小鼠海马区神经元的培养
- 格式:doc
- 大小:33.00 KB
- 文档页数:2



电针通过调节Notch3信号通路改善大脑中动脉闭塞小鼠海马神经元凋亡的作用机制杨语晗,许明军,穆敬平摘要 目的:探讨电针通过调节Notch3信号通路改善大脑中动脉闭塞小鼠海马神经元凋亡的作用机制㊂方法:将45只健康无特定病原体(SPF )级雄性C57BL/6小鼠随机分为对照组㊁大脑中动脉闭塞模型(MCAO )组和电针治疗组,各15只㊂通过2,3,5-氯化三苯基四氮唑(TTC )染色和小鼠神经行为评分比较小鼠脑梗死神经损伤情况㊂采用免疫组织化学分析小鼠海马神经元Bax 和Caspase -3蛋白表达㊂通过末端脱氧核苷酸转移酶介导的dUTP 缺口末端标记(TUNEL )荧光染色分析小鼠神经元细胞凋亡率;通过实时荧光定量聚合酶链式反应(RT -PCR )分析小鼠海马组织肿瘤坏死因子-α(TNF -α)㊁白细胞介素(IL )-1β㊁IL -6㊁诱导型一氧化氮合酶(iNOS )mRNA 表达;通过免疫组织化学分析小鼠海马神经元细胞凋亡;通过蛋白印迹分析小鼠海马组织Bax ㊁Bcl -2㊁Caspase -3及H19/EZH2/Notch3蛋白表达㊂通过免疫荧光染色分析小鼠脊髓组织Nestin 和神经胶质纤维酸性蛋白(GFAP )表达㊂结果:连续干预14d 后,MCAO 组小鼠神经损伤和脑梗死体积较对照组增加(P <0.05),电针治疗组小鼠神经损伤和脑梗死体积较MCAO 组减小(P <0.05)㊂MCAO 组神经细胞凋亡率较对照组升高(P <0.05),电针治疗组神经元细胞凋亡率较MCAO 组减少(P <0.05);MCAO 组TNF -α㊁IL -1β㊁IL -6㊁iNOS mRNA 表达较对照组升高(P <0.05),电针治疗组TNF -α㊁IL -1β㊁IL -6㊁iNOS mRNA 表达较MCAO 组降低(P <0.05)㊂MCAO 组Bax 和Caspase -3蛋白表达较对照组升高(P <0.05),电针治疗组Bax 和Caspase -3蛋白表达较MCAO 组降低(P <0.05),MCAO 组Bcl -2蛋白表达较对照组降低(P <0.05),电针治疗组Bcl -2蛋白表达较MCAO 组升高(P <0.05)㊂MCAO 组H19㊁EZH2和Notch3蛋白表达较对照组升高(P <0.05),电针治疗组H19㊁EZH2和Notch3蛋白表达较MCAO 组降低(P <0.05)㊂MCAO 组Nestin 蛋白表达较对照组降低(P <0.05),GFAP 蛋白表达较对照组升高(P <0.05);电针治疗组Nestin 蛋白表达较MCAO 组升高(P <0.05),GFAP 蛋白表达较MCAO 组降低(P <0.05)㊂结论:MCAO 可诱导海马神经元凋亡,电针通过调控Notch3激活,调节Bax/Bcl -2及Nestin 和GFAP 蛋白,从而阻断Caspase -3激活,抑制神经元凋亡㊂关键词 大脑中动脉闭塞;Notch3信号通路;电针;海马神经元;实验研究d o i :10.12102/j.i s s n .1672-1349.2023.24.010 脑卒中是全球第二大死亡原因和第三大残疾原因[1-2]㊂据估计,到2030年,中风每年将导致1200万例病人死亡,缺血性脑卒中主要由大脑中动脉脑血流阻塞引起,约占所有脑卒中的87.9%,导致严重的中枢神经系统损伤甚至死亡[3-4]㊂缺血性脑卒中是世界范围内死亡和残疾的主要原因[5-6]㊂缺血性脑卒中后血脑屏障受损导致损伤进一步发展和扩大㊂由于缺血通过剥夺氧气和代谢底物扰乱脑组织,因此在缺血条件下细胞分泌有害信号[7-9]㊂这些有害分子促进小胶质细胞活化,通过释放促炎调节剂[包括肿瘤坏死因子-α(TNF -α)㊁白细胞介素(IL )-1β㊁IL -6]引起促炎反应,从而导致炎症诱导的坏死性神经元细胞死亡㊂神经元坏百会穴死和神经元与小胶质细胞的破坏进一步增强炎症反应㊂坏死性凋亡是一种受调节的细胞死亡形式,对缺血性脑卒中损伤的进展有影响㊂抑制坏死性凋亡可显著减小梗死面积,抑制炎症,减轻运动能力,并最终改善脑缺血性损伤后认知功能㊂Notch 信号通路是一种进化上相对保守的信号通路,参与海马神经元细基金项目 湖北省卫生健康委中医药科研项目(No.ZY2021M006)作者单位 十堰市太和医院,湖北医药学院附属医院(湖北十堰442000)通讯作者 穆敬平,E -mail :****************引用信息 杨语晗,许明军,穆敬平.电针通过调节Notch3信号通路改善大脑中动脉闭塞小鼠海马神经元凋亡的作用机制[J ].中西医结合心脑血管病杂志,2023,21(24):4530-4535.胞迁移㊁分化和凋亡㊂Notch 信号通路在一些人类疾病中被激活,包括动脉粥样硬化㊁结肠直肠癌和缺血性脑卒中㊂有研究显示,抑制Notch 信号通路可能通过诱导神经元干分化并减少神经炎症和神经元凋亡[10]㊂针灸长期用于疾病预防,一项研究支持其在动物模型中诱导脑缺血耐受作用的证据[11],结果显示,在脑缺血前于百会穴(GV20)进行电针可减轻动物模型中缺血后神经功能缺损程度㊂目前,已从抑制炎症㊁抑制氧化应激㊁抑制内质网应激㊁调节大麻素系统㊁自噬㊁保护血脑屏障㊁抗凋亡等多方面探讨了电针预处理诱导脑缺血耐受的机制㊂多项研究表明,电针可抑制神经元凋亡,改善神经元微环境,加速水肿减轻和神经功能修复;抑制促炎细胞因子和Notch 信号通路相关因子的表达,表明抑制电针诱导的Notch 信号通路有助于缓解缺血性脑卒中[12-13]㊂本研究旨在探讨电针通过调节Notch3信号通路改善大脑中动脉闭塞(middle cerebral artery occlusion ,MCAO )小鼠海马神经元凋亡的机制,为脑梗死等脑血管疾病类型提供新的治疗方向㊂1 材料与方法1.1 实验动物45只健康无特定病原体(SPF )级雄性C57BL/6小鼠,体质量20~25g ,购自北京维通利华实验动物科技有限公司㊂小鼠生存温度(23.0ʃ2.0)ħ,环境湿度45%~50%,光照时间每日12h ;适应期间可随意获取水和适应性饲料㊂1.1.1小鼠MCAO模型的建立MCAO模型采用改良的Zea Longa方法建立㊂腹腔注射10%水合氯醛300mg/kg后,将小鼠仰卧固定于37ħ恒温电加热板上经颈正中切口分离右侧颈总动脉㊁颈外动脉和颈内动脉,结扎颈总动脉近端㊂使用无创血管夹住颈总动脉远端和颈外动脉㊂在颈总动脉分叉处做一切口,放置头部钝化的尼龙线㊂将尼龙线缓慢向前推8~10mm㊂当感觉到阻力时,停止并固定在颈总动脉上㊂栓塞1h后,拔出线塞恢复血流㊂再灌注2h后,使用神经功能评分验证模型是否成功㊂1.1.2动物分组将45只小鼠随机分为3组,每组15只,对照组除不将细丝推进至大脑中动脉起点处外,其他处理与MCAO组相同;MCAO组建立小鼠MCAO模型;电针治疗组:MCAO小鼠模型建立后,百会穴位于矢状中线与两耳连线的交点处,使用XYD-IV仪器(翔宇医疗,型号20152260115),以1mA的强度和2~15Hz的频率刺激,每日20min,连续14d㊂1.2实验方法1.2.1小鼠神经损伤评估神经行为评分包括6方面:自发活动(笼中5min, 0~3分)㊁运动对称性(四肢,0~3分)㊁前肢对称性(尾握时伸展,0~3分)㊁铁丝笼爬墙(1~3分)㊁躯干两侧的触摸反应(1~3分)和触须触摸的反应(1~3分)㊂总分18分,得分越低提示神经功能受损程度越小,计算脑梗死体积㊂1.2.22,3,5-氯化三苯基四氮唑(TTC)染色计算脑梗死体积将小鼠麻醉并处死后提取脑组织,使用冰盐水冲洗后,在-20ħ冰箱中快速冷冻脑组织20min,使脑组织变硬进行切割㊂沿冠状面切片,厚度为2mm,每个脑切5片或6片㊂将脑切片放入用锡纸包裹并置于含有2%TTC(北京索莱宝公司)的培养皿中,在37ħ培养箱中染色30min㊂在此期间,轻轻转动大脑切片,使其充分暴露于反应液中㊂取出培养皿,用1mL针管吸出TTC溶液,将切片置于4%多聚甲醛溶液中固定5min,取出切片按顺序摆放㊂采用Image-Pro Plus图像软件计算每个脑切片的脑梗死体积比㊂为减少脑缺血性半球水肿引起的误差,梗死体积为对侧正常脑组织体积-同侧正常组织体积,结果以梗死体积百分比表示:梗死面积体积百分比=(对侧正常脑组织体积-同侧正常脑组织体积)/对侧正常组织体积ˑ100%㊂1.2.3末端脱氧核苷酸转移酶介导的dUTP缺口末端标记(TUNEL)荧光染色检测细胞凋亡率小鼠脑组织经二甲苯脱蜡㊁乙醇脱水后,石蜡切片加入20μg/mL蛋白酶K工作液(北京索莱宝公司),室温放置15min㊂向每个样品中加入51μL的TUNEL 溶液(45μL平衡缓冲液ʒ5μL核苷酸混合物ʒ1μL rTdT酶),避光孵育1h㊂加入2ˑSSC缓冲液在室温下孵育15min以终止反应㊂加入含有抗荧光淬灭的4',6-二脒基-2-苯基吲哚(DAPI),黑暗中孵育5min,盖上盖玻片㊂通过荧光显微镜(倒置荧光显微镜,德国)获得海马CA1区图像(200倍)㊂计算不同视野内凋亡阳性细胞百分比,取平均值进行统计分析㊂凋亡率=阳性细胞/细胞总量ˑ100%㊂1.2.4免疫组织化学分析小鼠海马神经元细胞Bcl-2㊁Bax和Caspase-3蛋白表达组织包埋,切片,烘烤,脱蜡,补液和抗原修复后,在37ħ条件下,将细胞在2%的低聚甲醛中固定4h,在封闭缓冲液[含10%山羊血清和0.01%Tween-20的磷酸缓冲盐溶液(PBS)]中孵育1h㊂37ħ条件下,在封闭缓冲液中将组织与第一抗体[兔鼠抗小鼠Bcl-2多克隆抗体(1ʒ200)㊁兔鼠抗小鼠Bax多克隆抗体(1ʒ200)㊁兔抗小鼠Caspase-3多克隆抗体(1ʒ200)] 4ħ过夜㊂将组织用含0.05%Tween-20的PBS洗涤3次,在室温下添加1ʒ200的山羊抗小鼠二抗(Aspen 公司),在37ħ条件下,细胞孵育4h㊂使用PBS洗涤切片3次,加入显色剂(DAB)㊂最后,将切片冲洗, DAPI复染密封㊂在Zeiss LSM710共聚焦显微镜下观察,使用Zen2.31SP(黑色版)软件(Zeiss)处理图像㊂1.2.5实时荧光定量聚合酶链式反应检测TNF-α㊁IL-1β㊁IL-6㊁诱导型一氧化氮合酶(iNOS)mRNA表达收集小鼠的海马组织,在1mL TRIzol Reagent (15596026,赛默飞世尔科技公司,美国)中研磨成组织匀浆,转移到1.5mL离心管中,加入200μL氯仿,混合并在4ħ㊁10000r/min下离心5min以获得总mRNA(在上层水相中)㊂加入500μL异丙醇,混匀,在相同条件下再次离心,纯化mRNA㊂弃上清,采用75%乙醇洗涤异丙醇,得到离心管底部富集的mRNA 沉淀㊂加入50μL焦碳酸二乙酯(DEPC)水溶解mRNA,用Nano Drop微管分光光度计(Invitrogen公司,美国)测量mRNA浓度㊂根据试剂盒说明,使用Transcriptor First Strand cDNA Synthesis Kit (***********,罗氏公司,德国)将mRNA逆转录为cDNA㊂通过实时PCR系统(LightCycler 480,罗氏公司,德国)检测每个样品TNF-α㊁IL-1β㊁IL-6㊁iNOS mRNA表达水平㊂1.2.6蛋白免疫印迹法检测海马H19㊁EZH2和Notch3蛋白表达水平1)提取蛋白:人卵巢癌细胞,加入10mL/mg蛋白免疫印迹法及IP细胞裂解液,使用匀浆器匀浆,利用超声组织破碎仪破碎细胞(破碎条件为每管4次/300W),细胞破碎后冰上放置30min,将破碎后的细胞进行离心, 4ħ㊁12000ˑg(重力加速度)离心30min,收集上清液,即为蛋白抽提液㊂2)蛋白定量:使用二喹啉甲酸(BCA)法测定蛋白浓度,将蛋白浓度调节一致㊂3)凝胶电泳:取待测标本15μL进行上样,该过程中选取甘油醛-3-磷酸脱氢酶(glyceraldehyde-3-phosphate dehydrogenase,GAPDH)单抗(1ʒ500)作为蛋白质上样量的标定物,每泳道上样30μg蛋白量㊂使用十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate-polyacrylamide gel electroresis,SDS-PAGE)进行电泳分析,至目的分子量出现㊂4)湿法转膜:采用湿法将蛋白条带电转至Immun-blot聚偏乙烯膜(PVDF)上,加入浓度为50g/L的脱脂奶粉在20ħ条件下封闭震荡处理3h,加入以下抗体[兔鼠抗小鼠H19多克隆抗体(1ʒ400)㊁兔抗小鼠EZH2(1ʒ400)多克隆抗体㊁兔抗小鼠Notch3多克隆抗体(1ʒ400)],4ħ孵育过夜,用TBS缓冲液(Tris50mmol/L,NaCl100 mmol/L,pH7.5)冲洗3次,每次10min㊂使用经过辣根酶标记的IgG(1ʒ1000),在37ħ条件下孵育2h,漂洗3次,每次10min㊂5)发光显影:使用电化学发光(ECL)试剂盒进行显影操作㊂首先等体积混合ECL 试剂盒中的A液和B液,20ħ保存备用,将二抗加入于反应液中进行孵育,使用PBS进行多次洗涤,取出膜后使用洁净的吸水纸轻轻吸去多余的液体,将膜放置在洁净的保鲜膜上,整个操作过程需要谨慎小心,避免触碰蛋白电泳一侧㊂按照0.125mL/cm2滴加发光液,使发光液均匀覆盖在膜上静置5min,取出膜,去除上面的发光液㊂将膜固定在两片保鲜膜中间,于暗室内压片5min后完成显影定影操作㊂选择GAPDH单抗(1ʒ1000)作为内参㊂6)图像分析:胶片干燥后,使用Mieroteck扫描仪扫描图像,利用Quantity-One软件进行积分光密度分析㊂1.2.7免疫荧光染色法检测MCAO和电针治疗小鼠脊髓组织中Nestin和神经胶质纤维酸性蛋白(GFAP)的表达取10μm厚的脊髓组织切片在4ħ下在4%多聚甲醛中固定㊂将脊髓组织与封闭溶液(0.2%T riton X-100)和5%牛血清白蛋白(V/V)在室温下孵育1h,在4ħ下与相应的溶液孵育过夜㊂针对胶质纤维酸性蛋白(GFAP1ʒ1000,Abcam公司,美国)和巢蛋白(1ʒ200,Abcam公司,美国)的特异性抗体,冲洗后,将切片与Alexa Fluor555(1ʒ500;Abcam公司,美国)抗兔IgG 作用1h㊂用4',6-diamidino-2-phenylindole(DAPI)复染细胞核㊂采用Nikon Eclipse80i显微镜(Nikon公司,日本)捕获荧光图像㊂1.3统计学处理采用SPSS20.0统计软件(美国IBM公司)对数据进行分析,符合正态分布的定量资料以均数ʃ标准差(xʃs)表示,多组间比较采用单因素方差分析,两组间比较采用最小显著差异法(LSD-t)㊂以P<0.05为差异有统计学意义㊂2结果2.1各组小鼠脑神经损伤得分和脑梗死体积比较MCAO组小鼠神经损伤和脑梗死体积较对照组增加(P<0.05),电针治疗组小鼠神经损伤和脑梗死体积较MCAO组减少(P<0.05)㊂详见表1㊂表1各组小鼠神经损伤得分和脑梗死体积比较(xʃs)组别只数神经损伤得分(分)脑梗死体积(%)对照组15 2.64ʃ0.52 1.21ʃ0.36 MCAO组1515.39ʃ2.17①37.52ʃ4.24①电针治疗组15 6.26ʃ1.33②13.53ʃ2.17②F值12.77316.572P<0.001<0.001注:MCAO组与对照组比较,①P<0.05;电针治疗组与MCAO组比较,②P<0.05㊂2.2各组小鼠神经细胞凋亡率比较MCAO组神经细胞凋亡率较对照组升高(P<0.05),电针治疗组神经细胞凋亡率较MCAO组降低(P< 0.05)㊂详见表2㊂表2各组小鼠神经细胞凋亡率比较(xʃs)组别只数神经细胞凋亡率(%)对照组157.25ʃ3.42 MCAO组1545.52ʃ8.69①电针治疗组1518.34ʃ3.66②F值13.527P<0.001注:MCAO组与对照组比较,①P<0.05;电针治疗组与MCAO组比较,②P<0.05㊂2.3各组炎性因子mRNA表达比较MCAO组TNF-α㊁IL-1β㊁IL-6㊁iNOS mRNA表达较对照组升高(P<0.05),电针治疗组TNF-α㊁IL-1β㊁IL-6㊁iNOS mRNA表达较MCAO组降低(P<0.05)㊂详见表3㊂表3各组炎性因子mRNA表达比较(xʃs)组别只数TNF-αmRNA IL-1βmRNA IL-6mRNA iNOS mRNA 对照组15 1.02ʃ0.02 1.04ʃ0.01 1.00ʃ0.01 1.01ʃ0.01 MCAO组15 1.86ʃ0.15① 1.74ʃ0.11① 1.85ʃ0.16① 1.92ʃ0.16①电针治疗组15 1.24ʃ0.08② 1.13ʃ0.04② 1.07ʃ0.05② 1.16ʃ0.09②F值15.77312.62511.42412.702 P<0.001<0.001<0.001<0.001注:MCAO组与对照组比较,①P<0.05;电针治疗组与MCAO组比较,②P<0.05㊂2.4电针治疗抑制MCAO小鼠海马神经元细胞Bax㊁Bcl-2和Caspase-3蛋白表达通过免疫组织化学分析小鼠海马神经元细胞凋亡,MCAO组Bax和Caspase-3蛋白表达较对照组升高(P<0.05),电针治疗组Bax和Caspase-3蛋白表达较MCAO组降低(P<0.05);MCAO组Bcl-2蛋白表达较对照组降低(P<0.05),电针治疗组Bcl-2蛋白表达较MCAO组升高(P<0.05)㊂详见图1㊁表4㊂图1各组Bax㊁Bcl-2和Caspase-3蛋白表达免疫组织化学分析表4各组Bax㊁Bcl-2和Caspase-3蛋白表达比较(xʃs)组别只数Bax蛋白Bcl-2蛋白Caspase-3蛋白对照组15 1.12ʃ0.03 1.95ʃ0.18 1.07ʃ0.04 MCAO组15 1.86ʃ0.16① 1.13ʃ0.10① 1.85ʃ0.13①电针治疗组15 1.08ʃ0.02② 1.82ʃ0.16② 1.14ʃ0.06①②F值10.63511.2539.557P0.0030.0120.026注:MCAO组与对照组比较,①P<0.05;电针治疗组与MCAO组比较,②P<0.05㊂2.5电针治疗对H19/EZH2/Notch3信号通路的影响MCAO组H19㊁EZH2和Notch3蛋白表达较对照组升高(P<0.05),电针治疗组H19㊁EZH2和Notch3蛋白表达较MCAO组降低(P<0.05)㊂详见表5㊁图2㊂表5各组H19/EZH2/Notch3信号通路相关蛋白表达比较(xʃs)组别只数H19蛋白EZH2蛋白Notch3蛋白对照组15 1.02ʃ0.01 1.05ʃ0.04 1.03ʃ0.02 MCAO组15 1.87ʃ0.15① 1.92ʃ0.17① 1.95ʃ0.14①电针治疗组15 1.06ʃ0.03② 1.12ʃ0.08② 1.09ʃ0.07②F值10.72112.3358.897P0.0120.0040.011注:MCAO组与对照组比较,①P<0.05;电针治疗组与MCAO组比较,②P<0.05㊂图2各组相关蛋白表达条带图2.6电针治疗对脊髓组织中神经干细胞Nestin和GFAP蛋白表达的影响MCAO组Nestin蛋白表达较对照组降低(P<0.05), GFAP蛋白表达较对照组升高(P<0.05);电针治疗组Nestin蛋白表达较MCAO组升高(P<0.05),GFAP 蛋白表达较MCAO组降低(P<0.05)㊂免疫荧光染色检测MCAO和电针治疗小鼠脊髓组织中Nestin和GFAP蛋白表达,红色代表GFAP阳性,绿色代表Nestin阳性,免疫荧光染色结果显示,与MCAO组比较,电针治疗组Nestin蛋白水平增加,GFAP蛋白水平降低㊂表明电针治疗显著增加神经元干细胞增殖并抑制向星形胶质细胞分化㊂详见图3㊁表6㊂图3各组神经干细胞免疫荧光染色图表6各组神经干细胞Nestin和GFAP蛋白表达比较(xʃs)组别只数Nestin蛋白GFAP蛋白对照组15 1.95ʃ0.17 1.11ʃ0.06 MCAO组15 1.14ʃ0.07① 1.93ʃ0.18①电针治疗组15 1.85ʃ0.16② 1.03ʃ0.02②F值11.33915.586P<0.001<0.001注:MCAO组与对照组比较,①P<0.05;电针治疗组与MCAO组比较,②P<0.05㊂3讨论脑血管病是一种临床的常见疾病,具有高发病率㊁高死亡率㊁高致残率和高复发率的特点[14-15]㊂脑梗死约占脑血管疾病的70%以上,是一类影响大㊁预后差㊁治疗选择有限的脑血管疾病[16-17]㊂针灸治疗阿尔茨海默病㊁中风㊁缺血性脑卒中痉挛性偏瘫可发挥良好的治疗作用,但作用机制尚未明确[18-19]㊂因此,电针治疗脑梗死的作用机制是目前的研究热点㊂本研究证实电针刺激对脑梗死小鼠具有保护作用,可改善MCAO小鼠神经功能损伤,减小脑梗死体积,降低小鼠炎性细胞因子水平等㊂细胞凋亡是神经系统缺氧缺血性神经元丢失的重要原因㊂本研究通过TUNEL荧光染色分析发现,电针降低了MCAO小鼠海马神经元凋亡率;蛋白印迹分析发现电针治疗降低MCAO小鼠凋亡蛋白Bax和Caspase-3表达,促进Bcl-2升高,并抑制MCAO引起的炎性因子水平升高㊂有研究表明,电针可保护或预防MCAO早期和中期功能性神经元死亡,抑制Notch信号通路[20]㊂MCAO后是急性但持久的炎症反应,特点是活性氧产生增加㊂载脂蛋白E(ApoE)在MCAO中具有抗炎㊁抗氧化和抗凋亡特性,电针通过增加ApoE表达,减轻MCAO炎症和氧化应激反应㊂电针刺激在MCAO后发挥神经保护作用,与脑源性神经营养因子和神经营养因子上调相关㊂电针可抑制疼痛信号的诱导和传递,通过重新平衡神经-免疫-内分泌相互作用,介导抗伤害和抗炎作用㊂脑梗死可触发多种细胞信号通路,导致细胞存活或细胞损伤㊂电针通过调节脑梗死大鼠模型中的p38㊁细胞外信号调节激酶(ERK1/2)和c-Jun N末端激酶(JNK)抑制脑梗死区域的细胞凋亡㊂本研究结果显示,MCAO组小鼠Notch3信号通路相关基因表达显著增加;电针通过Notch3信号通路抑制脑梗死海马神经元凋亡,Notch3通路在电针治疗脑梗死中起关键作用㊂EZH2增强子是一种组蛋白甲基转移酶,在细胞分化㊁炎症㊁肌成纤维细胞转化和组织纤维化中发挥着重要作用㊂EZH2在人类胆管癌和系统性硬化症中充当了Notch信号通路的调节剂㊂分子量为2.3kDa的长链非编码RNA(lncRNA)H19在MCAO㊁糖尿病和类风湿性关节炎中高表达,通过调节let-7b㊁Wnt和Notch信号通路促进神经元细胞凋亡㊂H19可调节异常细胞和组织中的EZH2表达,表明其与EZH2参与MCAO的进展㊂EZH2在细胞增殖和凋亡中发挥着关键作用㊂在毛囊干细胞中,EZH2通过抑制miR-22,促进细胞增殖和分化,激活信号转导和转录激活因子3(STAT3),增加细胞凋亡和促炎细胞因子的释放㊂H19与miR-138的相互作用可调节EZH2表达㊂本研究结果表明,H19通过激活Notch3信号通路在提高因子基因表达方面发挥作用,EZH2和H19表达与Notch3信号通路的激活有关㊂相关研究显示,2㊁50 Hz的电针刺激在运动性能方面表现出持续且显著的增强[21]㊂本研究结果显示,2Hz的电针刺激可促进神经干细胞增殖,抑制Notch3信号通路,促进神经干细胞增殖,即抑制Notch3信号通路可作为保护MCAO 的神经元凋亡的靶点㊂综上所述,MCAO可诱导海马神经元凋亡,电针通过调控Notch3激活,调节Bax/Bcl-2及Nestin和GFAP蛋白,从而阻断Caspase-3激活抑制神经元凋亡,该结果可为脑梗死的治疗提供实验依据㊂参考文献:[1]WANG D,LI L J,ZHANG Q,et bination ofelectroacupuncture and constraint-induced movement therapyenhances functional recovery after ischemic stroke in rats[J].Journal of Molecular Neuroscience,2021,71(10):2116-2125. [2]LIU L,ZHANG Q,XIE H Y,et al.Differences in post-ischemicmotor recovery and angiogenesis of MCAO rats followingelectroacupuncture at different acupoints[J].Current NeurovascularResearch,2020,17(1):71-78.[3]LIU L,ZHANG Q,LI M Y,et al.Early post-strokeelectroacupuncture promotes motor function recovery in post-ischemic rats by increasing the blood and brain irisin[J].Neuropsychiatric Disease and Treatment,2021,17:695-702. [4]DUAN X D,ZHANG L,YU J H,et al.The effect of differentfrequencies of electroacupuncture on BDNF and NGFexpression in the hippocampal CA3area of the ischemichemisphere in cerebral ischemic rats[J].NeuropsychiatricDisease and Treatment,2018,14:2689-2696.[5]KANG Z,YE H M,CHEN T,et al.Effect of electroacupuncture atsiguan acupoints on expression of BDNF and TrkB proteins inthe hippocampus of post-stroke depression rats[J].Journal ofMolecular Neuroscience,2021,71(10):2165-2171.[6]XU X Y,FANG Q,HUANG W,et al.Effect of electroacupunctureon neurological deficit and activity of clock and Bmal1in cerebralischemic rats[J].Current Medical Science,2020,40(6):1128-1136.[7]YU Q,LI X H,LI Y M,et al.Effects of combinedelectroacupuncture and exercise training on motor function andmicrotubule-associated protein2expression in the middle andlate stages of cerebral infarction in rats[J].Acupuncture inMedicine,2020,38:175-180.[8]SHA R,HAN X H,ZHENG C X,et al.The effects ofelectroacupuncture in a rat model of cerebral ischemia-reperfusion injury following middle cerebral artery occlusioninvolves microRNA-223and the PTEN signaling pathway[J].Medical Science Monitor,2019,25:10077-10088.[9]XING Y,YANG S D,WANG M M,et al.Electroacupuncturealleviated neuronal apoptosis following ischemic stroke in ratsvia midkine and ERK/JNK/p38signaling pathway[J].Journal ofMolecular Neuroscience,2018,66(1):26-36.[10]CAO S Q,YANG Y F,YU Q,et al.Electroacupuncture alleviatesischaemic brain injury by regulating the miRNA-34/Wnt/autophagy axis[J].Brain Research Bulletin,2021,170:155-161.[11]SHA R,ZHANG B,HAN X H,et al.Electroacupuncture alleviatesischemic brain injury by inhibiting the miR-223/NLRP3pathway[J].Medical Science Monitor,2019,25:4723-4733.[12]WEN T,ZHANG X F,LIANG S X,et al.Electroacupunctureameliorates cognitive impairment and spontaneous low-frequency brain activity in rats with ischemic stroke[J].Journal ofStroke and Cerebrovascular Diseases,2018,27(10):2596-2605.[13]ZHANG Y,MAO X,LIN R H,et al.Electroacupuncture amelioratescognitive impairment through inhibition of Ca2+-mediatedneurotoxicity in a rat model of cerebral ischaemia-reperfusioninjury[J].Acupuncture in Medicine:Journal of the British MedicalAcupuncture Society,2018,36(6):401-407.[14]DONG H,QIN Y Q,SUN Y C,et al.Electroacupunctureameliorates depressive-like behaviors in poststroke rats viaactivating the tPA/BDNF/TrkB pathway[J].NeuropsychiatricDisease and Treatment,2021,17:1057-1067.[15]WANG Z F,LIN B B,LIU W L,et al.Electroacupuncture ameliorateslearning and memory deficits via hippocampal5-HT1A receptors andthe PKA signaling pathway in rats with ischemic stroke[J].Metabolic Brain Disease,2020,35(3):549-558.[16]WANG H Y,CHEN S H,ZHANG Y M,et al.Electroacupunctureameliorates neuronal injury by Pink1/Parkin-mediated mitophagyclearance in cerebral ischemia-reperfusion[J].Nitric Oxide:Biology and Chemistry,2019,91:23-34.[17]CHEN B,LIN W Q,LI Z F,et al.Electroacupuncture attenuatesischemic brain injury and cellular apoptosis via mitochondrialtranslocation of cofilin[J].Chinese Journal of IntegrativeMedicine,2021,27(9):705-712.[18]WANG H L,LIU F L,LI R Q,et al.Electroacupuncture improveslearning and memory functions in a rat cerebral ischemia/reperfusion injury model through PI3K/AKT signaling pathwayactivation[J].Neural Regeneration Research,2021,16(6):1011-1016.[19]ZHANG G F,YANG P,YIN Z,et al.Electroacupuncturepreconditioning protects against focal cerebral ischemia/reperfusion injury via suppression of dynamin-related protein1[J].Neural Regeneration Research,2018,13(1):86-93. [20]CHEN X B,SUN X Y,LIU N H,et al.Analysis of human acupointbiological information and neural electric activity based onultrasonographic image[J].World Neurosurgery,2021,149:481-491.[21]JIA B R,CHENG C Y,LIN Y W,et al.The2Hz and15Hzelectroacupuncture induced reverse effect on autonomic functionin healthy adult using a heart rate variability analysis[J].Journalof Traditional and Complementary Medicine,2011,1(1):51-56.(收稿日期:2022-06-21)(本文编辑薛妮)。

中药复方开心散调控慢性压力应激小鼠海马神经新生抗抑郁作用机制研究郑嘉妮;黄灵欣;陆韫青;李璇;陈杨;仝佳祥;朱梓强;段金廒;李乐军;朱悦【期刊名称】《世界科学技术-中医药现代化》【年(卷),期】2024(26)1【摘要】目的评价开心散调控慢性不可预知压力应激模型小鼠海马神经新生抗抑郁效用,探讨作用机制。
方法采用慢性不可预知压力应激构建抑郁小鼠模型,给以开心散水提取物。
通过糖水偏嗜率检测、强迫游泳实验、悬尾实验等评价模型小鼠抑郁样行为。
采用免疫荧光法检测模型小鼠海马区神经新生情况。
采用蛋白免疫印迹法检测小鼠海马神经干细胞标志物巢蛋白以及Wnt/β-catenin信号通路相关蛋白的表达。
采用ELISA法检测相应组织中压力应激因子如促肾上腺皮质激素释放因子、促肾上腺皮质激素、皮质醇与神经营养因子如脑源性神经营养因子、神经生长因子的表达。
采用小鼠原代神经干细胞,采用免疫荧光法检测开心散对原代神经干细胞增殖情况,采用蛋白免疫印迹法检测巢蛋白以及Wnt/β-catenin信号通路相关蛋白的表达。
结果开心散水提取物显著增加抑郁模型小鼠糖水偏嗜率,降低强迫游泳与悬尾不动时间;同时促进海马组织神经元新生,降低压力应激因子表达,提高神经营养因子表达,上调巢蛋白表达并调控Wnt/β-catenin信号通路。
开心散提取物还可促进小鼠原代神经干细胞增殖及蛋白表达,并促进β-catenin入核,调控Wnt/β-catenin信号通路。
结论开心散具有改善小鼠抑郁样行为效用,促进海马神经新生以及调控Wnt/β-catenin通路可能是其抗抑郁的重要作用环节与机制。
【总页数】12页(P19-30)【作者】郑嘉妮;黄灵欣;陆韫青;李璇;陈杨;仝佳祥;朱梓强;段金廒;李乐军;朱悦【作者单位】南京中医药大学药学院/江苏省方剂高技术研究重点实验室/中药资源产业化与方剂创新药物国家地方联合工程研究中心/江苏省中药资源产业化过程协同创新中心;南京中医药大学附属苏州市中医医院【正文语种】中文【中图分类】R285.5【相关文献】1.抗抑郁剂对慢性应激小鼠海马神经元再生的影响2.开心散配伍比例对慢性压力应激抑郁小鼠皮层与海马神经营养因子系统调控的影响3.中药复方开心散调控神经营养因子抗抑郁物质基础与作用机制研究4.麋鹿角调控慢性压力应激小鼠神经营养因子和压力应激因子抗抑郁作用研究5.中药复方开心散调控慢性压力应激小鼠海马炎性细胞因子水平抗抑郁作用机制研究因版权原因,仅展示原文概要,查看原文内容请购买。

细胞与分子生物学TSA对不同糖浓度下小鼠海马神经元HT-22细胞凋亡的影响许雯1,2,许永劼3,刘歆蕾2,沈婕2,朱科静2,林海容2,李兴4,潘卫1,2,3△摘要:目的探究曲古抑菌素A(TSA)对不同糖浓度下小鼠海马神经元细胞系HT-22细胞凋亡的影响。
方法分别用高糖培养基(葡萄糖浓度55mmol/L)及普通培养基(葡萄糖浓度25mmol/L)培养小鼠海马神经元HT-22细胞,高糖培养基及普通培养基各分为对照组(NC组)和抑制剂组(TSA组)。
用1mmol/L的TSA作用细胞1、4、8h,同时用0.4mmol/L的TSA作用细胞1、4、8、12、14、16、20、24h,CCK8法检测细胞存活率,确定抑制剂最佳作用浓度和作用时间;酶联免疫吸附测定(ELISA)法检测组蛋白去乙酰化酶(HDAC)和组蛋白乙酰转移酶(HAT)活性;流式细胞术检测细胞凋亡情况;蛋白免疫印迹法检测B淋巴细胞瘤-2(Bcl-2)、Bcl-2相关X蛋白(Bax)、半胱氨酸天冬氨酸蛋白酶3(Caspase-3)的表达情况。
结果以0.4mmol/L、20h为TSA处理HT-22细胞的最适条件;ELISA结果显示,TSA作用后,HDAC显著减少,HAT显著增加(P<0.05);流式细胞术结果显示,在TSA抑制20h后,细胞凋亡率显著增加,高糖环境下加入TSA,细胞凋亡情况更加严重;TSA作用20h后,Bcl-2表达显著下调,Caspase-3显著上调(P<0.05)。
结论TSA可能通过抑制HDAC增加组蛋白乙酰化水平,导致小鼠神经元HT-22细胞的凋亡。
关键词:葡萄糖;糖尿病;脑疾病;海马;细胞凋亡;组蛋白脱乙酰基酶类;曲古抑菌素A中图分类号:R587.2文献标志码:A DOI:10.11958/20202807Effects of TSA on the apoptosis of HT-22cells in mouse hippocampal neurons under differentconcentrations of glucoseXU Wen1,2,XU Yong-jie3,LIU Xin-lei2,SHEN Jie2,ZHU Ke-jing2,LIN Hai-rong2,LI Xing4,PAN Wei1,2,3△1The Affiliated Hospital of Guizhou Medical University,Guizhou Prenatal Diagnosis Center,Guiyang550004,China;2School of Clinical Laboratory Science,Guizhou Medical University;3School of Public Health,Guizhou Medical University;4Guizhou University of Traditional Chinese Medicine△Corresponding Author E-mail:****************Abstract:Objective To explore the effect of trichostatin A(TSA)on the apoptosis of mouse hippocampal neuron cell line HT-22cells under different sugar concentrations.Methods The mouse hippocampal neuron HT-22cells in high glucose medium(glucose concentration55mmol/L)and normal medium(grape concentration25mmol/L)were cultured.The cells in high glucose medium and normal medium were divided into control group(NC group)and inhibitor group(TSA group).Cells were treated with1mmol/L TSA for1,4and8h,or0.4mmol/L TSA for1,4,8,12,14,16,20and24h.The cell viability was detected by the CCK8method after treatment.Histone deacetylase(HDAC)and histone acetyltransferase (HAT)enzyme activities were detected by ELISA.Cell apoptosis was detected by flow cytometry.The expression levels of B lymphoma-2(Bcl-2),Bcl-2related X protein(Bax)and cysteine protease3(Caspase-3)were detected by Western blot assay.Results The0.4mmol/L of TSA and20hours were the optimal condition for culturing HT-22cells after testing. ELISA results showed that after treatment with TSA,HDAC significantly decreased and HAT increased significantly(P<0.05).Flow cytometry results showed that after TSA inhibiting for20h,the apoptosis rate increased significantly.While TSA was added to cells under high glucose conditions,the apoptosis rate increased more.After20hours of TSA treatment,Bcl-2 expression was significantly down-regulated,while Caspase-3was significantly up-regulated(P<0.05).Conclusion TSA基金项目:国家自然科学基金地区科学基金项目(81960151,81960822);贵州省教育厅创新群体重大研究项目(黔教合KY字[2018]021号);贵州省科技计划项目(黔科合支撑[2019]2802号)作者单位:1贵州医科大学附属医院贵州省产前诊断中心(邮编550004);2贵州医科大学医学检验学院;3贵州医科大学公共卫生学院;4贵州中医药大学作者简介:许雯(1994),女,硕士在读,主要从事糖尿病慢性并发症发病机制研究。

大脑提升能力的六种方法大脑是一个让人迷惑的器官,就像生和死、意识、睡眠和其他更多的东西,都是人类至今也没有解开的谜团。
下面就是小编给大家带来的大脑提升能力的六种方法,希望大家喜欢!大脑提升能力的六种方法:科学家发现,成年人大脑的可塑性远比预计的强。
我们的日常行为和周边环境,都能引起大脑神经回路的重新连接、功能重组和位置的改变。
有些人甚至认为,不同的思考方式也足以重塑大脑。
方法一:运动研究表明,小鼠在滚轴上训练一段时间之后,海马区神经元数量显著增加,在学习和记忆测试中表现得更好。
针对人类的研究也显示,体育锻炼能提高大脑的执行能力(包括计划、组织、多任务处理能力等)。
众所周知,运动能改善情绪,经常运动的人老了以后也不容易患痴呆症(dementia)。
常运动的老人的执行力,比久坐不动的老人更强。
即使是那些在沙发上窝了一辈子的老人,如果现在就开始运动,也能提高执行力。
这种现象可能与很多机制有关。
运动可以增加大脑的血流量,将更多的氧分、能量和营养物质运送给忙碌的神经细胞。
研究表明,运动可以促进大脑分泌神经营养因子(brain-derived neurotrophic factor, BDNF),促进神经细胞的生长,改善神经细胞间的联系,提高它们的存活率。
不过,所有研究都无法解释“四肢发达,头脑简单”又是怎么回事。
你知道吗?·大量研究表明,运动能改善睡眠质量,提高免疫力——还有它办不到的事吗?·想让大脑从运动中获益,我们并不需要像查克·诺里斯(chuck norris,美国动作电影演员)一样成为世界空手道冠军。
针对老人的研究表明,每天步行20分钟,就能保持身心健康。
方法二:饮食大脑和身体一样需要营养和能量。
那么,哪些食物能提升大脑智力,哪些又会损害智力呢?众所周知的坏家伙是饱和脂肪酸,它对大脑和身体健康都有严重的不良影响。
如果给大鼠喂食饱和脂肪酸含量较高的食物,它们在学习和记忆测试中就会表现不佳,常吃这类食物的人患痴呆症的风险也较高。
2024北京海淀高三一模生 物2024.04本试卷共10页,100分。
考试时长90分钟。
考生务必将答案答在答题卡上,在试卷上作答无效。
考试结束后,将本试卷和答题卡一并交回「第一部分本部分共15题,每题2分,共30分。
在每题列出的四个选项中,选出最符合题目要求的一项。
1.乳酸菌、黑藻叶肉细胞、人体小肠上皮细胞虽形态各异,但它们也有共同之处,表现在A.有细胞膜和核膜B.可进行有丝分裂C.以DNA作为遗传物质D.线粒体中进行能量转换2.端粒是染色体末端的一段DNA片段。
端粒酶由RNA和蛋白质组成,该酶能结合到端粒上,以自身的RNA 为模板合成并延伸端粒DNA。
在正常情况下,端粒酶只在不断分裂的细胞中具有活性。
下列有关端粒酶的叙述,正确的是A.仅由C、H、O、N四种元素组成B.催化过程以4种脱氧核苷酸为底物C.组成成分都在核糖体上合成D.在所有细胞中均具有较高活性3.新生无毛哺乳动物体内存在一种含有大量线粒体的褐色脂肪组织,褐色脂肪细胞的线粒体内膜含有蛋白质U。
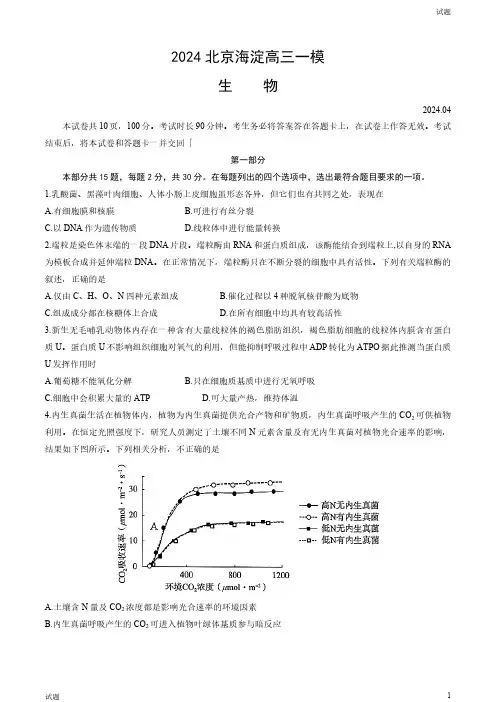
蛋白质U不影响组织细胞对氧气的利用,但能抑制呼吸过程中ADP转化为ATPO据此推测当蛋白质U发挥作用时A.葡萄糖不能氧化分解B.只在细胞质基质中进行无氧呼吸C.细胞中会积累大量的ATPD.可大量产热,维持体温4.内生真菌生活在植物体内,植物为内生真菌提供光合产物和矿物质,内生真菌呼吸产生的CO2可供植物利用。
在恒定光照强度下,研究人员测定了土壤不同N元素含量及有无内生真菌对植物光合速率的影响,结果如下图所示。
下列相关分析,不正确的是A.土壤含N量及CO2浓度都是影响光合速率的环境因素B.内生真菌呼吸产生的CO2可进入植物叶绿体基质参与暗反应C.图中A点制约植物光合速率的主要环境因素是光照强度D.在土壤高N含量下内生真菌可提高植物的光合速率5.C品系果蝇仅在X染色体上存在D基因,使其具有棒眼性状,果蝇中D基因纯合会致死。
用X射线照射大量雄果蝇,为检测照射后的这批雄果蝇X染色体是否发生突变,使它们与C品系雌蝇交配。



《基于AMPK-PGC-1α信号通路探讨形神共养改善衰老小鼠海马区老化程度的机制研究》基于AMPK-PGC-1α信号通路探讨形神共养改善衰老小鼠海马区老化程度的机制研究一、引言随着人类社会不断发展,老龄化问题逐渐成为社会关注的焦点。
随着年龄的增长,机体各系统功能逐渐衰退,尤其是中枢神经系统,如海马区,其老化与认知功能下降、记忆力减退等密切相关。
因此,探讨如何延缓中枢神经系统老化、提高生活质量成为科研工作的重点。
近年来,形神共养作为一种传统的养生方法,在抗衰老、改善认知功能等方面显示出良好的效果。
本研究基于AMPK/PGC-1α信号通路,探讨形神共养对衰老小鼠海马区老化程度的改善机制。
二、材料与方法1. 实验动物与分组选用一定数量的健康老年小鼠作为实验对象,根据实验需求将其分为实验组和对照组。
2. 形神共养干预方法实验组小鼠接受形神共养干预,包括适当的运动锻炼、合理的饮食调整及中医养生方法等。
对照组小鼠则不进行任何干预。
3. 取材与样本处理通过取材得到实验组和对照组小鼠的海马区组织样本,采用冷冻、切片、固定等处理后,用于后续实验分析。
4. 实验方法与检测指标采用免疫组化、荧光定量PCR、Western Blot等方法检测AMPK、PGC-1α等关键分子的表达水平及海马区组织形态学变化等指标。
三、实验结果1. 形神共养对小鼠海马区组织形态学的影响实验结果显示,经过形神共养干预后,实验组小鼠的海马区组织形态学较对照组有明显改善,神经元结构更加清晰,神经纤维排列更加整齐。
2. 形神共养对AMPK/PGC-1α信号通路的影响实验组小鼠的AMPK和PGC-1α表达水平较对照组明显升高,表明形神共养可以激活AMPK/PGC-1α信号通路。
此外,通过进一步分析发现,形神共养还可以调节相关下游分子的表达,如线粒体相关基因等。
3. 形神共养对小鼠认知功能的影响通过行为学实验发现,经过形神共养干预后,实验组小鼠的认知功能较对照组有明显改善,如空间记忆能力、学习能力等。

---文档均为word文档,下载后可直接编辑使用亦可打印---摘要】目的:研究不同浓度的氯化钴 (CoCl2)诱导小鼠海马神经元细胞(HT22)缺氧损伤模型的比较。
方法:用不同浓度 (0、50、100、200μmol/L) CoCl2建立HT22细胞缺氧模型。
通过HE染色观察HT22细胞轴突生长的特征,CCK8法检测HT22细胞体外增殖活性的变化,同时,Western Blot检测HT22细胞中HIF-1α、tau5的表达情况。
结果: HE显示CoCl2可致HT22细胞轴突断裂,CCK8显示CoCl2对HT22细胞增殖抑制作用表现出浓度依赖性。
Western Blot显示CoCl2浓度为200μmol/L时, HIF-1α、Tau5蛋白表达与对照组相比差异有统计学意义。
结论:CoCl2可剂量依赖性上调HT22细胞HIF-1α表达,致轴突损伤及tau5降低,建立细胞缺氧模型简单易行。
【关键词】HT22细胞; 氯化钴; 轴突;HIF-1αComparison of hypoxic injury models of hippocampal neurons induced by different concentrations of cobalt chloride in miceXiao Tingting1, Wang Li1,Cao Xiangmei2,Ni Xinli1 *( 1.School of Clinical Medicine, 2.School of Basic Medicine,,Ningxia MedicalUniversity Yinchuan 750004, China)[Abstract]: Objective: To compare the hypoxia injury model of mouse hippocampal neuron cells (HT22) induced by different concentrations of cobalt chloride (CoCl2). Methods: The hypoxia model of HT22 cells was established with different concentrations (0, 50, 100, 200 μmol / L) of CoCl2. The characteristics of axon growth of HT22 cells were observed by HE staining. The CCK8 method was used to detect the changes of HT22 cell proliferation activity in vitro. At the same time, Western Blot was used to detect the expression of HIF-1α and tau5 in HT22 cells. Results: HE showed that CoCl2 could cause axon rupture in HT22 cells, and CCK8 showed that CoCl2 had a concentration-dependent inhibition effect on HT22 cell proliferation. Western Blot showed that when the concentration of CoCl2 was 200 μmol / L, the expressions of HIF-1α and Tau5 protein were significantly different from those of the control group. Conclusion: CoCl2 can up-regulate HIF-1α expression in HT22 cells in a dose-dependent manner, cause axonal damage and decrease tau5, and it is easy to establish a model of cell hypoxia.[Keywords] HT22 cells; Cobalt chloride; axons; HIF-1α缺氧缺血性(hypoxia ischemia,HI)脑损伤是窒息、心脏骤停以及心肺复苏后最常见的神经系统后遗症,也是致残致死的严重疾病之一。
小鼠海马区神经元的培养:
实验材料:
1、所有用于细胞培养的器械、器皿剂溶液必须灭菌消毒。
2、多聚赖氨酸铺板以磷酸盐缓冲液(PBS)或蒸馏水荣分解多聚赖氨酸(分子量为30-70KD)至1mg/ml。
分装储存。
临用前稀释至终浓度为20-60mg/ml过滤备用。
在培养板或皿中加少量多聚赖氨酸溶液,以覆盖底部为宜,在37℃、CO2孵箱中过夜。
次日,用移液管吸取多聚赖氨酸,并用水洗2-3遍,晾干备用,如果培养目的适用于电生理的单细胞记录或者免疫组化和荧光染色,则在培养皿的底部加上盖玻片,多聚赖氨酸铺于盖玻片上,则更易得到强烈的荧光信号。
3解剖液(HEPES平衡盐溶液)取100ml 10x 平衡盐溶液(Hank’s balance salt solution,HBSS)
HEPES 3.9g,
NaHCO3 0.84g,
青霉素104 U,
链霉素100mg,
H2O 800ml。
以1mol/LHCl调节PH至7.2,再加H2O之后总体积为1L,以0.2μm直径的滤菌器过滤,4℃保存。
4、DMEM 培养基(Dulbecco’s modified Eagle’s medium)DMEM 培养基500ml,加小牛血清和或马血清各10%(血清需经56℃,30min 灭活)1% 的青/链霉素.
5、无血清培养基最常用的一种无血清培养基为,Neurobasal+2%的b27+1%抗生素
6、2.5%胰蛋白酶(typsin)溶液临用前以解剖液稀释至终浓度0.125%。
7、台盼蓝(typan blue )溶液终浓度为0.2%-0.4%。
8、阿糖胞苷终浓度为8-10 μmol/L。
9、动物妊娠天数的选择通常选择出生24h之内的乳鼠。
实验步骤:
1、脱臼处死乳鼠,75%的酒精浸泡3-5min,将清洗完的乳鼠放入含预冷解剖液的培养皿中。
2、用两把尖镊逐个去除鼠的头部皮肤和颅骨,取出全脑,置于另一含预冷解剖液的(PBS?)培养皿中,小心剥离并去除脑膜及脉络丛。
3、在中线处用尖镊分离大脑半球,在解剖显微镜下小心去除嗅球、中隔、丘脑及下丘脑,剥离脑膜及脉络丛,在大脑皮层下方小心取出海马,置于含冰冷解剖液的6cm培养皿中。
4、用弯头尖镊把海马剪成小碎片,收集全部海马碎片,置于一含有0.25%胰蛋白酶溶液的培养皿中(该溶液必须在37℃孵箱中预暖),在孵箱中消化5-7min。
5、收集全部海马碎片,置于10ml预暖的DMEM培养基的试管中(培养基中含有10%的胎牛血清和或马血清各,1%抗生素,以及谷氨酰胺)。
最好用培养基洗2-3遍,以中止消化反应。
6、加少量培养基于试管中,以中空塑料管反复吹打10-15次,直成细胞悬液。
加培养基于同一试管中至约10ml,静置约10min。
取1-2μl细胞悬液于99μl0.2%-0.4%台盼蓝溶液中,用细胞计数板计算存活和死亡的细胞数,从而测定细胞密度(细胞计数:活体细胞具有排斥台盼蓝的能力,而死亡细胞则能吸收台盼蓝变成蓝色)。
7、根据实验需要,以培养基稀释细胞悬液至所需的细胞密度,接种细胞至经多聚赖氨酸铺板过夜处理的培养皿或培养板中。
通常细胞密度为(2-2.5)x105细胞/cm2.
8、待6-8h细胞贴壁后,换培养基为含2%B27的Neurobasal,含有B27 和0.5mM的谷氨酰胺。
9、培养2-3d培养基中加入8-10μmol的阿糖胞苷,以抑制或阻止非神经细胞和原代胶质增殖。
10、每三天用Neuronbasal使用液半量换液
三、形态学观察
细胞接种6-12h,开始贴壁,细胞成圆形或椭圆形,两周后神经元最为丰满,胞体圆形或多角形,四周晕光明显,胞核及核仁清晰可见,位于细胞中央或一边,神经多而粗大,分支成网络。
培养一月后,细胞开始退化。
一般认为,培养2-4周开始实验比较适宜。
附:
血细胞计数法及台盼蓝测细胞活度
(一)
实验材料和实验前准备
1、显微镜,血细胞计数器,盖玻片
2、毛细移液管,1 ml血清移液管,5ml试管,擦镜纸或软布
3、0.4%的台盼蓝溶液,0.3%苯胺黑(nigrosin)溶液,95%,乙醇
4、用擦镜纸或者软布将血细胞计数板的计数池擦干净。
再用酒精95%血细胞技术板和盖
玻片,晾干备用。
5、讲清洁干净的盖玻片覆盖于血细胞技术板的计数池上。
6、胞浓度=4大格细胞数/4×稀释倍数×104/m
(二)实验步骤
1、制备制备细胞悬液取一试管,分别加入0.4%台盼蓝溶液0.2ml (或0.3%苯胺黑溶液)及混合均匀的细胞悬液0.8 ml,用手轻柔振荡试管,以混合悬液和染料。
2、以中空移液管轻柔混匀细胞悬液后,立即用毛细移液管吸取少量细胞悬液,置移液管尖头于细胞计数板计数池的边缘(小心不要移动盖玻片),缓缓释放细胞悬液。
通常一侧计数池可以容纳20 ul细胞悬液。
3、细胞计数每个正方形的边侧都有三根线,在左侧和上方,应将接触到三根线的中线的内侧线的细胞计算在内,技术两侧计数池,共八个正方形。
四、注意事项
1、L-多聚赖氨酸(0.1mg/ml)包被培养板后,要待其干后才能接种,因为其对神经元有毒性。
L-多聚赖氨酸(0.1mg/ml)能回收再利用。
2 、取脑组织时动作尽量要快,并且尽量低温操作。
3 、分离皮层和海马时,要尽量分离脑膜和血管,否则有大量的成纤维细胞。
4 、用吸管轻轻吹打细胞时,尽量不起泡沫,以免损伤神经元。
5 、接种细胞的过程,动作要快,以免细胞聚集。
6 、接种细胞后,24h勿移动,以免影响细胞贴壁。
五、结果、
大多数神经元在接种1h内贴壁,3-5h开始变平,并长出1-2个突起。
3天后,许多细胞从胞体长出数个突起。
神经元在7-10天开始成熟,胞体逐渐长至30-40um,晕光明显,胞核和核仁清晰可见,突起变粗,变长并且有分支,边缘清楚,发亮,形成明显的神经网络。
神经元可存活1-2月。