第四章数量性状的遗传
- 格式:doc
- 大小:163.00 KB
- 文档页数:5




第五章数量性状§1 数量性状的特征一、数量性状的特征:遗传性状分为质量性状和数量性状两类。
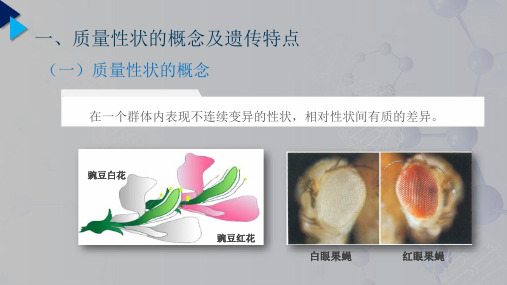
质量性状:在可以遗传的性状中,性状在后代的变异中是表现为不连续的变异,在它们之间可以明显的分组,进行频率的统计。
是由一对或少数n对基因所控制的遗传方式一般能符合孟德尔的遗传原则。
例如前几章所讲的豌豆的红花和白花,豆粒的黄色和绿色,家鸡的羽毛的黑白、芦花等,都是彼此差别明显,一般没有中间过渡的类型的性状。
数量性状:性状的变异是呈现连续性的,性状间的变化没有明显的类别,不易分组,是由微效的多基因控制的。
例如:产量,荚的多少、粒的大小,蛋白、脂肪的含量等。
数量性状的特点:1.性状的变化表现为连续的,不易分组进行组内的频率统计。
例如:水和小麦、玉米等植株的高矮。
生育期的长短,产量的高低等性状。
不同品种杂交后F2、F3等后代群体都有广泛的变异的类型,不能明显的求出分离的比例,只能用一定的度量单位进行测量。
2.数量性状极易受环境的影响而发生混淆,使遗传的动态和性质模糊不清。
二、数量性状在遗传中的特点:而数量性状在遗传中的特点又是怎样呢?我们以玉米果穗不同长度的品种的杂交及后代的分布频率来说明此题:P2页表从上面这个典型的数量性状遗传的例子中,可以看出数量性状遗传的主要的特点:1.某一数量性状在杂交中,F1的平均值大约介于两个亲本的平均数之间,呈中间型,但有时倾向于其中的一个亲本。
2.F2个体的平均数与F1的平均数相似。
3.F2出现明显的连续性变异,不容易分组,因而也就不能求出不同组之间的比例,变异的幅度比较大,变异基本上是以平均数为中心的对称的正态分布曲线,即常态分布。
100cm(A1A1A2A2a3a3) × 70cm(a1a1a2a2A3A3)↓F1 A1a1A2a2A3a3 (85cm)↓F2 A1A1A2A2A3A3(130cm):a1a1a2a2a3a3(40cm)4.F2会出现超出双亲的变异的类型:<当双亲不是极端类型时>100天 A×B 80天↓F1 90天→F2:75~110天值得说明的是:数量性状和质量性状之间的划分不是绝对的,同一性状在不同亲本的杂交组合中可以表现不同。

第四章数量性状的遗传目的要求掌握数量性状与质量性状的区分、特征,多基因假说的要点,数量性状表现值的分解,遗传力的概念;了解通径系数概念与意义,基因的非加性效应与加性效应的意义,遗传力公式的推导及计算方法;掌握遗传力的应用。
第一节数量性状的遗传基础生物的性状基本上可分为两大类:质量性状(qualitative trait):变异可以截然区分为几种明显不同的类型,一般用语言来描述;数量性状(quantitative trait):个体间性状表现的差异只能用数量来区别,变异是连续的。
阈性状(threshold trait):表现型呈非连续变异,与质量性状类似,但不是由单基因决定,性状具有一个潜在的连续型变量分布,遗传基础是多基因控制的,与数量性状类似。
一、数量性状的一般特征数量性状的特点:①数量性状是可以度量的;②数量性状呈连续性变异;③数量性状的表现容易受到环境的影响;④控制数量性状的遗传基础是多基因系统。
学习数量性状的方法①统计学思想贯穿数量性状遗传的全部内容;②确定性与不确定性的矛盾时时体现;③研究对象在个体与群体间的相互转换;④遗传与变异的矛盾。
二、数量性状的遗传基础1.多基因假说瑞典遗传学家尼尔迩·埃尔(Nilsson-Ehle)通过对小麦籽粒颜色的遗传研究,提出了数量性状遗传的多基因假说。
多基因假说的要点(1)数量性状是由许多微效基因决定的,每个基因的作用的微效的;(2)基因的作用是相等的,且可以累加、呈现剂量效应,等位基因间通常无显隐关系;(3)基因在世代相传中服从孟德尔定律,即分离规律和自由组合规律,以及连锁交换规律2.基因的非加性效应基因的非加性效应包括显性效应和上位效应。
(1)显性效应由等位基因间相互作用产生的效应。
例1:有两对基因,A1、A2的效应各为20cm,a1、a2的效应名为10cm,基因型A1A1a2a2按加性效应计算其总效应为60cm 。
而在杂合状态下,即A 1a 1A 2a 2同样为两个A 和两个a ,其总效应可能是75cm(2)上位效应或互作效应 由非等位基因之间相互作用产生的效应。
数量性状的遗传名词解释数量性状,是指在自然界或人工条件下产生的各种特征以数量的方式表现出来的遗传性状。
它指的是通过对种群中大量个体进行测量或计量,将结果以数量化的形式呈现出来的遗传特征。
数量性状通常具有连续变异的特征,即在一个种群中存在着一系列不同的表现形式,而不是像离散性状那样只有几个确定的表型。
在数量性状的研究中,有一些重要的遗传名词需要加以解释。
其中包括基因型、表型、遗传方差、环境方差、遗传相关等。
基因型是指个体在基因水平上的遗传组成。
它决定了个体对特定数量性状的表现。
每个数量性状通常由多个基因共同决定,因此基因型的组合将决定这些基因在个体上的表现形式。
表型是指个体在外部表现上的特征。
它受到基因型和环境的共同影响。
数量性状的表型通常呈现连续性变化,这是因为数量性状通常受到多种基因的共同作用,以及环境因素的影响。
例如,人体身高就是一种典型的数量性状,它受到多基因的影响,同时还受到营养、运动等环境因素的调节。
遗传方差是指数量性状中由基因所引起的表型变异程度。
它可以通过研究个体间的表型差异以及表型与基因型之间的关系来估计。
遗传方差的大小反映了数量性状中遗传因素的重要程度。
如果遗传方差较大,说明遗传因素在数量性状的表达中起到了重要作用,反之则说明环境因素的贡献较大。
环境方差是指数量性状中由环境因素所引起的表型变异程度。
环境方差通常通过比较同一种群中不同个体之间的差异来估计。
环境方差的大小表示了环境对数量性状的影响程度。
如果环境方差较大,说明环境因素在数量性状的表达中起到了重要作用,反之则说明遗传因素的贡献较大。
遗传相关是指在同一种群中不同数量性状之间的遗传联系。
它反映了一种或多种数量性状随着基因型的变化而变化的程度。
通过研究数量性状之间的遗传相关,可以了解不同数量性状之间的遗传关系及其对进化和适应的影响。
例如,身高和体重之间的遗传相关可以帮助我们理解这两个数量性状在人类进化中是如何相互影响的。
以上介绍了数量性状的遗传名词解释,包括基因型、表型、遗传方差、环境方差和遗传相关等概念。
第四章数量性状的遗传
目的要求
掌握数量性状与质量性状的区分、特征,多基因假说的要点,数量性状表现值的分解,遗传力的概念;了解通径系数概念与意义,基因的非加性效应与加性效应的意义,遗传力公式的推导及计算方法;掌握遗传力的应用。
第一节数量性状的遗传基础
生物的性状基本上可分为两大类:
质量性状(qualitative trait):变异可以截然区分为几种明显不同的类型,一般用语言来描述;
数量性状(quantitative trait):个体间性状表现的差异只能用数量来区别,变异是连续的。
阈性状(threshold trait):表现型呈非连续变异,与质量性状类似,但不是由单基因决定,性状具有一个潜在的连续型变量分布,遗传基础是多基因控制的,与数量性状类似。
一、数量性状的一般特征
数量性状的特点:
①数量性状是可以度量的;
②数量性状呈连续性变异;
③数量性状的表现容易受到环境的影响;
④控制数量性状的遗传基础是多基因系统。
学习数量性状的方法
①统计学思想贯穿数量性状遗传的全部内容;
②确定性与不确定性的矛盾时时体现;
③研究对象在个体与群体间的相互转换;
④遗传与变异的矛盾。
二、数量性状的遗传基础
1.多基因假说
瑞典遗传学家尼尔迩·埃尔(Nilsson-Ehle)通过对小麦籽粒颜色的遗传研究,提出了数量性状遗传的多基因假说。
多基因假说的要点
(1)数量性状是由许多微效基因决定的,每个基因的作用的微效的;
(2)基因的作用是相等的,且可以累加、呈现剂量效应,等位基因间通常无显隐关系;(3)基因在世代相传中服从孟德尔定律,即分离规律和自由组合规律,以及连锁交换规律2.基因的非加性效应
基因的非加性效应包括显性效应和上位效应。
(1)显性效应由等位基因间相互作用产生的效应。
例1:有两对基因,A1、A2的效应各为20cm,a1、a2的效应名为10cm,基因型A1A1a2a2
按加性效应计算其总效应为60cm 。
而在杂合状态下,即A 1a 1A 2a 2同样为两个A 和两个a ,其总效应可能是75cm
(2)上位效应或互作效应 由非等位基因之间相互作用产生的效应。
例2:上例中A 1A 1A 2A 2按加性效应应是80cm ,但实际总效应可能是90cm
3.数量性状基因座
对数量性状有较大影响的基因座称为数量性状基因座(quantitative trait locus ,QTL),它是影响数量性状的一个染色体片段,而不一定是一个单基因座。
三、通径系数理论
通径系数就是标准化的偏回归系数 。
通径系数的主要优点是能够借图解之助简明地阐明各变量之间的关系。
两个有关变量之间可有两种关系,一是平行关系,一是因果关系。
具有平行关系的双方不分因果,只是由于两者具有共同原因,或互为因果,因而彼此相关。
X1Y X2X1Y X3
X2
单箭头线叫做通径,双箭头线叫做相关线。
用一个数字来表明每条线的相对重要性,这种数字叫做系数。
相关线的系数就是相关系数,通径的系数叫做通径系数。
通径系数就是标准化回归系数,在多自变量情况下,就是标准化偏回归系数。
Sb y ·x =P y ·x =b y ·x σx /σy
箭头线叫做通径,双箭头线叫做相关线。
用一个数字来表明每条线的相对重要性,这种数字叫做系数。
相关线的系数就是相关系数,通径的系数叫做通径系数。
回归系数b yx =SP/SS X
相关系数
第二节 数量性状的遗传力
一、遗传力的概念
(一)表型值的剖分
P=G+E
P=G+E=A+D+I+E=A+R
R=D+I+E
V p =V G +V E
遗传力:是遗传方差在总表型方差中所占的比率 y x xy xy SS SS SP r =A I D A G E G P =++==+=
H 22称为广义遗传力
狭义遗传力(h 2)
遗传力估计值可以用百分数或者小数来表示
1.数量性状的遗传力估计值介于0~1之间。
2.根据性状遗传力值的大小,可将其大致划分为三等,即0.5以上者为高遗传力;0.2以下者为低遗传力。
3.遗传力估计值只是说明对后代群体某性状的变异来说,遗传与环境两类原因影响的相对重要性。
并不是指该性状能遗传给后代个体的绝对值。
二、遗传力的估算方法
1.由亲仔关系估算遗传力
在家畜育种中,一般用女母回归法来估算遗传力。
h 2=2b op
P 是母亲某性状的表型值,O 是女儿该性状的表型值。
=母亲的变量
在畜群中母畜数大大超过公畜数,所以一般都利用母畜中的子亲回归,即女母回归(b o p )。
为了消除各公畜对畜群影响大小的差异(各公畜的女儿不等),在家畜育种中常以公畜内女母回归(b w (o p ))表示全群的女母回归。
公畜内女母回归和遗传力的计算步骤:
①整理资料。
以女儿的个体表型值为O ,以其母亲的个体表型值为P ;
②分别计算各公畜组的平方和SS p 、SS 0、乘积和SP op ;
③将各组的上述数据总加起来,得到公畜内平方和、公畜内乘积和;
P G V V H =2P
A V V h =22p op op Cov b σ=
N P P O O
Cov op ∑--=))((N P P p ∑-=22)(σ
N AR A N R A A Cov op ∑∑∑+=+=2121)(212∑=0AR A op V Cov 21=22121h V V b P A op ==op
b h 22=∑=P P W SS SS )(∑=OP OP W SP SP )(∑=O O W SS SS )(
计算公畜内女母回归和遗传力:
h 2=2b w(op)
2.半同胞相关法
半同胞就是同父异母或同母异父的兄弟姐妹。
通过对若干公畜(或母畜)的女儿某性状表型
值的产量,用方差分析法可求得组间(公畜或母畜间)方差和组内方差。
h 2=4r H S
组内相关系数即是半同胞相关系数,用r HS 表示
M S S 为公畜间均方,M S w 为公畜内均方,n 为各公畜的女儿数。
利用上式计算出半同胞组内相关系数后,再乘上4,即为该性状的遗传力。
半同胞相关和遗传力的计算步骤如下:
①整理资料。
②分别计算出各组的∑X ,∑X 2和
然后将各种的上述数据总加起来,得到∑∑∑X 、∑∑X 2、∑C X 和∑n ;
③计算平方和、自由度和均方:
S S w =∑∑X 2-∑C X
d f S =S -1
d f W =∑n -S
④ 计算各组的平均同胞数n 0。
⑤计算半同胞相关和遗传力:
h 2=4r HS
3.利用“全同胞—半同胞”混合家系计算遗传力
各公畜加权平均子女数
)()()(p w op w op w SS SP b =
222w S S HS r σσσ+=W S W S HS MS n MS MS MS r )1(-+-=⎪⎪⎭⎫ ⎝⎛--=∑∑∑i i i n n n S n 2011()n X C X 2∑=()∑∑∑∑-=n X C SS X S 2S S S df SS MS =W W W df SS MS =W S W S HS MS n MS MS MS r )1(0-+-=⎪⎪⎭⎫ ⎝⎛-=∑∑
∑i i i S n n n df n 201
N 为子女总数,n i 为每头母畜子女数,Σn i 为每头公畜子女数 遗传力为:
三、遗传力的应用
1.估计种畜的育种值
2.确定繁育方法
3.确定选择方法
4.应用于综合选择指数的制订
思考题:
1.质量性状与数量性状各有何特点?如何区分?
2.多基因假说的要点是什么?
3.数量性状的遗传机制是什么?
4.如何正确剖分数量性状的表现值?
5.什么是遗传力?如何估算数量性状的遗传力?估算的方法有哪些?
6.数量性状的遗传力有何应用价值? W
S W S I MS n MS MS MS r )1(0-+-=⎥⎥⎦
⎤⎢⎢⎣⎡--+=∑∑∑∑N n N n r i i A 22)(125.0A I r r h =2。