数量性状的遗传(2)
- 格式:ppt
- 大小:349.50 KB
- 文档页数:17





第五章 数量性状的遗传畜禽的大多数经济性状属于数量性状。
掌握数量性状的遗传规律和遗传参数对种畜生产中种畜群的生产性能的保持、对地方品种经济性能的提高、对新品种新品系的培育等工作都是十分必要的。
数量性状的遗传是有规律所循的,虽然在不同群体、在不同条件下、因估计方法不同,得到的参数有所变化,但遗传参数反映的数量性状的基本遗传规律的趋势是一定的。
第一节 数量性状的遗传基础质量性状的变异一般遵从孟德尔遗传规律,但数量性状的遗传规律与质量性状的遗传规律有一定区别。
数量性状是由大量的、效应微小而类似的、可加的基因控制,呈现连续变异,数量性状的表现还受到大量复杂环境因素的影响。
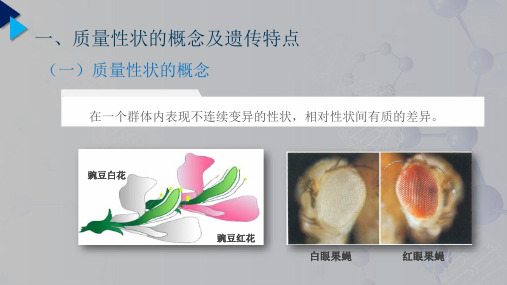
一、Nilsson-Ehle 假说及其发展生物的性状按照其表现和对其研究的方式,可大致分为质量性状、数量性状和阈性状。
质量性状的变异通常可以区分为几种明显不同的类型,遵从孟德尔遗传规律。
畜禽重要质量性状的遗传规律已经在上一章中进行了阐述。
在动物生产中所关注的绝大多数经济性状呈连续性变异,其在个体间表现的差异只能用数量来区分,这类性状称为数量性状,如奶牛的产奶量、鸡的产蛋量、肉用家畜的日增重、饲料转化率、羊的产毛量等。
与质量性状相比较,数量性状主要有以下特点:①性状变异程度可以用度量衡度量;②性状表现为连续性分布;③性状的表现易受到环境的影响;④控制性状的遗传基础为多基因系统。
遗传基础为多基因控制,而表现为非连续性变异的性状称为阈性状。
如羊的产羔数、肉质的分类、对疾病抗性的有无等。
严格说来,鸡的产蛋数、猪的窝产仔数等也属于这一类性状,但其表型状态过多,作为阈性状分析过于复杂,通常近似的将其作为数量性状来看待。
数量性状在畜牧生产中占有非常重要的地位。
但是,到目前为止,对数量性状的遗传基础的解释主要还是基于Yule (1902,1906)首次提出、由Nilsson-Ehle (1908)总结完善、并由Johannsen (1909)和East (1910)等补充发展的多因子假说,也称为多基因假说或Nilsson-Ehle 假说。