第八章脂类代谢
- 格式:pdf
- 大小:180.76 KB
- 文档页数:8
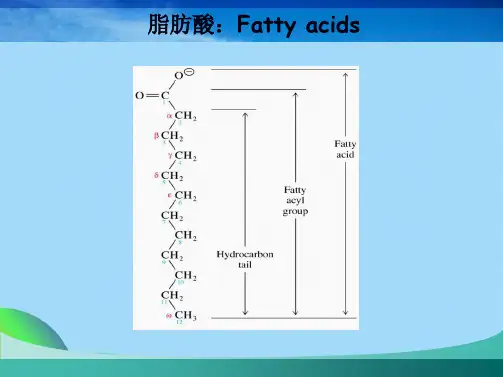



第八章+脂类代谢(王镜岩考研生物化学)第八章脂类代谢(对应教材第28章,29章)湖大生物学院一、脂质的消化、吸收和传送n1、脂类–什么是脂类生物体内通过弱极性或非极性溶剂抽提得到的不溶于水的有机分子李新梅湖大生物学院16人体内最重要的脂肪酸3n李新梅湖大生物学院胰脂酶n 李新梅湖大生物学院脂肪的吸收和转运–4、人体血浆中脂蛋白(第2章117-119页第29章292-294页)n(1)脂蛋白形式结构–以图中乳糜微粒为例,在脂蛋白中,疏水脂类构成核心,外面围绕着极性脂和载脂蛋白,以增加溶解度。
nn (nn 李新梅湖大生物学院n–李新梅湖大生物学院二、脂肪酸分解代谢n 1、脂肪组织释放脂肪酸受激素的调控(脂肪动员)n (1)甘油三酯脂肪酶分解脂肪–脂肪组织中脂肪颗粒的主要成分是三脂酰甘油–在激素作用下,甘油三酯脂肪酶被激活–脂肪经甘油三酯脂肪酶降解生成脂肪酸和甘油脂肪酸则转运到其他组织甘油绝大部分经糖异生途径转化为葡萄糖;李新梅湖大生物学院甘油三酯脂肪酶李新梅湖大生物学院甘油三酯n李新梅湖大生物学院李新梅湖大生物学院(3)甘油的分解和糖异生李新梅湖大生物学院n (1)甘油糖异生经过哪些过程–甘油→3-磷酸甘油→磷酸二羟丙酮→1,6-二磷酸果糖↓3-磷酸甘油醛↗→6-磷酸果糖→6-磷酸葡萄糖→葡萄糖(2)催化的酶是哪些?l 甘油磷酸激酶耗ATPl 3-磷酸甘油脱氢酶磷酸丙糖异构酶l 醛缩酶果糖二磷酸酶6-磷酸葡萄糖异构酶l 葡萄糖-6-磷酸酶2n 李新梅湖大生物学院β-氧化即每次从脂肪酸链上降解下来的是2碳单位n (1)脂肪酸的活化n(2)脂酰CoA转运入线粒体基质李新梅湖大生物学院②肉碱穿梭系统的成员肉碱脂酰转移酶I 膜间隙肉碱脂酰转移酶II 基质李新梅湖大生物学院–n n––(3)脂酰辅酶A的β氧化n SCoARCH 烯脂酰CoA 脱氢酶RCH 李新梅湖大生物学院nn3、脂肪酸——n李新梅湖大生物学院n (––其他产物:16分子CO –软脂酸的标准自由能是解产生自由能是773.8千卡,能量转化率为30%李新梅湖大生物学院4、奇数脂肪酸的氧化n 奇数碳脂肪酸在最后一轮β中生成丙酰CoA–反刍动物中利用奇数脂肪酸提供大多数哺乳动物很罕见奇数脂肪酸–在哺乳动物的肝脏中,丙酰的催化反应转化为琥珀酰?羧化,消旋,变构琥珀酰CoA 转换成草酰乙酸,进入糖异生途径。





第八章脂类代谢一、填空题:1.大部分饱和脂肪酸的生物合成在中进行。
2.自然界中绝大多数脂肪酸含数碳原子。
3.脂肪酸生物合成的原料是,其二碳供体的活化形式是。
4.生成丙二酸单酰CoA需要酶系催化,它包含有三种成份、_ 和。
5.饱和脂肪酸从头合成需要的引物是,其产物最长可含有碳原子。
6.人体必需脂肪酸是、和。
7.饱和脂肪酸从头合成的还原力是,它是由代谢途径和转换所提供。
8.大于十六碳原子的脂肪酸是生物体内相应的各个系统的酶催化合成。
9.在真核生物中,不饱和脂肪酸的脱饱和是通过途径完成的,催化反应的酶叫。
10.硬脂酸(C18)经β-氧化分解,循环次,生成分子乙酰CoA,FADH2和NADH。
11.脂肪酸β-氧化是在中进行的,氧化时第一次脱氢的受氢体是,第二次脱氢的受氢体是,β-氧化的终产物是。
12.真核生物中,一摩尔甘油彻底氧化成C02和H20生成摩尔A TP。
13.在油料种子萌发的时候,由脂肪酸分解生成的通过生成琥珀酸,再进一步生成后通过途径合成葡萄糖,供幼苗生长之用。
14.乙酰COA主要由、和降解产生。
二、选择题(只有一个最佳答案):1.在高等动、植物中,脂肪酸以下列哪种形式参与三酰甘油的生物合成( )①游离脂肪酸②脂酰ACP ③脂酰CoA④以上三种均不是2.脂肪酸生物合成中,将乙酰基运出线粒体进入胞液中的物质是( )①CoA②肉碱③柠檬酸④以上三种均不是3.1分子十八碳脂肪酸经β-氧化和三羧酸循环净产生( )A TP①130 ②129 ③147 ④1484.饱和脂肪酸从头合成和β-氧化过程中,两者共有( )①乙酰CoA②FAD ③NAD+④含生物素的酶5.长链脂肪酸从胞浆转运到线粒体内进行β-氧化作用,所需载体是( )①柠檬酸②肉碱③辅酶A④α-磷酸甘油6.脂肪酸从头合成所用的还原剂是( )①NADPH+H+②NADH+H+③FADH2④FMNH27.脂肪酸氧化作用的连续进行与下列哪种酶无关( )①脂酰CoA脱氢酶②烯脂酰CoA水合酶③β-酮脂酰CoA硫解酶④缩合酶8.β-氧化中,脂酰CoA脱氢酶催化反应时所需的辅因子是( )①FAD ②NAD+③A TP ④NADP+9.植物体内由软脂酸(C16)生成硬脂酸(C18)其原料是( )①乙酰CoA②乙酰ACP ③丙二酸单酰CoA④丙二酸单酰ACP10.在脂肪酸的合成中,每次碳链的延长都需要什么直接参加?()①乙酰CoA②草酰乙酸③丙二酸单酰CoA④甲硫氨酸11.合成脂肪酸所需的氢由下列哪一种递氢体提供?()①NADP+ ②NADPH+H+③FADH2④NADH+H+12.脂肪酸活化后,β-氧化反复进行,不需要下列哪一种酶参与?()①脂酰CoA脱氢酶②β-羟脂酰CoA脱氢酶③烯脂酰CoA水合酶④硫激酶13.软脂酸的合成及其氧化的区别为()(1)细胞部位不同 (2)酰基载体不同 (3)加上及去掉2C•单位的化学方式不同(4)β-酮脂酰转变为β-羟脂酰反应所需脱氢辅酶不同 (5)β-羟脂酰基的立体构型不同①(4)及(5) ②(1)及(2) ③(1)(2)(4) ④全部14.β-氧化的酶促反应顺序为:()①脱氢、再脱氢、加水、硫解②脱氢、加水、再脱氢、硫解③脱氢、脱水、再脱氢、硫解④加水、脱氢、硫解、再脱氢15.胞浆中合成脂肪酸的限速酶是()①β-酮酯酰CoA合成酶②水化酶③酯酰转移酶④乙酰CoA羧化酶16.脂肪大量动员肝内生成的乙酰CoA主要转变为:()①葡萄糖②酮体③胆固醇④草酰乙酸17.乙酰CoA羧化酶的变构抑制剂是:()①柠檬酸②ATP ③长链脂肪酸④CoA18.脂肪酸合成需要的NADPH+H+主要来源于()①TCA②EMP ③磷酸戊糖途径④以上都不是19.生成甘油的前体是()①丙酮酸②乙醛③磷酸二羟丙酮④乙酰CoA20.卵磷脂中含有的含氮化合物是:()①磷酸吡哆醛②胆胺③胆碱④谷氨酰胺21.脂肪酸彻底氧化的产物是:( )①乙酰CoA ②脂酰CoA ③丙酰CoA ④H2O、CO2及释出的能量22.甘油氧化分解及其异生成糖的共同中间产物是:()①丙酮酸②2-磷酸甘油酸③3-磷酸甘油酸④磷酸二羟丙酮三、是非题(在题后括号内打√或×):1.生物体内的脂肪酸主要以结合形式存在。
安徽师范大学生命科学院生物专业生物化学教案 13-10-12 1Chapter 8 Lipid Metabolism (7h)
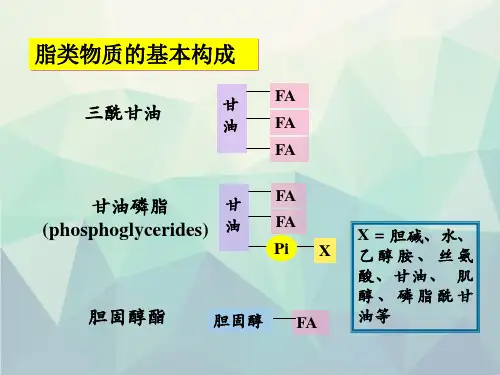
【教学目的】 通过本章教学,使学生掌握脂肪酸的β-氧化及其氧化过程中能量的计算;酮体的生成和利用。熟悉脂肪酸的合成及其与脂肪酸分解的区别;甘油磷脂的降解和合成。了解脂类的分类和生理功能;消化和吸收;胆固醇合成的基本过程及其转化。 【重点难点】 重点:脂肪酸氧化分解的全部过程,酮体的生成及利用,脂酰CoA的跨线粒体膜运输。 难点:脂肪酸的合成代谢,磷脂的代谢。 【教学内容】 Overview: 1. 脂类是脂肪和类脂的总称,是一大类不溶于水而易溶于有机溶剂的化合物。 按其化学组成可分为:单纯脂,复合脂和非皂化脂(油脂的碱水解变成甘油和脂肪酸称皂化)。
单纯脂---是脂肪酸和醇类所形成的脂。 它无极性,又称中性脂。 主要包括脂肪和蜡。(蜡是长链脂肪酸和长链一元醇或固醇形成的酯,如蜂蜡、白蜡、羊毛脂等。)
复合脂---由单纯脂和非脂溶性物质所构成的化合物,包括磷脂和糖脂。
2. Biological function of lipids: 1. 生物体的重要能源;9.3kcal/g脂肪 2. 构成生物膜的基本物质,其中磷脂,糖脂及胆固醇是膜脂类的三种重要类型; 3. 某些萜类(多个异戊二烯单位)及类固醇类物质 如VA,D,E,K,及固醇类激素具有营养代谢及调节功能;
4. 促进食物中脂溶性维生素及必需脂肪酸的吸收; 5. 作为细胞表面物质,与细胞识别,种特异性及组织免疫等有密切关系。
第一节 脂类的酶促水解(Hydrolyses of lipids) 脂类的消化(主要在十二指肠中) 食物中的脂类主要是甘油三酯 80-90% 还有少量的磷脂 6-10% 胆固醇 2-3% 胃的食物糜(酸性)进入十二指肠,刺激肠促胰液肽的分泌,引起胰脏分泌HCO-3 至
小肠(碱性)。脂肪间接刺激胆汁及胰液的分泌。胆汁酸盐使脂类乳化,分散成小微团,在胰腺分泌的脂类水解酶作用下水解。 胰腺分泌的脂类水解酶: ① 三脂酰甘油脂肪酶(水解三酰甘油的C1、C3酯键,生成2-单酰甘油和两个游离的安徽师范大学生命科学院生物专业生物化学教案 13-10-12 2脂肪酸。胰脏分泌的脂肪酶原要在小肠中激活) ②磷脂酶A2(水解磷脂,产生溶血磷酸和脂肪酸) ③胆固醇脂酶(水解胆固醇脂,产生胆固醇和脂肪酸) ④辅脂酶(Colipase)(它和胆汁共同激活胰脏分泌的脂肪酶原)
第二节 甘油三酯的分解代谢(Catabolism of Triacylglycerol) 由脂肪酶催化。 脂肪酶(激素敏感性甘油三酯脂肪酶,是限速酶)、甘油二酯脂肪酶、甘油单酯脂肪酶 肾上腺素、胰高血糖素、肾上腺皮质激素都可以激活腺苷酸环化酶,使cAMP浓度升高,促使依赖cAMP的蛋白激酶活化,后者使无活性的脂肪酶磷酸化,转变成有活性的脂肪酶,加速脂解作用。 胰岛素、前列腺素E1作用相反,可抗脂解。
油料种子萌发早期,脂肪酶活性急剧增高,脂肪迅速水解。
一、甘油的氧化分解(Oxidation of Glycerol) 在脂肪细胞中,没有甘油激酶,无法利用脂解产生的甘油。甘油进入血液,转运至肝脏后才能被甘油激酶磷酸化为3-磷酸甘油,再经磷酸甘油脱氢酶氧化成磷酸二羟丙酮,进入糖酵解途径或糖异生途径。
二、脂肪酸的β氧化(β-Oxidation Pathway) There are several different pathways of fatty acid oxidation in organisms, ainly including:β-oxidation,α-oxidation ,ω-oxidation
(一) β-oxidation of saturated fatty acids 饱和偶数碳脂肪酸的β氧化 1. β氧化学说 早在1904年,Franz 和Knoop就提出了脂肪酸β氧化学说。 用苯基标记含奇数碳原子的脂肪酸,饲喂动物,尿中是苯甲酸衍生物马尿酸。 用苯基标记含隅数碳原子的脂肪酸,饲喂动物,尿中是苯乙酸衍生物苯乙尿酸。 结论:脂肪酸的氧化是从羧基端β-碳原子开始,每次分解出一个二碳片断。 产生的终产物苯甲酸、苯乙酸对动物有毒害,在肝脏中分别与Gly反应,生成马尿酸和苯乙尿酸,排出体外。 β-氧化发生在肝及其它细胞的线粒体内。 2. 脂肪酸的β氧化过程 脂肪酸进入细胞后,首先被活化成酯酰CoA,然后再入线粒体内氧化。 (1)脂肪酸的活化(细胞质) RCOO- + ATP + CoA-SH → RCO-S-CoA + AMP + PPi 生成一个高能硫脂键,需消耗两个高能磷酸键,反应平衡常数为1,由于PPi水解,反应不可逆。 细胞中有两种活化脂肪酸的酶: 内质网脂酰CoA合成酶,活化12C以上的长链脂肪酸 线粒体脂酰CoA合成酶,活化4~10C的中、短链脂肪酸 (2)脂肪酸向线粒体的转运 中、短链脂肪酸(4-10C)可直接进入线粒体,并在线粒体内活化生成脂酰CoA。 安徽师范大学生命科学院生物专业生物化学教案 13-10-12 3长链脂肪酸先在胞质中生成脂酰CoA,经肉碱转运至线粒体内。 肉(毒)碱:L-β羟基-r-三甲基铵基丁酸 脂酰CoA进入线粒体后,在基质中进行β氧化作用 (3)β氧化4个循环的步骤。 脂酰CoA脱氢生成β-反式烯脂酰CoA 脂酰CoA脱氢酶 △2反式烯脂酰CoA水化生成L-β-羟脂酰CoA β-烯脂酰CoA水化酶
L-β-羟脂酰CoA脱氢生成β-酮脂酰CoA L-β羟脂酸CoA脱氢酶 β-酮脂酰CoA硫解生成乙酰CoA和(n-2)脂酰CoA 酮脂酰硫解酶
3. 脂肪酸β-氧化作用小结 脂肪酸β-氧化时仅需活化一次,其代价是消耗1个ATP的两个高能键 (1) 长链脂肪酸由线粒体外的脂酰CoA合成酶活化,经肉碱运到线粒体内;中、短链脂肪酸直接进入线粒体,由线粒体内的脂酰CoA合成酶活化。 (2) β-氧化包括脱氢、水化、脱氢、硫解4个重复步骤 (3) β-氧化的产物是乙酰CoA,可以进入TCA
4. 脂肪酸β-氧化产生的能量 以硬脂酸为例,18碳饱和脂肪酸 胞质中: ⑴活化:消耗2ATP,生成硬脂酰CoA 线粒体内: ⑵脂酰CoA脱氢:FADH2 ,产生2ATP
⑶β-羟脂酰CoA脱氢:NADH,产生3ATP ⑷β-酮脂酰CoA硫解:乙酰CoA → TCA,12ATP (n-2)脂酰CoA → 第二轮β氧化 活化消耗: -2ATP β氧化产生: 8×(2+3)ATP = 40 9个乙酰CoA: 9×12 ATP = 108 净生成: 146ATP 饱和脂酸完全氧化净生成ATP的数量:(8.5n-7)ATP (n 为偶数) 硬脂酸燃烧热值:–2651 kcal β-氧化释放:146ATP×(-7.3Kcal)=-1065.8Kcal 转换热效率
(二) Oxidation of unsaturated fatty acids(不饱和脂肪酸的β-氧化) Unsaturated fatty acids require some additional processing before they can be degraded complete.
1. 单不饱和脂肪酸的氧化——油酸的β氧化 △3顺—△2反烯脂酰CoA异构酶(改变双键位置和顺反构型)
(146-2)ATP 2. 多不饱和脂酸的氧化——亚油酸的β氧化 △3顺—△2反烯脂酰CoA异构酶(改变双键位置和顺反构型)
β-羟脂酰CoA差向酶(改变β-羟基构型:D→L型) (146—2—2)ATP
%2.4026518.1065安徽师范大学生命科学院生物专业生物化学教案 13-10-12 43. 奇数碳脂肪酸的β氧化 奇数碳脂肪酸经反复的β氧化,最后可得到丙酰CoA,丙酰CoA有两条代谢途径: (1)丙酰CoA转化成琥珀酰CoA,进入TCA。 动物体内存在这条途径,因此,在动物肝脏中奇数碳脂肪酸最终能够异生为糖。 反刍动物瘤胃中,糖异生作用十分旺盛,碳水化合物经细菌发酵可产生大量丙酸,进入宿主细胞,在硫激酶作用下产丙酰CoA,转化成琥珀酰CoA,参加糖异生作用。 (2)丙酰CoA转化成乙酰CoA,进入TCA 这条途径在植物、微生物中较普遍。 有些植物、酵母和海洋生物,体内含有奇数碳脂肪酸,经β氧化后,最后产生丙酰CoA。
(三) 脂酸的其它氧化途径 1. α—氧化(不需活化,直接氧化游离脂酸) 植物种子、叶子、动物的脑、肝细胞,每次氧化从脂酸羧基端失去一个C原子。 RCH2COOH→RCOOH+CO2
α—氧化对于降解支链脂肪酸、奇数碳脂肪酸、过分长链脂肪酸(如脑中C22、C24)
有重要作用 2. ω—氧化(ω端的甲基羟基化,氧化成醛,再氧化成酸) 动物体内多数是12C以上的羧酸,它们进行β氧化, ω—氧化涉及末端甲基的羟基化,生成一级醇,并继而氧化成醛,再转化成羧酸。 ω—氧化在脂肪烃的生物降解中有重要作用。泄漏的石油,可被细菌ω氧化,把烃转变成脂肪酸,然后经β氧化降解。
第三节 酮体代谢(Metabolism of Ketone Bodies) 脂肪酸β-氧化产生的乙酰CoA,在肌肉和肝外组织中直接进入TCA,然而在肝、肾脏细胞中还有另外一条去路:生成乙酰乙酸、D-β-羟丁酸、丙酮,这三种物质统称酮体。 酮体在肝中生成后,再运到肝外组织中利用。
一、酮体的生成(Formation of Ketone Bodies) 酮体的合成发生在肝、肾细胞的线粒体内。 形成酮体的目的是将肝中大量的乙酰CoA转移出去,乙酰乙酸占30%,β—羟丁酸70%,少量丙酮。(丙酮主要由肺呼出体外) 肝脏线粒体中的乙酰CoA走哪一条途径,主要取决于草酰乙酸的可利用性。饥饿状态下,草酰乙酸离开TCA,用于异生合成Glc。当草酰乙酸浓度很低时,只有少量乙酰CoA进入TCA,大多数乙酰CoA用于合成酮体。 当乙酰CoA不能再进入TCA时,肝脏合成酮体送至肝外组织利用,肝脏仍可继续氧化脂肪酸。 肝中酮体生成的酶类很活泼,但没有能利用酮体的酶类。因此,肝脏线粒体合成的酮体,迅速透过线粒体并进入血液循环,送至全身。
二、酮体的利用(Oxidation of Ketone Bodies) 肝外许多组织具有活性很强的利用酮体的酶。 乙酰乙酸被琥珀酰CoA转硫酶(β-酮脂酰CoA转移酶)活化成乙酰乙酰CoA 心、肾、脑、骨骼肌等的线粒体中有较高的酶活性,可活化乙酰乙酸。