,以应对环境的变化。
在人类生活中,我们也经常遇到各种应激事件,比如工作压力、生活压力等。
我们可以从冬眠的启示中看到,当我们遇到应激事件时,不要惊慌失措,而是要积极应对,降低代谢速度,减少消耗和浪费,以更好地应对生活中的压力和挑战。
二、关于节约能源冬眠期间,动物的代谢速度降低,需要的能量也相应减少。
这启示我们应该更加珍惜能源,减少浪费。
例如,在我们的日常生活中,可以通过减少不必要的照明和电器使用,关注家庭和企业的能源使用,以协助减缓气候变化并节约能源。
三、关于身心健康冬眠能够帮助动物保持身体健康,并缓解肌肉和骨骼所承受的压力。
同样地,在我们的日常生活中,放松自己的身体和心灵是非常重要的。
我们可以通过运动、阅读和冥想等方式来降低压力,缓解压力对身体和心理健康的影响。
四、关于进化动物进化过程中的冬眠现象,是对环境变化的一种适应方式。
我们在人类社会的进化过程中,面临着各种环境变化,例如技术进步、人口增长、人类对环境的影响等。
我们可以从动物的冬眠启示中,认识到适应环境变化的重要性,不断提高自己的适应能力和抗压能力。
综上所述,小寒动物世界中的冬眠现象,为人类提供了很多有益的启示和教训,引发我们的思考,为我们探索自己和环境之间的积极关系提供了借鉴。
我们希望,通过对小寒动物世界的观察和研究,我们可以更好地了解自然和生命的规律,为人类的可持续发展带来更多的启示和借鉴。

养殖场中降低动物应激的方法养殖场中的动物应激,是指动物在特定环境下遭受到的各种不利刺激,包括噪音、光线、环境温度等。
这些应激因素对动物的健康和生产效益有着直接的影响,因此,养殖场需要采取一系列的措施来降低动物的应激。
以下是一些降低养殖场动物应激的方法:1. 提供合适的饲料和饮水:饲养场可以根据不同动物的需求,提供适宜的饲料和饮水,确保动物的饮食和水分摄入达到平衡。
饲料和饮水的种类和质量都会对动物的生理和心理状态产生影响。
2. 维持舒适的环境温度:动物对环境温度的适应能力有限,如果环境温度过高或过低,都会给动物带来应激反应。
因此,养殖场需要根据不同季节和动物的特点,合理调节环境温度,为动物创造舒适的生活环境。
3. 注意动物的疾病防控:养殖场需要建立健全的疫病防控制度,定期进行疫苗接种和疾病检测,并及时给予治疗。
疾病的爆发会导致动物免疫力下降,进而增加其应激的程度。
4. 减少噪声干扰:噪声是养殖场中常见的应激因素之一。
养殖场可以通过设置隔音墙、调整设备运转时段等方式,降低噪声对动物的干扰,提供安静的生活环境。
5. 规律的作息时间:动物对于作息时间有一定的习惯,如果作息时间不规律,容易给动物带来应激。
因此,养殖场可以制定出一份科学的作息时间表,确保动物能够按照规律的作息时间进行活动。
6. 提供良好的栖息和活动空间:动物需要有足够的空间进行栖息和活动。
栖息和活动空间的不足会导致动物产生焦虑和压力,从而增加应激程度。
养殖场可以根据不同动物的需求,提供足够的栖息和活动空间,让动物能够自由活动。
7. 减少人为干扰:养殖场的工作人员在处理动物时需要注意技巧和仪态,尽量减少对动物的人为干扰。
过度的人为干扰会增加动物的应激,甚至引发攻击行为。
8. 提供适宜的社交环境:一些动物对于同伴之间的互动有一定的需求,如果缺乏社交环境,容易产生应激。
因此,养殖场可以根据动物的特点,提供适宜的社交环境,让动物能够与同类进行互动。

种公猪应激分析及预防措施近年来,随着集约化、规模化养猪业的兴起,饲养密度大,人为干扰多,使种猪应激反应综合症的发生率大幅度上升,对猪的种用价值和猪的生长状况都造成了一定的影响。
因此,加强预防工作显得尤为必要。
1.概念应激反应(stress)指由于应激因子(stressor)对动物体的有害作用所引起的非特异性的一切紧张状态。
这是H.Selye(1936)根据机体在寒冷条件下的反应而提出的概念,也可以说是机体遭到侵害而产生反应的状态。
应激反应有时为局部性,如炎症,有时为全身性,如全适应综合症,但不论是局部性的还是全身性的都与作用因子的种类无关,通常是机体的统一反应,因此认为具有适应性的意义。
不过应激反应对动物体也有其破坏作用的一面,即所谓的适应性疾病。
动物处于应激状态时,丘脑下部-脑垂体-肾上腺皮质系统活动增强,因此这些器官的激素分泌增加。
另外,也影响到免疫系统,因而机体的防御机制等也发生变动。
J.J.Christiam(1950)曾主张应激反应与哺乳类动物的个体数的自然控制有关,即在高密度条件下个体间的互相干挠,会产生社会心理性的应激反应,因而可带来生殖机能的降低和死亡率的增加,结果,严重产生应激反应的种群趋向于崩溃,但也有反对这种学说的。
机体受到各种有害刺激时,血中促肾上腺皮质激素和糖皮质激素增多,并引起一系列全身反应以抵抗有害刺激,成为应急反应。
猪应激综合症(PSS),即猪受到外界多种不良因素(如转圈分群、捕捉、特异气味、噪声恐吓、高温缺水、饲料变更、长途运输等)的异常刺激而引起的非特异性的防御反应的统称。
2.应激反应在品种之间的表现2.1、PSS突变基因的由来、作用与扩散20世纪20年代前后,在比利时布拉邦特省的皮特兰镇,人们利用当地诺尔曼猪与巴克夏猪杂交,选育背脂少、肌肉极端丰满的“双肌型”猪,称为皮特兰猪,1953年被列入比利时种猪名录。
皮特兰猪肌肉极端丰满的特征是由一个突变基因所决定的,这个突变基因最初基于它对氟烷敏感,而被称为氟烷基因。




畜禽应激反应(上)任邦军【期刊名称】《四川畜牧兽医》【年(卷),期】2016(043)004【总页数】2页(P52-53)【作者】任邦军【作者单位】四川省梓潼县定远乡畜牧兽医站,四川梓潼 622150【正文语种】中文【中图分类】S858.225.5应激或应激反应又称狩猎式反应,指动物受到应激源或应激因子的刺激而处于一种非特异性“紧张”状态时,出现的一系列神经和内分泌反应,这些反应可调整、维持机体内环境的相对稳定,提高机体对外环境的适应力。
1.1 环境变化风雨雷电、冰霜雪霾、电离辐射、环境污染、温度过低、温度过高、忽冷忽热、贼风侵袭等可导致畜禽发生应激。
1.2 惊吓噪音、强光、其他动物的惊扰等可导致畜禽发生应激。
1.3 长途运输长途密集运输、捕捉、饥饿、疲劳、恐惧等可导致畜禽发生应激。
1.4 饲养管理不当不适当的饲养管理,如随意组群、饲养密度过大、突然换料,环境卫生差、阴暗、潮湿,中毒、缺氧、粗暴驱赶等可导致畜禽发生应激。
断喙、断角、断奶、去势、强制换羽,创伤、品改、手术、失血、感染治疗等也可导致畜禽发生应激。
1.5 免疫接种免疫程序不合理,连续接种或接种方法不当,疫苗质量差等可导致畜禽发生应激。
2.1 第一阶段为警觉反应期,又称动员期,其分为休克期和抗休克期。
畜禽休克期神经系统受到抑制,血压、体温下降,血糖降低,白细胞数减少,抗休克期其交感神经兴奋性增高,肾上腺活动增强、循环血量增加、血压上升、体温升高、血糖浓度增高、嗜中性粒细胞增多。
如果应激源很快消失,将不会对机体造成严重危害,有时还能提高机体抵抗力。
如果应激源继续作用,便进入第2阶段。
2.2 第二阶段为抗休克期的继续,是机体克服应激源造成的损害,维护内环境的稳定,趋向恢复的阶段。
这一阶段畜禽各种反应逐渐减弱,原来出现的症状逐步缓解或消失,反之则畜禽进入衰竭期。
2.3 第三阶段又称衰竭期,由于应激源强度过大或作用持久,畜禽因不能克服应激源造成的伤害,内环境的稳定性受到致命的破坏而出现衰竭、死亡。
寒冷应激条件下脂多糖诱导肺损伤过程中MPO mRNA表达水平的变化朱雪;王亮【摘要】目的探讨寒冷应激条件下大鼠肺损伤过程中MPO mRNA表达水平变化,及肺组织病理形态学变化.方法将Wistar大鼠随机分为正常对照组(N)、寒冷刺激+脂多糖组(CL)、脂多糖组(L),每组各10只.CL组在实验前4天每天予寒冷刺激4 h,CL、L组在第5天及第12天分别予气管内滴加脂多糖.至实验第19天采集大鼠肺组织观察肺组织形态学变化,RT-PCR半定量法检测肺组织匀浆中MPO mRNA 的表达.结果模型组肺组织病理观察与对照组相比有明显变化,MPO mRNA的表达各组间比较有显著统计学意义(P<0.01).结论寒冷刺激后可造成大鼠免疫功能一定程度的降低,在这种情况下的细菌内毒素刺激可造成肺组织MPO合成和分泌的增加及肺组织更严重的损伤.【期刊名称】《泰山医学院学报》【年(卷),期】2010(031)003【总页数】4页(P174-177)【关键词】寒冷刺激;脂多糖;肺损伤;MPO;病理;动物实验【作者】朱雪;王亮【作者单位】山东中医药大学,山东,济南,250014;济南市第六人民医院,山东,济南,250020【正文语种】中文【中图分类】R56寒冷应激过强可使机体内环境的稳定性发生变化,引起机体的许多异常反应,近年国内外对冷应激损伤的免疫及分子机制研究有了很大进展[1-5],但未见寒冷应激条件下脂多糖诱导肺损伤过程中髓过氧化物酶(myeloperoxidase,MPO) mRNA 表达水平变化的报道。
MPO是一种重要的含铁溶酶体,为中性粒细胞所特有的酶类,能反映中性粒细胞浸润及活性[6]。
1 材料与方法1.1 实验动物与分组SPF级健康雄性Wistar大鼠30只,体质量(200±20 g)。
采用随机数字表法分为正常对照组(N)10只,寒冷刺激+脂多糖组(CL)10只,脂多糖组(L)10只。
1.2 主要试剂及仪器脂多糖(Life Sciences 公司,LOT2011/08);TAKARA RT-PCR 试剂盒 Version 3.0。
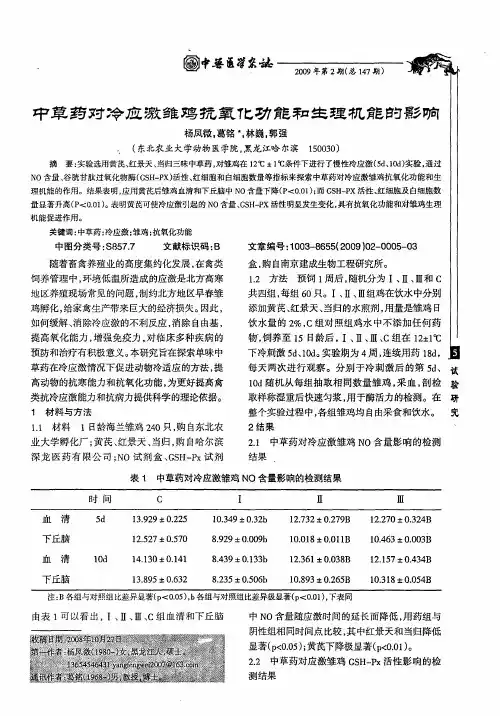
N-乙酰半胱氨酸和冷应激对肉鸡肠道组织形态、能量代谢和抗氧化能力的影响王惠云;廖满;王蕾;赵迪;易丹;刘玉兰;丁斌鹰【期刊名称】《中国畜牧杂志》【年(卷),期】2018(054)002【摘要】本试验旨在研究N-乙酰半胱氨酸(NAC)和冷应激对肉鸡肠道组织形态、能量状况和抗氧化能力的影响.试验将200只1日龄健康雄性科宝肉鸡随机分为4个处理组:对照组(基础日粮)、NAC组(基础日粮+0.1%NAC)、冷应激组(基础日粮)与冷应激+NAC组(基础日粮+0.1%NAC),每组5个重复,每个重复10只鸡.试验第1~2周按常规饲养方法控制温度,试验第3~6周冷应激组和冷应激+NAC组的温度控制在(10±2)℃,而对照组与NAC组维持在(26±2)℃.结果表明:冷应激处理使肉鸡平均日采食量、平均日增重、空肠低氧诱导因子1 (HIF-1) mRNA水平显著下降(P<0.05),饲料转化率、回肠一磷酸腺苷(AMP)含量和空肠隐窝深度极显著升高(P<0.01);日粮添加0.1% NAC显著提高了冷应激肉鸡肠道绒毛高度(VH)、十二指肠与空肠绒毛高度与隐窝深度比值(VH/CD)、空肠三磷酸腺苷(ATP)和腺苷酸池(TAN)含量、回肠ATP和二磷酸腺苷(ADP)及能荷(AEC)含量、十二指肠超氧化物歧化酶(SOD)和过氧化氢酶(CAT)活力、回肠谷胱甘肽过氧化物酶(GSH-Px)活力、空肠血红素加氧酶(HMOX) mRNA水平(P<0.05).因此,日粮中添加0.1%的N-乙酰半胱氨酸可改善肠道组织形态和能量状况,提高冷应激肉鸡肠道抗氧化能力.%This experiment was conducted to investigate the effects of N-acetylcysteine (NAC) and cold stress on intestinal morphology,energy status and antioxidant capacity in broilers.The experiment was divided into4 groups according to the factorial design (2 x 2).Two hundred 1-day-old male Cobb chickens were randomly divided into 4 groups:control group (basal diet),NAC group (basal diet + 0.1% NAC),cold-stressed group (basal diet),and cold stress + NAC group (basal diet + 0.1% NAC).Each group had5 replicates with 10 chickens per replicate.The temperature form first to second week was controlled with conventional feeding management,whereas that from the third to sixth week in the cold-stressed and cold-stress + NAC groups was maintained at (10 ± 2)℃,which was differed with the other groups (26 ± 2)℃.The humidity was 50%~60%.The results showed that cold stress treatment decreased the broiler average daily feed intake (ADFI),average daily gain (ADG),and HIF-1 mRNA level in the jejunum (P<0.05),whereas increased the feed conversion ratio (FCR),the level of AMP (P<0.05),and crypt depth (CD) in the ileum and jejunum (P<0.01) of broilers.Additionally,diet supplemented with 0.1%NAC increased (P<0.05) the intestinal villus height (VH),VH/CD in the duodenum and jejunum,the levels of ATP and TAN in the jejunum,the levels of ATE ADP,and AEC in the ileum,the activities of SOD and CAT in the duodenum,the GSH-Px activity and the HMOX mRNA level in the ileum of cold-stressed broilers.Taken together,diet supplemented with 0.1% NAC could improve the intestinal morphology and energy status and increase the intestinal antioxidant capacity of cold-stressed broilers.【总页数】8页(P52-59)【作者】王惠云;廖满;王蕾;赵迪;易丹;刘玉兰;丁斌鹰【作者单位】动物营养与饲料安全湖北省协同创新中心,动物营养与饲料科学湖北省重点实验室,武汉轻工大学动物科学与营养工程学院,湖北武汉 430023;动物营养与饲料安全湖北省协同创新中心,动物营养与饲料科学湖北省重点实验室,武汉轻工大学动物科学与营养工程学院,湖北武汉 430023;动物营养与饲料安全湖北省协同创新中心,动物营养与饲料科学湖北省重点实验室,武汉轻工大学动物科学与营养工程学院,湖北武汉 430023;动物营养与饲料安全湖北省协同创新中心,动物营养与饲料科学湖北省重点实验室,武汉轻工大学动物科学与营养工程学院,湖北武汉430023;动物营养与饲料安全湖北省协同创新中心,动物营养与饲料科学湖北省重点实验室,武汉轻工大学动物科学与营养工程学院,湖北武汉 430023;动物营养与饲料安全湖北省协同创新中心,动物营养与饲料科学湖北省重点实验室,武汉轻工大学动物科学与营养工程学院,湖北武汉 430023;动物营养与饲料安全湖北省协同创新中心,动物营养与饲料科学湖北省重点实验室,武汉轻工大学动物科学与营养工程学院,湖北武汉 430023【正文语种】中文【中图分类】S831.5【相关文献】1.三乳酸甘油酯对冷应激肉鸡肠道抗氧化能力和能量代谢状况的影响 [J], 廖满;段瑞;谢佳倩;周建;杨书慧;丁斌鹰;刘玉兰;易丹2.N-乙酰半胱氨酸对肉鸭肝脏能量代谢状况及组织抗氧化指标的影响 [J], 谭灵琳;侯永清;易丹;廖满;王蕾;丁斌鹰3.N-乙酰半胱氨酸对樱桃谷鸭肠道能量代谢和抗氧化功能的影响 [J], 杨书慧; 毕云天; 程强; 谭灵琳; 易丹; 侯永清; 郭双双; 丁斌鹰4.卵内注射亮氨酸对慢性热应激雌性肉鸡生长性能、肠道组织形态和抗氧化能力的影响 [J], 崔洋洋;韩国锋;李延森;李春梅5.L-精氨酸和α-酮戊二酸对热应激肉鸡肠道吸收功能、抗氧化能力、能量代谢的影响 [J], 晏利琼;廖满;谢佳倩;谭灵琳;丁斌鹰;杨书慧;刘玉兰;杨鹰因版权原因,仅展示原文概要,查看原文内容请购买。
关于寒冷应激对动物影响的研究进展摘要本文阐述了寒冷应激的主要概念并综述了动物冷应激时神经系统的生理机制,及其对动物生产性能、免疫功能、抗氧化功能和血液中相关生理指标的影响,以及相关基因的研究进展。
通过对动物冷应激时动物机体内的一系列变化规律的总结,为提高管理效率,减少动物冷应激带来的经济损失,以及冷应激相关科学研究的开展奠定了基础。
关键词动物冷应激; 神经内分泌反应; 生产性能; 生理免疫加拿大病理学家Hans Selye 在1936年提出了应激学说,应激即内部环境或外界环境的具有伤害性刺激扰乱内部环境的平衡稳定所导致的一种状态。
应激在典型情况下可分三个阶段: 第一阶段为惊恐反应或动员阶段,这是机体对应激原作用的早期反应; 第二阶段为适应或抵抗阶段,机体克服了应激原作用而获得了适应; 第三阶段为衰竭阶段,表现很像惊恐反应,但反应程度急剧增强,出现各处营养不良,异化作用占优势的现象[1]。
就动物而言,创伤、环境的冷热、惊吓等均是应激源。
环境温度的高或低所导致的动物热、冷应激直接影响动物的生产性能、繁殖性能、自身免疫以及所提供的产品质量,是制约畜牧养殖业发展的重要因素之一。
在北方高寒地区, 早春及晚秋母猪产仔, 冬季母牛产犊及早春孵化雏鸡等是促进畜牧业飞速发展的必要环节, 但因环境低温所导致的冷应激常引起动物生长缓慢、抗病性差, 甚至死亡是制约北方畜牧业发展的主要因素之一。
因此, 解决这一影响畜牧业生产中的难题, 目前已成为营养生理、家畜环境营养学研究的前沿和热点。
寒冷应激在动物的神经内分泌、生理、组织免疫等学科的研究均得到了较系统的成果。
这里以总结冷应激对动物的影响,为预防动物的冷应激,培育耐寒、抗病和抗应激的动物品种奠定基础。
一、寒冷应激对动物机体影响的生理学机制1 寒冷应激的一般概念应激通常是指动物受到频率比较大, 持续时间长或时间虽短但变化剧烈的刺激时所引起的机体反应, 这种反应主要使动物内环境稳定性、生理和行为等发生改变。
应激所引起机体的非特异性变化叫做全身适应综合症( GAS) ,凡能引起机体出现GAS 的刺激即称为应激原( Stressor ) , 而寒冷应激即为应激原之一。
关于寒冷应激, 人们通常都只是讨论它所引起的影响或是一般症状, 到现在似乎还没有一个十分精确的定义, 对不同动物, 寒冷的概念亦不一样, 寒冷应激的范围和程度也有所不同。
动物急性( 短期) 暴露于寒冷环境中, 开始产生冷应激( Cold stress) 反应, 这一反应称之/ 报警反应( Alarm reaction)。
在慢性( 长期) 冷暴露( Cold ex posure) 情况下, 冷应激首先产生的报警反应消逝后, 动物的代谢发生特异性变化, 持续稳定的寒冷刺激, 使儿茶酚胺和甲状腺素分泌增强, 机体的能量代谢持续升高, 但体热绝缘即保温能力( 被毛厚实, 血管收缩) 不受影响,动物的存活时间可以延长, 这种情况称为惯习或气候适应( Acclimatio n) 。
随着季节性变化或迁换地理环境机体生理调节逐渐发生变化, 例如, 由秋季进入严冬后, 动物的保温能力明显提高, 但能量代谢变化不大, 有的甚至反而降低, 这种情况称为冷驯化( Acclimat ization) 。
2 动物对寒冷应激的反应寒冷应激反应的机制十分复杂, 几乎涉及所有的器官和组织, 其中, 中枢神经系统及其高级部位( 大脑皮层) 是起整合调节作用的主导部门, 通过交感- 肾上腺髓质系统、下丘脑- 垂体- 肾上腺皮质系统及下丘脑- 垂体- 甲状腺轴等实现的。
根据Selye 用大鼠所做的试验表明, 应激在典型情况下可分三个阶段: 第一阶段为惊恐反应或动员阶段( Alarm reactionor stag e of mobilizatio n) , 这是机体对应激原作用的早期反应,此期机体尚未获得适应。
第二阶段为适应或抵抗阶段( Stageof adaptat ion or r esistance) , 机体克服了应激原作用而获得了适应, 诸多表现与惊恐反应相反, 新陈代谢趋于正常, 同化作用占优势。
第三阶段为衰竭阶段( Stag e of ex haustion) , 表现很象惊恐反应, 但反应程度急剧增强, 出现各处营养不良, 异化作用占优势。
[ 2]调节体温的基本中枢在下丘脑。
当外界环境温度下降引起体内温度下降超过体温调定点的阈值( 如37 ℃) 时, 则机体冷敏神经元兴奋, 发放的神经冲动频率增加, 促使产热活动加强, 如骨骼有紧张性增加, 皮肤血管收缩等, 同时热敏神经元放电频率减少, 使散热活动受到抑制, 结果使体温不致过度降低,从而保持体温相对恒定。
二、寒冷应激对动物能量代谢的影响低温环境对动物能量代谢影响明显。
动物暴露于临界温度下限的环境中时, 机体静止能量代谢率升高同时产热量增加。
绵羊暴露于0 ℃与20 ℃比较其产热量增加2. 14 倍。
研究报道, 在临界温度下限时, 外界温度每降1 ℃, 静止能量代谢率成年羊升高4 . 2 KJ /kg - 0. 75 d- 1 , 生长期犊牛升高2. 5KJ/kg- 0. 75d- 1 , 肉牛升高2. 9 KJkg - 0. 75d- 1。
肉牛饲养在30 ℃环境中时, 静止能量代谢率为13. 08KJ/kg- 0. 75 d- 1, 但将牛饲养在17. 4 和12. 7 ℃的冬季室外环境中时, 其静止能量代谢率分别为16. 51 和17. 93KJ/kg- 0. 75d- 1。
将泌乳绵羊暴露于21℃和0℃环境中, 第1、21d 和41 d 后, ℃0试验组的产热量较21℃组分别提高20%( P< 0.05) 、43%( P< 0. 01) 和55%( P< 0. 001) 。
而且所有冷暴露级乳脂含量亦明显增加( P< 0. 05) , 冷暴露21 d 后, 乳蛋白和乳糖含量明显增加, 短链脂肪酸在冷暴过程中相对降低。
说明冷应激后乳中能量损失有增加趋势, 但每日乳产量无明显改变。
当动物进行急性冷暴露时, 实际的日粮能量主要从生产功能而转向产生大量的体热, 以维持体温恒定。
若热量产生不足, 可引起生长发育的继发性改变及某些疾病的产生, 严重者将导致死亡。
McBr ide , G. E[ 3] 将青年羔羊置于0℃和21℃的环境中饲养, 10 周后发现在0 ℃环境中饲养的羔羊, 其跖骨长度缩短, 冷暴露组的皱胃和甲状腺重量有轻微的增加( P< 0. 01) , 对干物质和氮的消化率无明显影响, 但体内氮的潴留因冷暴露而显著降低( P< 0. 05) 。
他们还将马从18 ℃的室温环境移入- 10 ℃和- 40 ℃的室外低温环境进行冷暴6 h 后发现, 二者的代谢率明显提高, 其中在- 40 ℃条件下, 代谢率比冷暴前提高了142%, 冷暴期间血浆T3 和T 4 的基础浓度亦明显增加( P< 0. 05) , 饲养于0. 8 ℃环境中的绵羊与17. 7 ℃环境中饲养的绵羊相比, 其干物质消化率下降, 水的消耗亦降低, T3 和T4 浓度升高( P< 0. 05) , 并且认为环境温度对消化的影响是由于饲料在消化道中的停留时间发生改变而引起的。
李玉芝等[ 4] 利用东北民猪研究表明, 当温度下降至13 ℃时, 仔猪平均产热量由20 ℃的29. 2 kJ/ kg0. 75h 和27 ℃的31. 1kJ/ kg0. 75 h 增加为38. 0 kJ/ kg0. 75h。
任涛认为[ 5] , 寒冷应激使鸡的基础代谢率升高, 能量代谢和采食增加, 呼吸加深, 耗氧量增加, 使心输出量和血流量以及肺动脉血压升高, 导致右心室血流量和压力超负荷, 产生右心室肥大, 功能衰竭, 进而产生腹水。
如果产生进行性腹水时, 因为补偿性增加静脉血压, 又会导致淋巴液回流受阻, 从而加重腹水现象, 最后鸡由于胸腔压力增加产生肺水肿, 致使呼吸衰竭而死。
三、冷应激对动物生理免疫的影响机体免疫功能的高低取决于胸腺、腔上囊和脾等免疫器官的生理状况,它们在淋巴细胞的形成、诱导、分化和对抗原的刺激进行免疫过程中起着重要作用。
冷应激可引起脾脏和淋巴结对刀豆素以及植物血凝素A 的反应性减弱,自然杀伤细胞的杀伤活性下降,对靶细胞的攻击力下降; 抑制淋巴细胞特别是T 淋巴细胞的有丝分裂和DNA 合成; 损伤浆细胞,阻碍免疫球蛋白的合成和分泌; 抑制巨噬细胞对抗原的吞噬和处理,从而对机体的免疫功能造成一定的影响。
Shu 等对小鼠进行冷水应激试验发现,在1 d 的冷应激反应中,CD4+T 细胞比率及血液和脾淋巴细胞内自然杀伤细胞的杀伤活性均下降; 而 5 d 的冷应激后,以上指标均增加[6]。
袁学军等则发现,伊褐红雏鸡在急性冷应激24 h 内T淋巴细胞数极显著减少; 在冷应激10 d 后,淋巴细胞的总数极显著减少,但T 淋巴细胞数极显著增加[7]。
王玉海等发现,贵妃雏鸡和海兰雏鸡在冷应激( 比常温低5 ℃) 1 h 期间,碱性粒细胞和脾脏中的肥大细胞数减少; 但2 和4 h 后数目又显著增加; 在冷应激15 d 后,脾脏免疫水平略有提高,而胸腺的免疫水平显著提高[8-9]。
井霞等也同样发现,慢性冷应激会导致荷斯坦奶牛的外周血白细胞总数、淋巴细胞数和单核细胞数均显著降低,CD21+B 淋巴细胞百分率和CD3+T淋巴细胞百分率均有所下降[10]。
以上试验均表明,冷应激会对动物的免疫功能造成一定程度的抑制,使得机体的免疫功能下降。
此外,Rybakina 等指出,冷暴露可使小鼠血清皮质酮的含量显著升高,机体的体液免疫受到明显的抑制[11]。
Frank 等在仔猪的冷应激试验中发现,血浆促肾上腺皮质激素、皮质醇和白细胞介素等含量较高[12]。
吕彩霞等证实,长期冷应激可刺激机体生成多种血清免疫抑制因子,抑制白细胞介-2、肾上腺糖皮质激素及前列腺素的产生[13]。
说明冷应激加剧了机体的应激和免疫反应,使机体动员了更多的免疫资源。
这些化合物使成熟和已分化的淋巴器官中的环腺苷酸的含量提高,导致淋巴细胞的免疫功能降低,但在长期应激中,机体有一个逐渐适应的过程。
四、冷应激对动物抗氧化功能的影响研究发现,冷应激可改变机体的促氧化和抗氧化间的平衡,通过黄嘌呤氧化酶活性的升高及髓过氧化酶活性的降低,使机体产生过多的自由基,导致脂质过氧化作用增强,诱发氧化损伤。
Kaushik 等发现,慢性冷应激可使小鼠脑、心脏、肾脏、肝脏和小肠的超氧化物歧化酶( SOD) 的活性显著下降,心脏、肝脏和小肠的过氧化氢酶( CAT) 的活性显著下降,但肾脏中的CA T 的活性显著提高[14]。