应用分子生物学与同位素示踪技术研究厌氧氨氧化菌活性及功效_叶磊
- 格式:pdf
- 大小:281.13 KB
- 文档页数:6

中国环境科学 2021,41(12):5912~5920 China Environmental Science 微生物诱导碳酸盐沉积介导的Cd2+固定朱广森,汪文军,吴峥,司友斌*(安徽农业大学资源与环境学院,农田生态保育与污染防控重点实验室,安徽合肥230036)摘要:实验室条件下,研究了碳酸盐矿化菌施氏假单胞菌(Pseudomonas stutzeri)、枯草芽孢杆菌(Bacillus subtilis)和巴氏芽孢杆菌(Bacillus pasteurii)的生长对Cd2+的耐受性和固定效果,以及羟基磷灰石对3种菌固定Cd2+的影响,通过扫描电镜(SEM)、X射线能谱(EDS)、傅里叶红外光谱(FTIR)和X射线衍射(XRD)对碳酸盐矿化菌的诱导矿化产物进行了表征.结果表明,3种菌生长过程中B. pasteurii的脲酶活性最强,是P. stutzeri和B. subtilis脲酶活性的10倍左右,且P. stutzeri, B. subtilis, B. pasteurii产CO32-的最高浓度分别为588.19, 661.72, 1735.18mg/L,培养体系中pH值均呈现升高的趋势,最高pH值分别为8.23, 9.06, 9.52;Cd2+浓度在0~10mg/L范围内对P. stutzeri的生长影响较小,而当Cd2+浓度高于1mg/L时则会抑制B. subtilis和B. pasteurii的生长.初始Cd2+浓度为0.5mg/L时,培养120h, P. stutzeri, B. subtilis和B. pasteurii对Cd2+的固定去除率分别为96.37%, 99.40%, 97.57%;加入羟基磷灰石能够提高碳酸盐矿化菌对Cd2+的去除率.SEM和EDS结果显示,P. stutzeri和B. subtilis诱导形成的矿化产物多聚集在菌体周围或附着在菌体表面,呈球状或网状结构,表面疏松多孔,B. pasteurii的矿化产物附着在菌体表面,结构致密,呈不规则的球状; FTIR分析表明矿化产物中存在CO32-,结合XRD结果,证实3种菌诱导沉淀矿化产物主要是CaCO3,而Cd2+与Ca2+会以同晶置换的方式形成CdCO3晶体,B. pasteurii在诱导矿化的过程中可将Cd2+以Ca0.67Cd0.33CO3共沉淀的方式固定.关键词:微生物矿化;Cd2+;羟基磷灰石;碳酸盐沉淀中图分类号:X172 文献标识码:A 文章编号:1000−6923(2021)12-5912-09Immobilization of Cd2+ by microbial induced carbonate precipitation. ZHU Guang-sen, WANG Wen-jun, WU Zheng, SI You-bin* (Anhui Province Key Laboratory of Farmland Ecological Conservation and Pollution Prevention, School of Resources and Environment, Anhui Agricultural University, Hefei 230036, China). China Environmental Science, 2021,41(12):5912~5920 Abstract:Under laboratory culture conditions, three carbonate mineralization bacteria, Pseudomonas stutzeri, Bacillus subtilis and Bacillus pasteurii were used to study the growth specialities, the resistance and removal of Cd2+ by the bacteria, and the effect of hydroxyapatite on Cd2+ immobilization. The scanning electron microscope (SEM), energy dispersive spectrum (EDS), fourier transform infrared spectroscopy (FTIR) and X-ray diffraction (XRD) were employed to characterize the biomineralization products. The results showed that the urease activity of B. pasteurii was about 10 times than those of P.stutzeri and B.subtilis. The CO32- was produced and the pH of culture media had an increasing during the growth of the three bacteria. The highest CO32-concentrations were 588.19, 661.72, 1735.18mg/L, while the highest pH value could reach 8.23, 9.06 and 9.52 for P. stutzeri, B. subtilis and B. pasteurii, respectively. The growth of P. stutzeri was not affected within the Cd2+ concentration of 0~10mg/L, while the growth of B. subtilis and B. pasteurii were inhibited in the Cd2+ concentration beyond 1mg/L. With the initial Cd2+ concentration of 0.5mg/L and biomineralization for 120h, the removal rates of Cd2+ by P. stutzeri, B. subtilis and B. pasteurii were 96.37%, 99.40% and 97.57%, respectively. The addition of hydroxyapatite could enhance the immobilization of Cd2+ by the bacteria to a certain extent. SEM and EDS analysis showed that the mineralized products induced by P. stutzeri and B. subtilis were mostly clustered around or attached to the surface of the bacteria, with spherical, reticulated structure and porous surface, while the mineralized products by B. pasteurii were attached to the surface of the bacteria, with compact structure and irregular spherical structure. FTIR analysis suggested that there was CO32- in the mineralized products. XRD identified, the mineralized products CaCO3 induced by the three bacteria. The CdCO3 crystal were forms by isomorphic substitution of Ca2+ with Cd2+. As an alternative, Ca0.67Cd0.33CO3 co-precipitation were induced by B. pasteurii for Cd2+ immobilization.Key words:microbiological mineralization;cadmium;carbonate precipitation;hydroxyapatite近年来由于铅锌矿的开采、冶炼、大气沉降和农业化学用品的高通量投入等途径的增多,导致重金属镉(Cd)的环境污染问题日益严重[1].重金属Cd 通常存在于土壤,岩石和水体等环境中,经食物链传递到人体中,从而导致不可逆转的慢性肝损伤,骨质疏松症,癌症等疾病[2].据报道,我国Cd污染超收稿日期:2021−04−09基金项目:国家重点研发计划(2019YFC1805203);国家自然科学基金资助项目(42077123);安徽高校协同创新项目(GXXT-2021-061)* 责任作者, 教授,*****************.cn12期朱广森等:微生物诱导碳酸盐沉积介导的Cd2+固定 5913标土壤达7%[3].生物矿化修复因其成本低廉,适应性强,原位无二次污染等特点[4],近年来逐渐成为环境修复领域的热点.微生物诱导碳酸盐沉淀(MICP)是近年来生物矿化领域研究热点之一[4].目前,国内外学者已将MICP成功应用于Pb, Cd, Zn, Cu, Sr和Ni等多种重金属及放射性金属污染修复[5-6].MICP技术关键在于微生物的选择,目前多以从污染土壤中筛选分离出稳定高效的溶脲菌为研究对象[7],例如P. stutzeri Pb12[8], Stenotrophomonas rhizophila A323, Variovorax boronicumulans C113[9]和Enterobacter sp.(CJW-1)[10]等.有研究表明,上述溶脲菌对污染土壤中的Pb, Cd, Zn等重金属具有较高的固定效果.此外,有些菌对环境的适应能力较强,如在10℃的低温条件下, Exiguobacterium undae YR10经过14d培养后可转化污染土壤中90%以上的Cd2+[11].有些研究还进一步证实MICP将可交换态重金属以碳酸盐结合态形式固定[12],或通过形成方解石、六方岩和白云石等生物矿物吸附等途径将重金属固定[10].然而微生物Pseudomonas stutzeri, Bacillus subtilis和Bacillus pasteurii对重金属Cd的固定机制以及化学材料的添加对微生物诱导碳酸盐沉积固定重金属效果的影响研究相对较少,因此,本研究可为环境生物矿化修复技术提供理论参考,也为微生物与化学材料联合修复重金属污染环境提供实践基础.本研究以P. stutzeri, B. subtilis和B. pasteurii 3种碳酸盐矿化菌作为对象,研究碳酸盐矿化菌生长过程中的脲酶活性、产CO32-能力、pH值变化,以及碳酸盐矿化菌对Cd2+耐受性和固定去除效果,探究了羟基磷灰石联合碳酸盐矿化菌对Cd2+固定的影响,并通过SEM, EDS, FTIR, XRD等表征矿化产物的特征,阐明碳酸盐矿化菌的生物矿化及固定Cd2+的作用机制,以期为微生物诱导碳酸盐沉积用于重金属污染土壤与地下水的生物修复提供理论参考.1 材料与方法1.1 试验材料无水氯化镉(CdCl2,分析纯,纯度≥99%),尿素(CO(NH2)2,分析纯,纯度≥99%),乙酸钙(Ca (C2H3O2)2,分析纯,纯度≥98%),均购于上海阿拉丁生化科技股份有限公司;羟基磷灰石(Ca10(PO4)6(OH)2),Ca/P物质的量比1.67,密度3.16g/cm3,粒径4.5μm,溶解度0.04mg/L,购自西安冰禾生物科技有限公司.菌种来源:施氏假单胞菌(P. stutzeri ATCC 17588)由中国微生物菌种保存库提供,枯草芽孢杆菌(B.subtilis A TCC 6633),巴氏芽孢杆菌(B.pasteurii ATCC 11859)由北纳创联生物技术有限公司提供.1.2培养基与菌悬液制备碳酸盐矿化菌培养基(NBU培养基):牛肉膏3.0g,蛋白胨10.0g,氯化钠5.0g,尿素20.0g, pH=(7.0±0.2),蒸馏水定容至1000mL.除尿素外其他组分121℃高压灭菌30min,然后将配制好的尿素溶液过0.22μm一次性无菌滤膜加入其中.菌悬液的制备[13]:在无菌条件下,将保存的P. stutzeri, B. subtilis, B. pasteurii接种到固体NBU 培养基中,在30℃,150r/min条件下培养24h,无菌条件下将碳酸盐矿化菌转移至已灭菌的液体NBU培养基中,培养24h后以4000r/min离心10min,用0.9%无菌生理盐水冲洗2~3次,使菌体重悬于无菌生理盐水.1.3试验方法1.3.1 碳酸盐矿化菌生长过程中脲酶活性,CO32-浓度及pH值变化测定采用电导率法测定微生物的脲酶活性,测量方法:取1mL菌液加入9mL 1.11mol/L的尿素溶液混匀,用电导率测定仪测量混合体系在25℃时5min电导率变化量,记录5min内的电导率平均变化量(mS/(cm⋅min))乘以稀释倍数即为菌液的脲酶活性(mmol/(L⋅min))[14].吸取2mL制备好的菌悬液接种到200mL已灭菌的培养基中,恒温振荡培养箱中30℃,150r/min连续培养72h,分别设置3组平行.培养过程中按照0,2,4,8,12,24,36,48,60,72h连续取样,每次取样5mL, pH计测定溶液pH值.CO32-浓度测定时按0,2,4,8, 12,24,48,72,96,120h连续取样,每次取样5mL,以4000r/min离心10min,然后取1mL,置于50mL锥形瓶中稀释5倍,采用化学滴定法(DZ/T 0064.49-1993)[15]测定[16].1.3.2 碳酸盐矿化菌对Cd2+耐受性试验吸取1mL制备好的菌悬液至Cd2+浓度分别为0,1,3,5,10, 15mg/L的100mL已灭菌的NBU培养基中,每组浓5914 中国环境科学 41卷度梯度设3组平行,30℃,150r/min下振荡培养72h,连续取样时间同1.3.1,每次取样3mL,测定各处理OD600值.其中尿素溶液配制好后过0.22μm一次性无菌滤膜加入培养基.1.3.3 碳酸盐矿化菌对Cd2+的固定试验吸取1mL制备好的菌悬液至含有0.5mg/L Cd2+和25mmol/L Ca2+ 的100mL已灭菌NBU培养基中,对照组接种等量无菌水,设置3组平行.置于30℃,150r/min下振荡培养120h,连续取样时间同1.3.1,每次取样5mL,4000r/min离心15min,利用ICP-MS测定上清液中Cd2+浓度.在培养120h后取适量样品进行扫描电镜与能谱分析,剩余样品以8000r/min离心15min,弃上清液,置于65℃烘箱中烘干4h后,用玛瑙研钵磨碎过200目筛后,对样品进行傅里叶红外光谱和X射线衍射能谱(XRD)分析.1.3.4 羟基磷灰石对碳酸盐矿化菌固定Cd2+的影响试验将100mg羟基磷灰石(Ca10(PO4)6(OH)2)加入到培养基中灭菌,冷却配制成200mL含1mg/L Cd2+的NBU培养基,再加入2mL制备好的菌悬液,对照组接种等量无菌水,设置3组平行,置于30℃, 150r/min下振荡培养120h, 连续取样时间同1.3.1,每次取样5mL, 4000r/min离心15min,利用ICP-MS 测定上清液中Cd2+浓度.1.4扫描电镜与能谱分析取20mL 1.3.3 中矿化样品于50mL离心管中, 4000r/min离心10min,弃上清液,以2.5%戊二醛固定4~12h,再以0.1mol/L磷酸盐缓冲液淋洗,4000r/ min离心10min,重复2次,弃上清液,依次以不同浓度乙醇梯度脱水,CO2临界点干燥仪干燥10h,镀膜处理后取适量样品进行扫描电镜(SEM,日立S-4800型)及能谱(EDS)分析,观察样品形貌和分析元素组成.1.5傅里叶红外光谱分析采用压片法制备样品,取1~2mg 1.3.3中矿化产物样品在玛瑙研钵中与干燥KBr粉末混合研磨均匀,装入模具内,在压片机上压制成片,进行傅里叶红外光谱(FTIR, Thermo Nicolette is50)分析[17].1.6X射线衍射能谱分析采用X射线衍射仪(XRD)[18]对1.3.3中制备的矿化产物进行分析,用Cu Kα射线检测,设置参数为36kV 和20mA,扫描角度为10°~70°,步长和扫描速度分别为0.02°和0.5°(2θ)/min.1.7数据处理Cd2+的固定去除率计算:R(%)=(C0-C t)/C0×100% (1) 式中:R为Cd2+去除率,%;C0为空白组Cd2+浓度,mg/L;C t为实验组Cd2+浓度, mg/L.采用Origin 9.1软件和Excel 2019进行数据处理、作图,XRD采用MDI Jade6软件分析,数据显著性分析用SPSS 22.0进行统计检验.2结果与分析2.1碳酸盐矿化菌生长过程中脲酶活性,产CO32-浓度及pH值变化不同种类的碳酸盐矿化菌代谢产生的脲酶活性存在较大差异.如图1a所示,3种菌接种至培养基中,最初生长较缓慢,脲酶活性相对较弱, P.stutzeri脲酶活性增加缓慢,24h后逐渐趋于稳定,B. subtilis脲酶活性在第72h达到最高;B. pasteurii前8h脲酶活性迅速增加,第24h达到最高,随后出现一定程度的下降,第48h后逐渐趋于稳定.3种碳酸盐矿化菌生长过程中产生CO32-浓度变化如图1b所示.B. pasteurii产CO32-的能力最强,P.stutzeri和B. subtilis相对较弱.培养初期12h, B. pasteurii产生CO32-的浓度增加很快,48h时达到1735.18mg/L,之后CO32-浓度趋于稳定.P. stutzeri 和B. subtilis在12h后CO32-的浓度开始增加,但增速较为缓慢,72h后均趋于稳定,产生的CO32-浓度分别为588.19和 661.72mg/L.脲酶活性[mmol/(L⋅min)]12期朱广森等:微生物诱导碳酸盐沉积介导的Cd 2+固定 5915C O 32-浓度(m g /L )时间(h)0 8 16 24 3240 48 56 64 72p H 值时间(h)图1 3种碳酸盐矿化菌生长过程中脲酶活性、产CO 32-浓度及pH 值变化Fig.1 Changes in urease activity, CO 32- concentrations and pH during the growth of three carbonate mineralized bacteria3种碳酸盐矿化菌生长过程中pH 值变化如图1c 所示.P . stutzeri 生长过程中pH 值虽在不断升高,但上升较为缓慢,至生长平稳期时pH 值稳定在8.23左右.B.subtilis 在0~8h 出现pH 值降低的现象,可能是由于在最初生长过程中产生乳酸等酸性物质导致其pH 值下降,培养8h 后开始逐渐升高.培养24h 时pH 值上升到8.39,之后逐渐趋于平稳状态,最终pH 值稳定在9.06左右.B. pasteurii 生长过程中pH 值升高迅速,4h 时可达8.67,培养8h 可达9.25,24h 后达到最高pH 值9.52. 2.2 碳酸盐矿化菌对Cd 2+的耐受性如图2所示,随着Cd 2+浓度升高,碳酸盐矿化菌生长受到一定抑制作用.P . stutzeri 对Cd 2+的耐受性较强,Cd 2+浓度0~1mg/L 时对其生长有一定促进作用, Cd 2+浓度1~3mg/L 时对其生长影响不显著,Cd 2+浓度5~10mg/L 时,细菌生长减缓,Cd 2+浓度15mg/L 时细菌生长延滞期达到12h,对数生长期相对缩短.B. subtilis 生长过程中,Cd 2+浓度1mg/L 时对数生长期迟滞至24h 后,对数生长期也相对缩短,而Cd 2+浓度3~15mg/L 时该菌几乎不生长.B. pasteurii 生长过程中,Cd 2+浓度0~1mg/L 时其生长受抑制,对数生长期相对缩短,Cd 2+浓度1~3mg/L 时其生长受显著影响,对数生长期迟滞至36h 后,而当Cd 2+浓度5~15mg/L 时该菌几乎不生长.00.30.60.91.21.51.8O D600时间(h)00.40.81.21.62.02.4O D600时间(h)0O D600时间(h)图2 3种碳酸盐矿化菌对重金属Cd 2+的耐受性 Fig.2 The resistance of three carbonate mineralized bacteriato Cd 2+2.3 碳酸盐矿化菌对Cd 2+的固定如图3所示,3种碳酸盐矿化菌均对Cd 2+有较强的固定能力.P . stutzeri 对Cd 2+的去除24h 可达80.65%,120h 可达96.37%; B. subtilis 对Cd 2+的去除24h 即可达98.80%,120h 可达99.40%; B. pasteurii 对Cd 2+的去除24h 可达93.66%, 120h 达97.57%.此外,对照组出现Cd 2+含量降低,可能是由于有氧条件5916 中 国 环 境 科 学 41卷下空气中的CO 2进入含钙培养基中生成碳酸钙,从而与Cd 2+络合形成沉淀.0 16 32 48 64 80 96 112 128C d 2+浓度(m g /L )时间(h)图3 3种碳酸盐矿化菌对Cd 2+的去除效果Fig.3 Removal of Cd 2+ by three carbonate mineralized bacteria2.4 羟基磷灰石对碳酸盐矿化菌固定Cd 2+的影响如图4所示,加入羟基磷灰石(HAP),能够增强3种菌对Cd 2+的固定效果,HAP 联合碳酸盐矿化菌对Cd 2+的固定效果优于单独添加HAP.培养72h 时, HAP 与P . stutzeri , B. subtilis , B. pasteurii 联合作用对Cd 2+的固定去除率分别为85.8%, 87.9%和86.6%, 120h 时Cd 2+的固定率达95%以上,其中HAP 与B. pasteurii 联合固定效果最好.16324864 80 96 11212800.20.40.60.8 1.0C d 2+浓度(m g /L )时间(h)图4 羟基磷灰石联合碳酸盐矿化菌对Cd 2+的固定效果 Fig.4 Immobilization of Cd 2+ effect of hydroxyapatite combined with carbonate mineralization bacteria2.5 碳酸盐矿化菌固定Cd 2+的矿化产物特征分析图5中SEM 和EDS 分析表明,P . stutzeri 和B. subtilis 形成的矿化产物呈现不规则球状或网状结构,表面疏松多孔,并且大多聚集在菌体周围,部分产物附着在菌体表面,这一结构使其对重金属具有良好的物理吸附作用[19];B. pasteurii 的矿化产物结构致密,呈不规则的球状,并附着在整个菌体表面,有的将整个菌体包裹[20].EDS 结果显示,3种碳酸盐矿化菌的诱导矿化产物元素组成均含有C 、O 、Ca 和Cd,表明Cd 2+已被吸附固定.2µm24680102030405060计数(C P S )能量(keV)CdCaOC P . stutzeri2µm02468020406080100120计数(C P S )能量(keV)C OCa CdB. subtilis12期朱广森等:微生物诱导碳酸盐沉积介导的Cd 2+固定 59172µm2468102030405060计数(C P S )能量(keV)COCaCd B. pasteurii图5 3种碳酸盐矿化菌矿化产物SEM 和EDS 分析Fig.5 The SEM and EDS analysis of mineralization products by three carbonate mineralization bacteria4000 3500 3000 25002000 1500 1000500100200300400a P . stutaerib B. subtilisc B. pasteurii透过率(%)波数(cm -1)10561443697876图6 3种碳酸盐矿化菌矿化产物的FTIRFig.6 The FTIR of mineralization products by three carbonatemineralized bacteria球霰石方解石图7 3种碳酸盐矿化菌矿化产物的XRDFig.7 The XRD of mineralization products by three carbonatemineralized bacteria由图6可知,3种菌形成的矿物主要成分为碳酸盐.1443cm -1附近的吸收峰为CO 32-反对称伸缩吸收峰,1056cm -1附近吸收峰为碳氧对称伸缩吸收峰[21],876cm -1附近吸收峰为CO 32-面外弯曲吸收峰,697cm -1附近吸收峰为CO 32-面内弯曲吸收峰[16].图7中XRD 分析表明,P . stutzeri 在生长过程中产生的矿物主要是球霰石,B. subtilis 形成的矿化产物主要为方解石和球霰石,而B. pasteurii 矿化产物为方解石,XRD 特征吸收峰比较尖锐,表明形成矿物中含杂质少,结晶度好[22].经XRD 主要衍射峰与标准PDF 卡片检索发现,3种菌对Cd 2+固定的生物矿化产物主要存在形式为CdCO 3,其中B. pasteurii 的矿化产物以稳定的Ca 0.67Cd 0.33CO 3共结晶的形式产生共沉淀[23]. 3 讨论3.1 诱导碳酸盐沉积的微生物诱导碳酸盐沉积的微生物广泛分布在土壤和水体中.这些微生物主要包括芽孢杆菌属,假单胞菌属,克雷伯菌属和芽孢八叠球菌属等[24],而不同种类的微生物在诱导碳酸盐沉积过程中的产脲酶能力及脲酶活性,碳酸根的产量也存在较大差异[25].例如,Dhami 等[26]从印度安得拉邦石灰性土壤中分离到B. megaterium SS3 和B. thuringiensis ,其产脲酶分别为690和620U/mL,产生碳酸根能力分别为 1870和1670mg/L;Li 等[27]研究表明Terrabacter tumescens A12和B6的脲酶活性分别为14.81 和1.25U/mL.本研究选取的3种碳酸盐矿化菌,B. pasteurii 的脲酶活性最强[28],而P . stutzeri ,B. subtilis 相对较弱;脲酶活性的强弱从侧面反映出3种碳酸盐矿化菌产CO 32-能力的强弱,B. pasteurii 产CO 32-浓度最高可达1735.18mg/L,P . stutzeri 和B. subtilis 产生的CO 32-浓度分别为588.19和 661.72mg/L.3.2 微生物诱导碳酸盐沉积介导Cd 2+固定机制微生物诱导碳酸盐沉积可通过尿素水解[29]、光5918 中国环境科学 41卷合作用[30-31]、硫酸盐还原[32]、反硝化作用[33]等途径实现,其中微生物水解尿素诱导碳酸盐沉淀因其机理相对简单,过程消耗少,对环境友好等优点而被广泛研究.微生物诱导碳酸盐沉淀固定重金属的基本原理是,通过微生物代谢产生脲酶,水解尿素产生NH4+和CO32-,NH4+释放NH3使pH值升高,从而在碱性环境下,足够浓度的Ca2+与CO32-结合,在细菌表面形成CaCO3沉淀,进而将Cd2+以类质同象置换方式占据Ca2+位置而固定[34],或Cd2+与CaCO3形成共结晶的方式被固定[35].本研究中3种碳酸盐矿化菌在水解尿素过程中产生OH-会提高环境pH值和平衡碳酸氢盐,从而形成CO32-,重金属Cd2+与游离的CO32-形成CdCO3晶体,从而降低有效态Cd的含量,这与Fujita等[36]研究结果一致.也正是上述固定机制的存在,3种碳酸盐矿化菌对培养环境中的Cd2+表现出高效的去除效果.基于微生物诱导碳酸盐沉积固定Cd2+的机理,对3种碳酸盐矿化菌固定Cd2+的次生矿物进行表征.SEM分析发现3种菌产生的矿化产物主要位于菌体表面或者菌体周围,这可能是由于菌体表面带有更多的负电荷,有更多的非均相成核位点[37],使Ca2+被大量吸附在菌体表面,在CO32-和碱性环境中,会沉淀形成CaCO3结晶[38].FTIR研究证实在1443cm-1附近特征吸收峰为碳酸盐官能团的对称伸缩振动产生,在1100~1000cm-1范围内是碳氧伸缩键的特征吸收峰[39],纯方解石和球霰石的典型振动带处于876,848,745cm-1[40],本研究中的FTIR图谱呈现出相似的结果,也进一步表明3种菌形成的次生矿物主要是碳酸盐.XRD图谱分析显示, B. pasteurii产生的次生矿物中检测到方解石型CaCO3和Ca0.67Cd0.33CO3,而P. stutzeri的次生矿物主要是球霰石型CaCO3和CdCO3,B. subtilis的次生矿物为方解石型CaCO3,球霰石型CaCO3共存和CdCO3.含Cd 矿物形成的可能机制是,Cd2+通过同构取代Ca2+而形成CdCO3或是与方解石的固溶体[41-42],之前的研究报道中也出现过类似的化学成分和晶体形式[43].3.3 羟基磷灰石对碳酸盐矿化菌固定Cd2+的影响机制当培养环境中加入化学材料羟基磷灰石(HAP)时,在一定程度上会减缓微生物诱导碳酸盐沉积的速率,但HAP和菌体共存体系提高了Cd2+的去除效果.HAP(Ca10(PO4)6(OH)2)因其较大的比表面积,微溶于水,有良好的吸附特性,而被广泛应用于重金属治理.本研究中加入HAP,除自身结构会对Cd2+吸附外[44],其特殊的空间结构,良好的生物相容性亦可作为微生物的吸附剂[45],为菌体诱导碳酸盐沉淀提供成核载体.本研究中可能存在的机制为:根据沉淀溶解平衡,HAP会有一部分Ca2+ 溶解到环境中,以此作为钙源,被吸附在HAP上的菌体会诱导CaCO3沉淀,进而对Cd2+生物矿化固定.上述机制仅是根据本研究所做出的推断, HAP和菌体联合作用过程,以及对多种重金属的固定产物及其机理还需更深入细致的研究.4结论4.1碳酸盐矿化菌P. stutzeri, B. subtilis, B. pasteurii生长过程中,B. pasteurii d的脲酶活性和产CO32-能力最强,同时,3种菌的代谢均使培养体系pH 值升高.4.2Cd2+浓度在0~10mg/L范围时对P. stutzeri生长影响较小,而Cd2+浓度高于1mg/L时则会抑制B. subtilis和B. pasteurii的生长.P. stutzeri对Cd2+的耐受能力比B. subtilis和B. pasteurii强.4.33种碳酸盐矿化菌均对0~1mg/L Cd2+有较强的固定能力,120h时Cd2+的固定率可达95%以上;加入羟基磷灰石能够增强碳酸盐矿化菌对Cd2+的固定效果.4.43种碳酸盐矿化菌的诱导矿化产物是碳酸盐.P. stutzeri的主要产物是球霰石,B. subtilis的矿化产物为方解石和球霰石共存,而B. pasteurii矿化产物以方解石为主;Cd2+与Ca2+同晶置换以CdCO3的形式沉淀,部分以稳定的Ca0.67Cd0.33CO3共结晶的形式存在.参考文献:[1] Zang F, Wang S L, Nan Zhongren,et al. Immobilization of Cu, Zn, Cdand Pb in mine drainage stream sediment using Chinese loess [J].Chemosphere, 2017,181:83-91.[2] 唐永安,胡军,杨祥良,等.镉系量子点的生物毒性及相应机制 [J].化学进展, 2014,26(10):1731-1740.Tang Y A, Hu J, Yang X L, et al. Biotoxicity of cadmium-based quantum dots and the mechanisms [J]. Progress in Chemistry, 2014, 26(10):1731-1740.[3] 陈能场,郑煜基,何晓峰,等.《全国土壤污染状况调查公报》探析 [J].12期朱广森等:微生物诱导碳酸盐沉积介导的Cd2+固定 5919农业环境科学学报, 2017,36(9):1689-1692.Chen N C, Zheng Y J, He X F, et al. Analysis of the Report on the national general survey of soil contamination [J]. Journal of Agro- Environment Science, 2017,36(9):1689-1692.[4] 吴洋,练继建,闫玥,等.巴氏芽孢八叠球菌及相关微生物的生物矿化的分子机理与应用 [J]. 中国生物工程杂志, 2017,37(8):96- 103.Wu Y, Lian J J, Yan Y, et al. Mechanism and applications of bio- mineralization induced by Sporosarcina pasteurii and related microorganisms [J]. China Biotechnology, 2017,37(8):96-103.[5] Gadd G M. Bioremedial potential of microbial mechanisms of metalmobilization and immobilization [J]. Current Opinion in Biotechnology, 2000,11(3):271-279.[6] Han L J, Li J H, Xue Q, et al. Sun Poon. Bacterial-inducedmineralization (BIM) for soil solidification and heavy metal stabilization: A critical review [J]. Science of the Total Environment, 2020,746:140967.[7] 赵越,姚俊,王天齐,等.碳酸盐矿化菌的筛选与其吸附和矿化Cd2+的特性 [J]. 中国环境科学, 2016,36(12):3800-3806.Zhao Y, Yao J, Wang T Q, et al. Screening of carbonate- biomineralization microbe and its cadmium removal characteristics based on adsorption and biomineralization [J]. China Environmental Science, 2016,36(12):3800-3806.[8] 王新花,赵晨曦,潘响亮.基于微生物诱导碳酸钙沉淀(MICP)的铅污染生物修复 [J]. 地球与环境, 2015,43(1):80-85.Wang X H, Zhao C X, Pan X L. Bioremediation of Pb-pollution basedon microbially induced calcite precipitation [J]. Earth and Environment, 2015,43(1):80-85.[9] Jalilvand N, Akhgar A, Alikhani H A,et al. Removal of heavy metalszinc, lead, and cadmium by biomineralization of urease-producingbacteria isolated from Iranian mine calcareous soils [J]. Journal of SoilScience and Plant Nutrition, 2020,20(1):206-219.[10] Peng D H, Qiao S Y, Yao L, et al. Performance of microbial inducedcarbonate precipitation for immobilizing Cd in water and soil [J].Journal of Hazardous Materials, 2020,400:123116.[11] Kumari D, Pan X L, Lee D J,et al. Immobilization of cadmium in soilby microbially induced carbonate precipitation with Exiguobacteriumundae at low temperature [J]. International Biodeterioration & Biodegradation, 2014,94:98-102.[12] Achal V, Pan X L, Zhang D Y, et al. Bioremediation of Pb-Contaminated soil based on microbially induced calcite precipitation [J]. Journal of Microbiology and Biotechnology, 2012,22(2):244-247. [13] 李劲,杨玉蓉,朱广森,等.Shewanella oneidensis MR-1和Shewanella putrefaciens对V(Ⅴ)的还原及其影响因素 [J]. 中国环境科学, 2020,40(1):414-421.Li J, Yang Y R, Zhu G S, et al. The reduction of V(V) by Shewanellaoneidensis MR-1and Shewanella putrefaciens and influencing factors [J]. China Environmental Science, 2020,40(1):414-421.[14] 李成杰,魏桃员,季斌,等.不同钙源及Ca2+浓度对MICP的影响 [J].环境科学与技术, 2018,41(3):30-34.Li C J, Wei T Y, Ji B,et al. Study on MICP affected by different calcium sources and Ca2+ concentrations [J]. Environmental Science &Technology, 2018,41(3):30-34. [15] DZ/T 0064.49-1993 地下水质分析方法第49部分:滴定法测定碳酸根、重碳酸根和氢氧根离子 [S].DZ/T 0064.49-1993, Methods for analysis of groundwater quality – Part 49: Determination of carbonate, bicarbonate ions, hydroxy by titration [S].[16] 许凤琴.蒙脱石-碳酸盐矿化菌对Sr2+,Pb2+的联合滞固研究 [D]. 西南科技大学, 2016.Xu F Q. The combined immobilization of Sr2+, Pb2+ by montmorillonite and carbonate mineralization bacteria [D]. Southwest University of Science and Technology, 2016.[17] Lin W T, Huang Z, Li X Z, et al. Bio-remediation of acephate–Pb(Ⅱ)compound contaminants by Bacillus subtilis FZUL-33 [J]. Journal of Environmental Sciences, 2016,45(7):94-99.[18] Kim J H, Lee J Y. An optimum condition of MICP indigenous bacteriawith contaminated wastes of heavy metal [J]. Journal of Material Cycles and Waste Management, 2019,21(2):239-247.[19] Liu R L, Lian B. Immobilization of Cd(II) on biogenic and abioticcalcium carbonate [J]. Journal of Hazardous Materials, 2019,378: 120707.[20] Anbu P, Kang C H, Shin Y J, et al. Formations of calcium carbonateminerals by bacteria and its multiple applications [J]. SpringerPlus, 2016,5(1):10-12.[21] Chen X, Achal V. Effect of simulated acid rain on the stability ofcalcium carbonate immobilized by microbial carbonate precipitation [J]. Journal of Environmental Management, 2020,264:110419.[22] Liu R L, Guan Y, Chen L, et al. Adsorption and desorptioncharacteristics of Cd2+ and Pb2+ by micro and nano-sized biogenic CaCO3 [J]. Front in Microbiology, 2018,9:41.[23] 周野,王一莹,李哲,等.氧化木糖无色杆菌(Achromobacterxylosoxidans) LAX2对Cu, Pb和Cd复合污染土壤的生物矿化修复研究 [J]. 环境科学学报, 2018,38(11):4497-4504.Zhou Y, Wang Y Y, Li Z, et al. The remediation of complex Cu, Pb and Cd polluted soil through biomineralization by Achromobacte rxylosoxidans LAX2 [J]. Acta Scientiae Circumstantiae, 2018,38(11): 4497-4504.[24] 党政,代群威,赵玉连,等.生物矿化在重金属污染治理领域的研究进展 [J]. 环境科学研究, 2018,31(7):1182-1192.Dang Z, Dai Q W, Zhao Y L, et al. Research progress of biomineralization in the treatment of heavy metal contamination [J].Research of Environmental Sciences, 2018,31(7):1182-1192.[25] 李成杰.产脲酶菌诱导碳酸钙沉积及对重金属Cd2+/Pb2+的去除研究[D]. 武汉科技大学, 2018.Li C J. Bioremediation of Cd2+/Pb2+ and calcium carbonate precipitation induced by urease producing bacteria [D]. Wuhan University of Science and Technology, 2018.[26] Dhami N K, Reddy M S, Mukherjee A. Biomineralization of calciumcarbonate polymorphs by the bacterial strains isolated from calcareous sites [J]. Journal of Microbiology and Biotechnology, 2013,23(5): 707-714.[27] Li M, Cheng X H, Guo H X,et al. Biomineralization of carbonate byTerrabacter tumescens for heavy metal removal and biogrouting applications [J]. Journal of Environmental Engineering, 2015,142(9): C4015005.5920 中国环境科学 41卷[28] 裴迪,刘志明,胡碧茹,等.巴氏芽孢杆菌矿化作用机理及应用研究进展 [J]. 生物化学与生物物理进展, 2020,47(6):467-482.Pei D, Liu Z M, Hu B R, et al. Progress on mineralization mechanism and application research of Sporosarcina pasteurii [J]. Progress in Biochemistry and Biophysics, 2020,47(6):467-482.[29] Cheng L, Cord-Ruwisch R. Selective enrichment and production ofhighly urease active bacteria by non-sterile (open) chemostat culture [J].Journal of Industrial Microbiology and Biotechnology, 2013, 40(10):1095-1104.[30] Bundeleva I A, Shirokova L S, Bénézeth P, et al. Calcium carbonateprecipitation by anoxygenic phototrophic bacteria [J]. Chemical Geology, 2012,291:116-131.[31] McConnaughey T A, Whelan J F. Calcification generates protons fornutrient and bicarbonate uptake [J]. Earth Science Reviews, 1997, 42(1):95-117.[32] Deng S C, Dong H L, Lv G, et al. Microbial dolomite precipitationusing sulfate reducing and halophilic bacteria: Results from Qinghai Lake, Tibetan Plateau, NW China [J].Chemical Geology, 2010,278 (3/4):151-159.[33] Zhu T T, Dittrich M. Carbonate precipitation through microbialactivities in natural environment, and their potential in biotechnology:a review [J]. Frontiers in Bioengineering and Biotechnology, 2016,4:4.[34] 王茂林,吴世军,杨永强,等.微生物诱导碳酸盐沉淀及其在固定重金属领域的应用进展 [J]. 环境科学研究, 2018,31(2):206-214.Wang M L, Wu S J, Yang Y Q, et al. Microbial induced carbonate precipitation and its application for immobilization of heavy metals: a review [J]. Research of Environmental Sciences, 2018,31(2):206-214.[35] Fonyo C, Machado C C, Meganck R, et al. Microbiologically InducedCalcite Precipitation biocementation, green alternative for roads – is this the breakthrough? A critical review [J]. Journal of Cleaner Production, 2020,262:4-6.[36] Fujita Y, Taylor J L, Gresham T L T,et al. Stimulation of microbialurea hydrolysis in groundwater to enhance calcite precipitation [J].Environmental science & technology, 2008,42(8):3025-3032.[37] Rajasekar A, Moy C K S, Wilkinson S. MICP and advances towardseco-friendly and economical applications [J]. IOP Conference Series:Earth and Environmental Science, 2017,78(1):012016.[38] K hadim H J, Ammar S H, Ebrahim S E. Biomineralization basedremediation of cadmium and nickel contaminated wastewater by ureolytic bacteria isolated from barn horses soil [J]. Environmental Technology & Innovation, 2019,14:100315.[39] Qian X Y, Fang C L, Huang M S, et al. Characterization offungal-mediated carbonate precipitation in the biomineralization of chromate and lead from an aqueous solution and soil [J]. Journal of Cleaner Production, 2017,164:198-208.[40] Trushina D B, Bukreeva T V, Kovalchuk M V,et al. CaCO vateritemicroparticles for biomedical and personal care applications [J].Materials Science & Engineering C, 2014,45:644-658.[41] Achal V, Pan X L, Zhang D Y. Bioremediation of strontium (Sr)contaminated aquifer quartz sand based on carbonate precipitation induced by Sr resistant Halomonas sp. [J]. Chemosphere, 2012,89(6): 764-768.[42] Bhattacharya A, Naik S N, K hare S K. Harnessing the bio-mineralization ability of urease producing Serratia marcescens and Enterobacter cloacae EMB19for remediation of heavy metal cadmium () [J]. Journal of Environmental Management, 2018,215:143Ⅱ-152. [43] 王瑞兴,钱春香,吴淼,等.微生物矿化固结土壤中重金属研究 [J].功能材料, 2007,(9):1523-1527,1530.Wang R X, Qian C X, Wu M, et al. Study on heavy metals in soil mineralized by bacteria [J]. Journal of Functional Materials, 2007,(9):1523-1527,1530.[44] Rötting T S, Cama J, Ayora C, et al. Use of caustic magnesia toremove cadmium, nickel, and cobalt from water in passive treatment systems: column experiments [J]. Environmental Science & Technology, 2006,40(20):6438-6443.[45] Piccirillo C, Pereira S I A, Marques A, et al. Bacteria immobilisationon hydroxyapatite surface for heavy metals removal [J]. Journal of Environmental Management, 2013,121:87-95.作者简介:朱广森(1991-),男,河南灵宝人,安徽农业大学硕士研究生,主要研究方向为环境生物技术.发表论文4篇.。
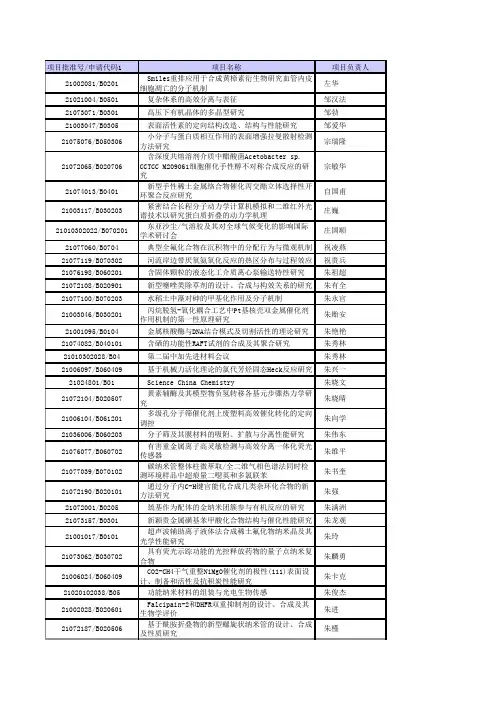
项目批准号/申请代码1项目名称项目负责人21002081/B0201 Smiles重排应用于合成黄樟素衍生物研究血管内皮细胞凋亡的分子机制左华21021004/B0501 复杂体系的高效分离与表征 邹汉法21073071/B0301 高压下有机晶体的多晶型研究 邹勃21003047/B0305 表面活性素的定向结构改造、结构与性能研究 邹爱华21075076/B050306 小分子与蛋白质相互作用的表面增强拉曼散射检测方法研究宗瑞隆21072065/B020706 含深度共熔溶剂介质中醋酸菌Acetobacter sp.CCTCC M209061细胞催化手性醇不对称合成反应的研究宗敏华21074013/B0401 新型手性稀土金属络合物催化丙交酯立体选择性开环聚合反应研究自国甫21003117/B030203 紧密结合长程分子动力学计算机模拟和二维红外光谱技术以研究蛋白质折叠的动力学机理庄巍21010302022/B070201 东亚沙尘/气溶胶及其对全球气候变化的影响国际学术研讨会庄国顺21077060/B0704 典型全氟化合物在沉积物中的分配行为与微观机制 祝凌燕21077119/B070302 河流岸边带厌氧氨氧化反应的热区分布与过程效应 祝贵兵21076198/B060201 含固体颗粒的液态化工介质离心泵输送特性研究 朱祖超21072108/B020901 新型噻唑类除草剂的设计、合成与构效关系的研究 朱有全21077100/B070203 水稻土中藻对砷的甲基化作用及分子机制 朱永官21003046/B030201 丙烷脱氢-氧化耦合工艺中Pt基核壳双金属催化剂作用机制的第一性原理研究朱贻安21001095/B0104 金属核酸酶与DNA结合模式及切割活性的理论研究 朱艳艳21074082/B040101 含硒的功能性RAFT试剂的合成及其聚合研究 朱秀林21010302028/B04 第二届中加先进材料会议 朱秀林21006097/B060409 基于机械力活化理论的氯代芳烃固态Heck反应研究 朱兴一21024801/B01 Science China Chemistry 朱晓文21072104/B020507 黄素辅酶及其模型物负氢转移各基元步骤热力学研究朱晓晴21006104/B061201 多级孔分子筛催化剂上废塑料高效催化转化的定向调控朱向学21036006/B060203 分子筛及其膜材料的吸附、扩散与分离性能研究 朱伟东21076077/B060702 有害重金属离子高灵敏检测与高效分离一体化荧光传感器朱维平21077039/B070102 碳纳米管整体柱微萃取/全二维气相色谱法同时检测环境样品中超痕量二噁英和多氯联苯朱书奎21072190/B020101 通过分子内C-H键官能化合成几类杂环化合物的新方法研究朱强21072001/B0205 巯基作为配体的金纳米团簇参与有机反应的研究 朱满洲21073157/B0301 新颖贵金属磺基苯甲酸化合物结构与催化性能研究 朱龙观21001017/B0101 超声波辅助离子液体法合成稀土氟化物纳米晶及其光学性能研究朱玲21073062/B030702 具有荧光示踪功能的光控释放药物的量子点纳米复合物朱麟勇21006024/B060409 CO2-CH4干气重整NiMgO催化剂的极性(111)表面设计、制备和活性及抗积炭性能研究朱卡克21020102038/B05 功能纳米材料的组装与光电生物传感 朱俊杰21002028/B020601 Falcipain-2和DHFR双重抑制剂的设计、合成及其生物学评价朱进21072187/B020506 基于酰胺折叠物的新型螺旋状纳米管的设计、合成及性质研究朱槿21004025/B040606 嵌段共聚物乳液液滴的界面不稳定现象机理与微结构调控朱锦涛21006054/B060702 新型齐聚物糖基水凝胶因子设计合成及其凝胶行为 朱金丽21071014/B0101 氮化物以及氮氧化物可见光光催化剂的制备与性能研究朱鸿民21077137/B0704 SPME原位采样技术监测土壤-农作物系统的持久性有机污染物朱芳21076234/B060203 火灾下热功能含湿织物的干燥收缩分形分析及湿热传递模型朱方龙21072151/B020706 生物催化不对称羰基还原胺化反应的探索 朱敦明21066014/B0608 内生真菌石杉碱甲生物合成途径及代谢调控研究 朱笃21077049/B070203 土壤中煤源颗粒对有机污染物的吸附、解吸研究朱东强21074056/B040606 利用介电松弛谱研究酶电极中导电高聚物与生物大分子界面微结构及电荷传输朱丹21001033/B010701 介孔材料-核酸适体的组装及在药物控释技术中的应用研究朱春玲21061003/B010303 含氮、氧配位供体原子的有机配体及其配合物的合成、结构及性能研究朱必学21002069/B0202 双核金属配合物“协同”活化惰性C-H键朱柏林21002048/B0206 新型黄酮类肿瘤血管阻断剂的合成与生物活性研究 周中振21073152/B030606 电催化过程中低覆盖度吸附态中间体的原位红外光谱检测周志有21073096/B0302 无机纳米薄片/条带的计算设计与嵌锂性能周震21071051/B010401 扭曲度可调型类血红素铁卟啉的合成及其复合物性能研究周再春21074134/B040502 PEO树枝齐聚物嵌段共聚物的自组装与结晶行为研究周云春21071143/B010303 纳米尺度金属有机骨架材料的设计合成及其催化构效关系研究周有福21075114/B0511 膜保护配位聚合物微固相萃取技术在多溴联苯醚预富集和分析中的应用周友亚21006129/B060409 微乳液中纳米粒子定点负载构建新型钯整体式催化剂的研究周永华21074069/B040502 超支化聚合物的支化拓扑结构和性能关系研究周永丰21003075/B030606 DMFC电催化剂载体材料氮掺杂石墨烯的基础与应用研究周盈科21062003/B0207 中药桑白皮对HIV-1 LTR启动子活性的调控作用研究周英21076142/B060304 氧化铁/一氧化碳循环分解水制氢基础研究 周亚平21001065/B0103 芴基发光金属-有机骨架材料的设计合成和性能研究周馨慧21076036/B060702 砜和手性亚砜的选择性氧化合成及其机理研究 周新锐21003115/B030201 太阳能光催化制氢材料吸光机制的理论研究周新21073173/B030402 乙烯及衍生物分子的电离能、键能及解离动力学研究周晓国21003110/B030607 氧化-还原分子电子输运的STM裂结技术和电化学超快循环伏安法研究周小顺21072132/B020104 水溶性金属配合物催化水相偶联反应的研究 周向葛21072155/B020702 可诱导核酸交联剂的设计、合成及生物活性研究 周翔21002016/B020402 醉鱼草属植物杀虫活性成分及其作用机制研究周霞21003088/B030605 现场表面振动光谱法研究锂离子电池电极和离子液体的界面结构周尉21003005/B030502 介电弛豫法对电流变液动态界面多重极化特性的解析研究周威21010302016/B06反应与分离系统的耦合与集成周涛21007059/B0707 长三角毗邻海域有机氯农药随食物链(网)的迁移转化及生态风险周珊珊21037002/B070203 土壤污染微界面过程及其分子诊断与调控原理 周启星21032003/B020104 高效不对称催化反应及其在天然产物和手性药物合成中的应用研究周其林21074025/B0404 灵芝有效降糖天然大分子提取物的组成结构及在糖尿病治疗中的作用机理周平21077130/B0703 甲基对硫磷及4-硝基酚污染土壤的微生物修复机理研究周宁一21074080/B040101 多种拓扑结构的环型偶氮苯聚合物的设计、合成和性能研究周年琛21006036/B0612 碱木质素的溶液行为及其化学修饰模型物在固液界面的吸附机理研究周明松21072218/B021102 荧光编码标记分子(“分子条形码”)概念性验证 周明21004053/B040501 生物相容超分子聚合物的合成及其在磁共振造影剂的应用周密21001045/B0104 硒通过调节内质网应激发挥类胰岛素作用的研究 周军21002118/B0208 有机小分子催化的不对称环加成反应机理研究 周静21002005/B020702 应用新型质谱技术研究人类端粒G-四链体DNA的结构特性及其与小分子的结合位点周江21063004/B030105 在线灌注活细胞的P-31核磁共振波谱学特征研究 周建威21003034/B030105 基于表面等离子体结构的WGM/SERS生物传感特性研究周吉21006022/B060802 定向嵌段共聚制备温度和pH双重敏感聚氨酯基智能材料周虎21001038/B010403 稀土纳米材料促小鼠成骨细胞增殖作用的分子机制研究周国强21027006/B040608 与显微结构研究集成的冷热台型高速扫描高灵敏热分析仪的研制周东山21071052/B010801 β-环糊精衍生物萃取分离对映体过程中的手性识别机理及构效关系周从山21073150/B030302 氨基多羧酸高效高选择性络合催化降解研究 周朝晖21001032/B010303 基于肟类桥连配体的单分子磁体的合成与性质研究 周爱菊21081220312/B060104 第四届中美“能源与环境:化学工程师的机遇与挑战”化工研讨会仲崇立21064004/B0403 壳聚糖新型衍生物作为农业杀菌剂研究 钟志梅21002104/B020203 基于共轭环状金属配合物的线型分子导线的合成与研究钟羽武21076195/B0608 大肠杆菌K5产肝素前体heparosan代谢控制研究 钟卫鸿21076194/B060409 基于Baylis-Hillman反应的手性膦杯芳烃的合成及催化性能研究钟为慧21006103/B060903 燃料电池用掺杂型非铂催化剂制备及其构效关系研究钟和香21074012/B0402 基于聚对苯撑乙炔分子链内环化反应构筑新型共轭高分子支俊格21073110/B030204 量子相空间动力学:轨线——密度函数方法 郑雨军21072067/B020506 基于有机分子聚集诱导发光特性的手性识别研究 郑炎松21073095/B030106 离子液体对TiO2的成核、物相及形貌的影响研究 郑文君21071062/B010403 新型有机硒化合物协同TRAIL诱导肿瘤细胞凋亡的分子机制研究郑文杰21003053/B0301 螺旋体为前体的配位聚合物的组装与动态组合化学库的建立郑盛润21077011/B070302 苯二氮类镇静催眠药物在A2/O工艺中的强化净化 郑少奎21037003/B07 典型工业过程中持久性有机污染物生成机制与控制原理郑明辉21006073/B060407 基于扩散层原位生长纳米碳纤维的燃料电池膜电极组件研究郑俊生21073129/B030608 锂离子电池中正负电极间的相互作用与机理研究 郑洪河91022011/B0103 含[MCuxSy] (M = Mo, W)功能基元的簇合物的合成及高阶非线性光学性能研究郑和根21071033/B0111 枝状结构硅纳米线的合成与高灵敏度生物传感器的制备郑耿锋21075085/B0503 多模式薄膜化学蒸气发生:装置、反应体系及应用 郑成斌21073228/B030105 硼掺杂TiO2光催化活性增强机制的固体NMR和量子化学计算研究郑安民21007062/B0701 近海海洋环境中PBDEs迁移转化机制研究 赵宗山21073235/B030301 FCC汽油选择性加氢脱硫新型L沸石基催化剂及反应机理研究赵震21006031/B060306 新型的ZIFs晶体膜的制备及其分离CO2/N2机理 赵祯霞21062024/B020102 DNA聚合酶抑制剂(+)-Aphidicolin全合成研究 赵元鸿21074081/B040101 多组分星形和接枝聚合物的合成及性能研究 赵优良21073114/B030301 氧化铝载体在含水加氢体系中的水合脱结构研究 赵永祥21005059/B050105 基于多功能免疫磁珠和微流控芯片的痕量循环肿瘤细胞检测赵永席91022022/B0211 低维有机微晶材料的设计、合成与光电性能研究 赵永生21074091/B040308 分子印迹中空纤维膜的制备及其对手性分子吸附与拆分性能的智能化调控赵义平21073146/B030203 具有多桥的复杂分子体系中的电子转移理论和应用 赵仪21075101/B0503 以纳米颗粒为载体的分子药物控制释放和监测体系研究赵一兵21003039/B030802 离子液体与有机分子的相互作用及其对若干化学反应选择性的影响赵扬21062030/B020506 新型桥联环糊精/DABO类HIV逆转录酶抑制剂包结配合物的超分子体系研究赵焱21066005/B060903 新型稀土/金属离子掺杂Ta2O5纳米电极材料的制备及在燃料电池中的应用赵彦宏21073216/B030503 氢/氘分子在微孔碳材料上的动力学量子筛分离机理的研究赵学波21076059/B061103 基于甘油氢解和CO2醇解反应过程集成的碳酸丙烯酯新绿色合成反应研究赵新强21005060/B050102 固定化功能蛋白质取向及构象表征和调控色谱新方法赵新锋21076040/B060905 锌离子调节酵母菌乙醇耐性的分子机制和胁迫耐受酵母的构建赵心清21007063/B0701 有机基团修饰的磁性纳米材料在复杂环境水样品分析中的应用研究赵晓丽21005091/B050206 分子印迹阵列传感器的研制及其在食品安全快速检测中的应用赵晓娟21073033/B0305 基于光子晶体编码微球的蛋白质SERS检测及其应用 赵祥伟21001043/B010101 具有超顺磁性的小粒径大孔径介孔药物载体的制备及性能研究赵文茹21075065/B050206 维持酶蛋白自然构型的新型生物传感器电极的研制及应用赵文波21072029/B020601 以天然生物碱冬青生菌素H为先导的药物设计、合成及生物活性研究赵圣印21075125/B0509 粒径单分散双孔型PGMA微球基质及小分子药物配基高效亲和色谱体系的研究赵睿21007035/B070102 竹炭样品处理技术分离检测痕量环境污染物及其作用机理研究赵汝松21061016/B010303 异核低维分子基磁性材料的合成和相关性质研究 赵琦华21062018/B0207 樟叶越橘中熊果苷咖啡酰基转移酶的基因克隆与功能分析赵平21077094/B0707 高污染电子拆解区多氯联苯孕产妇污染负荷及对胎盘功能影响的分子机理赵美蓉21072056/B020104 新型双官能有机膦催化剂的合成及应用 赵梅欣21076140/B060203 化学-生物耦合膜反应器的构建及其对水体中多氯联苯原位去除机理研究赵林21002057/B021102 基于氮杂杯芳烃的配位自组装及性能研究 赵亮21003158/B0302 气相过渡金属离子与有机分子若干基本反应的理论研究赵联明21062028/B020405 嗜盐放线菌新物种抗肿瘤活性代谢产物研究 赵立兴21003011/B030204 常温质子迁移反应分子力场的设计开发与应用研究 赵立峰21072068/B020703 分子设计的藻胆蛋白的光动力学效应及其光疗作用探索赵开弘21072164/B020102 二苯乙烯-苯丙木脂素类似物的不对称合成及其生理活性研究赵静峰21003113/B0302 TiO2 表面湿电子态及光化学反应的理论研究 赵瑾21073028/B030706 具有超长激发态寿命的环铂室温磷光染料的分子设计、合成、光物理性质与应用研究赵建章21011130154/B020506 检测糖类分子的模块化手性荧光分子探针的研究 赵建章21077132/B070303 假单胞菌ND6菌株高效降解萘和萘胁迫应答的分子机理赵化冰21077001/B070302 耦合离子交换功能的高分子絮凝剂及其去除水中小分子溶解性有机物的研究赵华章21006002/B060204 分子层面多元体系内纳微结构药物颗粒的自聚体构建机理研究赵宏21074058/B040101 侧链或侧基可控断裂的梳型聚合物的制备 赵汉英21001089/B0113 基于单分子转动产生铁电性的有机铁电体的合成与性能测试赵海霞21077077/B0703 电吸附调控-光催化氧化降解不透光污染体系的方法与协同机制赵国华21072115/B0206 以AKT为靶标的新型三取代吲哚衍生物的设计、合成及抗肿瘤活性研究赵桂森21005087/B0501 卷烟烟气中主要杂环胺类化合物的体内代谢研究 赵阁21032006/B02 含氟有机化合物的合成、反应规律及应用研究 赵刚21003071/B030301 纳米-亚纳米复合结构贵金属催化剂的合成与表征 赵丹21072117/B021101 新型硼桥联梯形pi-共轭分子材料的设计合成及性能研究赵翠华21075122/B0501 基于代谢组学的转基因植物的安全性评价研究 赵春霞21076093/B060804 中国红豆杉细胞合成多乙酰基紫杉烷的代谢规律及调控机制赵春芳21076219/B061103 高纯纳米Al13溶胶制备大孔拟薄水铝石及其转化机制研究赵长伟21073035/B030201 负载型前过渡金属氧化物团簇模型催化剂构效关系的理论研究章永凡21072096/B020901 新型烯丙基香豆素类分子的设计与合成及其抗菌活性研究章维华21076052/B060702 糖分子内P、N手性配体的设计、制备及在不对称合成中的应用章鹏飞21072173/B020706 基于双AAO酶的手性汇聚法研究 张子张21073224/B0305 基于金属纳米颗粒-氧化石墨烯复合体系的表面增强拉曼基底的构筑及其用于细胞与药物相互作用研究 张智军21002051/B0201 炔丙基全碳1,3-偶极子在构建呋喃并杂环衍生物中的应用张志国21076126/B060301 强极性流体的气液相平衡行为及其应用 张志刚21077043/B070301 基于转化频率的碳烟催化燃烧机制张昭良21073162/B030602 纳米结构CeO2薄膜的光助阳极电沉积机制及其腐蚀电化学行为研究张昭21072042/B020101 新型环状过氧化物的合成方法研究张占辉21006075/B0603 微波辅助离子液体提取天然产物有效成分的传质机理研究张越非21076143/B060306 磷酸促进型掺锆二氧化硅/聚合物杂化膜及性能的研究张裕卿21071096/B010303 配合物嫁接的具有"二合一"功能的纳米复合材料 张玉良21072169/B0202 过渡金属催化的C-O键活化反应研究张玉红21074020/B040101 聚丙烯腈基碳纳米微球的可控制备及其基础问题研究张幼维21075102/B050303 金银纳米粒子增强室温磷光机理及其应用研究 张勇21001097/B010701 陀螺状磷酸亚铁锂/碳纳米管复合材料的结构调控及电化学特性研究张勇21066009/B061201 富氧增强褐煤燃烧性能及其在水泥熟料煅烧中的应用研究张永锋21005020/B0505 介孔纳米材料富集质谱鉴定糖肽新方法用于大肠癌血清糖肽组的研究张莹21076207/B060806 细胞间质体外模拟系统的构建及物质传递基本规律的研究张英21002092/B020405 白蚁共生放线菌的抗菌成分研究张应烙21073003/B030106 石墨烯表面化学修饰以及量子尺寸效应研究 张艳锋21005086/B050202 环介岛等温基因扩增过程的电化学信息研究张旭志21075132/B0504 14N核磁共振研究赖氨酸三甲基化蛋白质的相互作用动力学张许21074141/B040607 烯烃嵌段共聚物及其与聚烯烃共混体系的结构与性能关系研究张秀芹21010402027/B030201 过渡金属二茂夹心卤化物的国际合作研究 张秀辉21075029/B0502 基于分子印迹聚合物固相微萃取β-兴奋剂电化学传感器的研究张修华21001103/B0107 电沉积制备石墨烯/导电聚合物复合薄膜电极材料及其电容特性张熊21076030/B060306 优先渗透分离CO2的ZIF型金属有机骨架结构类分子筛膜的设计制备研究张雄福21076188/B061102 低温氮等离子体改性碳纳米管吸附二氧化碳的基础研究张兴旺21002117/B020601 靶向Mcl-1蛋白的抗肿瘤药物先导化合物的优化改造 张兴龙21074106/B0401 基于CO2/环氧化物可控共聚技术合成可降解的聚合物分子刷张兴宏21002070/B0201 钯催化多组分环化反应合成含硫杂环化合物的研究 张兴国21005010/B050304 LED诱导化学发光适配体传感器张信凤21077070/B070701 丁二烯代谢产物1-氯-3-丁烯-2-醇和1-氯-3-丁烯-2-酮与DNA碱基反应的研究张新宇21073061/B030606 纳米碳纤维的微结构及表面性质与其电催化活性的内在规律研究张新胜21027013/B0505 小型质谱仪器关键技术研究张新荣21072002/B020401植物甾醇单体氧化物的制备及其细胞毒性的研究张欣21007025/B070403 从芳香烃受体(AhR)的基因型研究卤代芳烃污染物的鸟类种间敏感差异性张效伟21010302032/B0509 2010 年第四届生物分析、生物医学工程与纳米技术国际会议(ISBBN 2010)张晓兵21072025/B0211 新型甲基酮桥连的给体-共轭-受体类分子电子开关的研究张晓安21003079/B0306 染料敏化太阳能电池中染料的分子设计、合成与光电性能研究张晓21073001/B030607 金属基复合材料电极的构建及其在肼类化合物中的检测应用张小俊21036007/B061201 离子液体及其在清洁化工中的应用基础研究 张香平21007066/B070403 六溴环十二烷3种异构体在鱼体内代谢过程研究 张娴21071030/B01 介电可调的有机-无机杂化包合物 张闻21076130/B060805 皮胶原-有机交联剂反应机理的计算化学研究 张文华21075041/B0502 半导体复合纳米材料光电分析方法在乙酰胆碱酯酶活性检测中的应用研究张文21076012/B060306 膜吸收近膜壁面处的传质行为及其传质强化手段研究张卫东21007023/B070302 树脂基纳米M/Fe(0)的研制及其催化脱除硝酸根的研究张炜铭21003097/B030106 高效可控制备微/纳米条形码的新方法研究 张伟明21074035/B040501 新型特定结构功能性β-环糊精包结络合超分子聚合物的构建、形态和应用张伟安21075039/B050106 磁性纳米固定相毛细管电色谱分离系统研究 张维冰21074059/B040106 疏水单体在水中的RAFT聚合:壳-冠型空心微球反应器中的RAFT聚合研究张望清21003119/B030301 IrMOx双活性位催化剂CO选择氧化微观动力学研究 张万生21077134/B070201 城市大气碳质颗粒物综合测定及七组分来源研究 张烃21071007/B010404 钙化结节中钙磷酸盐的结构及其转变——内、外源性金属离子的作用张天蓝21081260197/B06 第三届两岸化学工程与产品工程高峰研讨会 张锁江21072143/B020101 低价稀土金属试剂在合成多烯及多官能团化合物中的应用研究张松林21075021/B050901 微流控体系下DNA-蛋白质相互作用的单分子检测技术研究张松21077095/B070101 环境水体中超痕量溴酸根的在线富集、聚焦和毛细管电泳分离分析研究张书胜21007006/B070302 膜曝气分离单级自养脱氮生物反应器过程控制和功能菌群研究张寿通21004048/B040309 聚丁二酸丁二醇酯的仿生改性及其仿细胞外层膜结构纳米胶束的研究张世平21001036/B010701 具有多种聚阴离子基团的锂离子电池正极材料的结构调控及脱嵌锂性能研究张森21072156/B020601 新型激酶抑制剂:8-羟基-2-芳基-1-异喹啉酮类的合成和抗肿瘤活性研究张三奇21004021/B040308 利用非氟嵌段聚合物和超临界二氧化碳制备纳米孔径的高分子功能化薄膜张锐21003144/B030204 汽车尾气催化剂Pt掺杂CaTiO3的自再生机理的计算研究张秋菊21006108/B060402 转鼓式生物浸出反应器的传递特性和放大规律研究 张庆华21002107/B021102 新型“类离子液体”软功能材料制备与性能研究 张庆华21003081/B030802 离子液体与极性溶剂间相互作用规律研究 张庆国21001061/B010303 具有化学传感功能的多孔发光MOFs材料的设计构筑及其检测大气中POPs的研究张庆富21074123/B0403 单个偶氮聚合物囊泡的光致可逆形变的放大研究 张其锦21062013/B0210 四氯化硅催化氢化制备三氯氢硅工艺及机理研究 张宁21006023/B060304 扩张床吸附原位提取中药有效成分的方法研究 张敏21007083/B0705 环境渐变过程实验模拟及初始浓度效应研究 张美一21005006/B0502 基于扫描电化学显微镜的指纹采集技术的研究 张美芹21006069/B060802 “智能”溶栓策略探索——多尺度分子动力学模拟 张麟21076176/B060306 含无机纳米水通道反渗透复合膜的结构设计与制备 张林21076107/B061202 微小空间反应器中生物激发合成沸石与纳米金属粒子的研究张利雄21071032/B010303 新型季铵盐功能化的金属有机骨架材料的合成、结构与性能研究张丽娟21027002/B0501 蛋白质样品多级预处理系统的研制 张丽华21073177/B0307 甲醛分子离子电子激发态的振动分辨光解离动力学研究张立敏21073069/B030304 自掺杂光催化剂的设计、制备及其可见光光催化性能研究张礼知21074152/B0404 含树枝化结构基元的新型聚多糖衍生物研究 张黎明21001004/B0107 功能氧化物/碳纳米纤维复合材料的制备和电催化性能研究张莉21075016/B0501 多维毛细管液相色谱-质谱联用新技术用于甲型H1N1流感病毒的研究张兰21002115/B0203 环状手性含氟亚砜亚胺、亚磺酰胺和磺酰胺的立体专一性合成及应用研究张来俊21003050/B030301 限域纳米空间内酸碱有机官能团分子的可控组装及其协同催化机理的研究张坤21076162/B0608 油包水乳化体系中新型交联酶聚集体的构建及其结构与性能张峻21074048/B040605 具有不对称构筑基元的微结构阵列及其各向异性光学性质张俊虎21076063/B061201 基于离子液体的氯硅烷催化反应-相控耦合分离研究 张军21006110/B060901 炭黑和气体组分在煤气化过程中的作用机理研究 张聚伟21004001/B040601 停流光谱技术研究阳离子聚合物/DNA络合与解络合动力学张璟焱21076144/B060404 气液固三相高剪切反应器性能与模型放大研究 张金利21001120/B010303 多孔金属多氮唑框架 张杰鹏21073191/B0301 基于非手性源的单手性材料的催化不对称结晶 张健21002062/B020601 STAT3选择性抑制剂的设计、优化及其功能研究 张健21004077/B040303 基于主-客体相互作用的三重化学响应性聚合物组装体:设计、构建及其药物传输性能研究张建祥21074063/B040401 微量填充生物可降解高分子共混体系的微结构与性能研究张建明21073053/B030803 桥联型双核稀土多取代芳香羧酸配合物结构与热分解反应机理及性能研究张建军21071025/B010303 异金属团簇化合物的合成和性能研究 张建军21071019/B010601 二元金属氨硼烷的制备、释氢机理及其在推进剂中应用研究张建国21061004/B010701 含Fe钨青铜结构化合物的结构调控与电磁特性研究 张辉21071021/B010902 锝-99m、氟-18、碘-125-VEGF多肽肿瘤显像剂的制备、体外评价及生物分布研究张华北21073055/B030802 溶液中无机阴离子在纳米晶表面上的复合与稳定化作用研究张虎成21073077/B0304 有机光电材料激子态动力学研究 张厚玉21071140/B0105 新型稀土纳米复合材料的合成、发光和磁性能的研究张洪杰21074071/B040609 透明质酸多糖自聚集物理水凝胶的形成机理和流变学研究张洪斌21071027/B0101 功能化的多金属氧酸盐组装分子磁性晶态材料的可控合成与性能研究张宏21007069/B0704 T-2毒素生物转化及毒性的分子机制研究 张红霞21005067/B050105 集成化多功能可控细胞操纵及分析微流控芯片的研制张何21073079/B030702 高双光子吸收截面有机半导体材料的设计与性质研究张浩力21004032/B040102 新型烯烃复分解催化剂的设计、合成以及在制备结构可控的高性能聚合物材料中的应用张浩21077030/B0704 固氮蓝藻修复多氯联苯污染水稻土的机理研究 张杭君21077102/B0706 大辽河流域代表性卤代阻燃剂的污染特征与演变趋势张海军21006066/B060409 CH4/CO2重整高抗积碳金属/炭材料催化剂的制备及机理研究张国杰21007002/B070301 三维有序大孔-介孔复合氧化物原位担载贵金属纳米粒子的可控制备及同时消除NOx和碳烟的催化性能研究张桂臻21081260019/B020402 中国西北部中药资源开发国际研讨会 张桂珍21076215/B060802 层析过程中界面上蛋白质结构及动态变化 张贵锋21071146/B0107 多酸基多元复合光电催化材料的设计,制备及其性能研究张光晋21075126/B0509 基于聚集荧光增强机理的化学/生物传感的研究 张关心21076095/B0608 慢消化淀粉与茶多酚对餐后血糖反应的协同作用 张根义21073105/B030505 基于alpha-烷氧基锌酞菁J聚集机理的酞菁光控自组装张复实21077120/B070304 含溴电子废物在超临界甲醇中的催化脱溴机制研究 张付申21064002/B040705 介孔材料环境下原位乳液聚合稳定性及其聚合物复合材料热学和力学性能研究张发爱21072226/B020601 基于小檗碱抗耐药真菌作用的小分子探针研究 张大志21003077/B030301 以废轮胎热解炭为载体的脱氢催化剂在有机液体储氢中的研究张翠21075129/B050901 病原体的超灵敏高通量单分子检测研究 张春阳21027007/B0506 电化学发光成像分析仪的研制 张成孝21005030/B0511 基于碳纳米管表面印迹技术的猕猴桃根中抗肿瘤活性成分分离及活性研究张朝晖21004080/B040303 组织诱导型可生物降解聚谷氨酸水凝胶支架材料制备及其在骨组织工程中的应用研究张超21077117/B070301 Pt/TiO2催化剂室温氧化甲醛的高活性机制研究及非贵金属化探索张长斌21005065/B050102 基于液滴技术的蛋白质组分离分析新方法 张博21077126/B070502 典型羟基多溴联苯醚拟/抗激素效应的H12定位选择机制及构效关系研究张爱茜21073087/B0303 多壁碳纳米管的结构缺陷及其自发氧化还原性能在催化反应中的作用研究张爱民21034004/B040101 大尺度螺旋聚合物的可控合成及其结构分析 张阿方21075077/B0503 痕量多溴联苯醚的表面增强拉曼光谱检测 占金华21072159/B0201 过渡金属催化下各类杂环化合物的新合成方法研究 詹庄平21076184/B060702 持久低表面能、环境友好含短氟碳链聚合物的分子设计与合成詹晓力。

北运河沉积物中氨氧化微生物的群落特征鲍林林;陈永娟;王晓燕【期刊名称】《中国环境科学》【年(卷),期】2015(035)001【摘要】采用T-RFLP、RT-qPCR和克隆测序等分子生物学技术,以氨单加氧酶基因(amoA)为分子标记,研究了北运河表层沉积物中氨氧化古菌(AOA)和氨氧化细菌(AOB)的群落多样性、丰度、系统发育及其与环境因子的响应关系.结果表明,沉积物中AOB的群落多样性和丰度均高于AOA,是北运河沉积物中氨氧化过程的主要功能微生物.沉积物中氨氧化微生物群落结构沿干流和支流存在明显的空间分异,而AOA的种类组成空间差异较小;沉积物的氨氮(NH4+)和硝态氮(NO3-+NO2-)是影响氨氧化微生物群落特征的主要因子,AOB对环境变化的敏感性更高;AOA和AOB 的amoA基因拷1贝数分别为1.32×105~1.91×106copies/g、5.39× 105~8.3×106copies/g.闸坝下游沉积物的氨氧化微生物丰度最高.系统发育分析表明,amoA基因序列多属于土壤/沉积物分支,较多AOB的克隆序列与土壤亚硝化螺菌属(Nitrosospira)的类群相似性可达98%.受污水处理厂退水的影响,部分amoA 基因序列与污水处理厂废水和活性污泥中发现的类群同源性高.污染物质来源、支流汇入和闸坝拦截对河流沉积物氨氧化微生物的群落特征影响显著.【总页数】11页(P179-189)【作者】鲍林林;陈永娟;王晓燕【作者单位】首都师范大学资源环境与旅游学院,北京100048;首都师范大学资源环境与旅游学院,北京100048;首都师范大学资源环境与旅游学院,北京100048;首都师范大学首都圈水环境研究中心,北京100048【正文语种】中文【中图分类】X172【相关文献】1.北运河沉积物中主要脱氮功能微生物的群落特征 [J], 鲍林林;王晓燕;陈永娟;张苓荣2.湛江湾沉积物中氨氧化微生物的丰度、多样性和分布特征 [J], 阳雯娜;董宏坡;侯庆华;李雁群;陈法锦;周欣;毛铁墙3.小型浅水湖泊表层沉积物氨氧化古菌群落结构特征 [J], 燕文明;刘凌;赵倩维;杨艳青4.翅碱蓬对盐沼沉积物微生物生物量及β-氨氧化细菌群落的影响——以双台河口为例 [J], 刘姣;刘长发;李盛德;李晋;陶韦;李璐瑶;马悦欣5.高原湖泊沉积物中反硝化微生物的群落特征——以包头南海湖为例 [J], 杨文焕; 石大钧; 张元; 李卫平; 敬双怡; 于玲红因版权原因,仅展示原文概要,查看原文内容请购买。

内蒙古农业大学博士学位论文31①取低速离心(4000 转/分)的瘤胃液4ml 加入偏磷酸与甲酸按3:1配置的混合液1ml,混匀,静置半小时以上,取离心管,内加入 2 克酸性吸附剂(无水硫酸钠:50%硫酸:硅藻土=30:1:20)和4ml 的31.25mM 的巴豆酸溶液,再加入上述混合瘤胃液1ml 摇匀,澄清,用1μl 的微量进样器取样品上机进行测定。
②上机条件:仪器为日本岛津GC-7A 气相色谱仪。
色谱柱为内径3mm长2m 的不锈钢柱,担体为10%FFAP 加1%磷酸的ChromosorbW 担体,柱温105 ℃,汽化室温度200℃,离子化室温度230℃,氮气流速为1.75kg/cm2,氢气压力为0.7kg/cm2,空气的压力为0.35 kg/cm2,灵敏度为10,衰减128。
1.6.5 原虫的计数采集的瘤胃液立刻用两层纱布过滤,取滤液1ml 加4ml MFS 染液(甲基绿福尔马林溶液)固定,静置30 分钟进行染色,然后用吸管(管口大于 1.0mm)将混匀的样品稀释液以连续不断的液流充满血球计数室,盖上盖玻片,在光学显微镜下进行镜检(100 倍)。
每个样品计数10 个大方格,上下各 5 个大方格,即四角4 个大方格和中央1 个大方格,其中四角每个大方格包含16个中方格,中央1 个大方格包括20 个小方格(可以折算成16 个中方格,因为16 个中方格的体积等于20 个小方格的体积),最后取平均数。
按下列公式计算纤毛虫数。
纤毛虫数/ml 16101000= 16 N×10×××D×= N×D×1000N:所有方格中纤毛虫的总数D:稀释倍数16:每个大方格中有16 个中方格10:共计数10 个大方格1000:1mm3换算为1ml 时的倍数MFS 溶液配方:将NaCl 8 克,甲基绿0.6 克,福尔马林溶液100ml 混合定容至1000ml 即可。

第28卷第12期2016年12月V ol. 28, No. 12Dec., 2016生命科学Chinese Bulletin of Life Sciences文章编号:1004-0374(2016)12-1493-54国家自然科学基金委员会生命科学部2016年度青年基金项目DOI: 10.13376/j.cbls/20161861微生物学细菌基因组歧化和遗传界限的产生:新物种形成的分子基础唐 乐哈尔滨医科大学我国沿海盐田嗜盐古菌胞外蛋白酶多样性研究侯 靖江苏大学反硝化细菌的鉴定及其与胆固醇降解相关蛋白、基因的初探丁 滨浙江中医药大学大连新港石油污染海域沉积物中厌氧微生物种群和功能基因多样性与氮源响应规律研究陈 超大连民族大学苏云金芽胞杆菌晶胞粘连表型菌株资源及形成机制的多样性研究王月莹华中农业大学粪产碱菌杀线虫活性物质挖掘及作用线虫方式研究鞠守勇华中农业大学冰川稀有低温细菌Cryobacterium物种多样性、分类学及冷适应性研究刘 庆中国科学院微生物研究所捕食青枯菌的粘细菌资源分离与功能评价李安章广东省微生物研究所环境因子对花生根瘤菌遗传多样性和分布的影响机制研究李 岩中国科学院烟台海岸带研究所家蚕病原细菌多样性、分布规律及其与蚕体共生菌关系研究周洪英湖北省农业科学院杏褪绿卷叶植原体宿主植物内生细菌的群落变化及与宿主抗/感病性的相关性研究韩 剑新疆农业大学竹虫肠道微生物群落结构及纤维素酶基因多样性分析王彦伟农业部沼气科学研究所木质纤维素水解残渣厌氧消化过程中微生物群落的组成与功能分析汤晓玉农业部沼气科学研究所海绵复杂共生体中放线菌新分类单元及新活性物质发现李 蕾上海交通大学贵州喀斯特洞穴放线菌多样性及生物活性菌株筛选房保柱中山大学放线菌中五角多酚类化合物的基因组挖掘刘力伟中国科学院微生物研究所疣孢菌CRISPR/Cas9基因组编辑技术的建立和评价谢 峰中国科学院微生物研究所3种黄连属濒危植物内生放线菌多样性及其抗菌消炎活性初探田守征云南中医学院中国丽赤壳属Calonectria种类及系统发育研究张云霞仲恺农业工程学院东北地区红菇真菌子实体及其地下菌根分子生态学研究冀瑞卿吉林农业大学山东苹果主产区红富士果面酵母菌多样性及对苹果炭疽病的生防潜能陈 汝山东省农业科学院中国毛霉属系统发育研究及DNA条形码筛选王亚宁中国科学院微生物研究所壳二胞属物种分类研究陈 倩中国科学院微生物研究所食药用菌真菌病害病原鉴定及系统分类研究孙敬祖中国科学院微生物研究所七种鞘翅目昆虫共生蛇口壳目真菌资源的分类与分子系统发育研究殷明亮广东省微生物研究所木兰科植物的丛枝菌根真菌多样性及协同进化特征杨安娜安徽师范大学我国海洋性冰川低温真菌多样性研究王曼曼河北大学云芝新颖苔色酸木糖苷衍生物的糖基化机制研究朱丽萍青岛农业大学中国芒果炭疽菌多样性及致病力变异分子机制的研究李其利广西壮族自治区农业科学院赤水河流域枯枝暗色丝孢菌Dematiaceous hyphomycetes多样性研究李小霞遵义师范学院鹅膏科系统发育框架构建及营养方式演化研究蔡 箐中国科学院昆明植物研究所西南地区葡萄座腔菌科四个重要属的分类和分子系统学发育研究刘建魁贵州省农业科学院严紧反应下乙酰化修饰对大肠埃希菌DnaA降解调控的研究张秋芬上海交通大学蓝细菌能量-还原力代谢重平衡新策略及其生理影响研究栾国栋中国科学院青岛生物能源与过程研究所海洋细菌胞外多糖 EPS273 抑制铜绿假单胞菌生物膜形成的分子机理研究吴仕梅中国科学院青岛生物能源与过程研究所1494生命科学第28卷里氏木霉sorbicillinoid类次级代谢产物生物合成机制的研究齐飞飞中国科学院青岛生物能源与过程研究所枯草芽胞杆菌产生羊毛硫细菌素subtilomycin促进其在植物体内定殖的机制邓 运华中农业大学沙雷氏菌属新种YD25中环脂肽类化合物与灵菌红素生物合成的共调控机制研究苏 春陕西师范大学青枯菌生物素合成途径中一种新型甲酯庚二酸单酰ACP酯酶的鉴定及其生物学功能研究贾 佳南京医科大学硫转移蛋白在嗜酸热硫氧化古菌Metallosphaera cuprina硫传递网络中的功能研究刘丽君西安医学院希瓦氏菌中D型β-内酰胺酶诱导表达及其耐药机制的研究音建华南昌大学酿酒酵母生物合成中长链二元酸的动态调控及其作用机制韩 丽郑州轻工业学院土壤杆菌胞外水溶性β-1,3-葡聚糖的生物合成机制研究程 瑞南京理工大学基于AFM力谱与SPR技术的启动子强度高效表征策略的研究张晓娟江南大学P450单加氧酶AveE催化阿维菌素呋喃环形成的机制研究马 莉中国科学院青岛生物能源与过程研究所丁香假单胞菌海藻糖合成与抗水分胁迫相关性研究余希岚湖北大学解脂耶氏酵母中蛋白激酶Snf1参与氮饥饿启动赤藓醇合成的调控机制刘晓燕淮阴师范学院转录因子RpoD调控运动发酵单胞菌乙醇耐受性的机制研究谭芙蓉农业部沼气科学研究所酮基合酶MarO催化maremycin中哌嗪二酮的酰胺键形成和成环释放机制黄婷婷上海交通大学多模块持续性内切纤维素酶CcCel9A的持续性驱动力研究张坤迪中国科学院青岛生物能源与过程研究所里氏木霉外切纤维素酶CBH I的水解机理研究和理性设计王业飞中国科学院青岛生物能源与过程研究所单酶催化多步连续反应中底物结合模式研究姚明东天津大学灰盖鬼伞胞外β葡聚糖苷酶BGL2不同变体产生机制和生理功能的研究刘中华南京师范大学蛹虫草隐花色素Cry-DASH功能及作用机制王 芬中国科学院微生物研究所酶分子对黄姜细胞壁降解及皂苷释放的影响机理研究魏 蜜湖北工程学院基因内置特征影响密码子偏好对重组蛋白表达调控的研究周 勉华东理工大学大肠杆菌脂多糖转运关键蛋白LptFG的功能研究向泉桔四川农业大学金黄色葡萄球菌七异戊二烯二磷酸合成酶SaHepPPs晶体结构和抗菌药物开发李 倩中国科学院天津工业生物技术研究所以产油微藻海洋微拟球藻为模式的二酰甘油酰基转移酶功能机制研究辛 一中国科学院青岛生物能源与过程研究所聚酮合酶pks7和pks11对海洋草酸青霉合成Oxalicumone A及其环境适应性的调控机制研究王 洁中国科学院南海海洋研究所大肠杆菌中galE转录暂停调控下游ρ依赖型转录终止的分子机制王 璕华中农业大学XsfP催化的膜内受控蛋白水解在黄单胞菌致病过程中的调控功能邓超颖中国科学院微生物研究所鞘脂合成调控因子Orm1在球孢白僵菌中的功能鉴定及其致病机理研究王娟娟济南大学糖多孢红霉菌中与红霉素合成相关调控因子间的干扰机制研究汪焰胜安徽大学特异腐质霉转录因子HiProA调控纤维素酶表达机制的研究徐欣欣中国农业科学院生物技术研究所利用代谢拨动开关调控大肠杆菌芳香型氨基酸的合成古鹏飞济南大学CRISPR/Cas9介导的基因组进化构建固态发酵耐热酵母及机理研究李鹏松清华大学代谢工程改造枯草芽孢杆菌合成N-乙酰神经氨酸关键问题的研究刘延峰江南大学米曲霉Sedolisin家族基因相关联的外源蛋白表达分泌机制的研究朱 琳江苏大学L-异亮氨酸合成途径基因在谷氨酸棒杆菌中的模块化协调表达和代谢调控机制研究尹良鸿浙江农林大学酿酒酵母合成灵芝酸类似物的研究肖 晗上海交通大学聚3-羟基丁酸-4-羟基丁酸酯的全新合成代谢通路的构建与优化尹 进清华大学二硫吡咯酮生物合成途径中N4甲基转移酶的表征及组合生物合成应用黄 胜湖北民族学院大肠杆菌中植物CPR:P450模块化自主共价连接系统的构建及其在咖啡酸合成中的应用李宜奎江苏省中国科学院植物研究所胆囊癌微环境菌群结构特征和功能学的初步研究吴文广上海交通大学国家自然科学基金委员会生命科学部2016年度资助项目第12期1495单细胞水平高产虾青素雨生红球藻高通量筛选新方法研究王喜先中国科学院青岛生物能源与过程研究所基于功能活性可视化追踪的芳烃降解微生物高通量筛选方法方 云广东省微生物研究所微生物漆酶数据挖掘及底物杂泛性分析张寅良安徽大学亚硝酸盐还原酶转录调控蛋白NsrR去阻遏机制研究令桢民兰州大学异丙甲草胺脱烷基酶基因的克隆及脱烷基酶特性和功能的研究陈 青枣庄学院面向人工定向重构纤维素菌群的共生分子机理研究杜 然清华大学迪茨氏菌转录调控蛋白AlkX对烷烃降解过程的全局调控研究梁洁良中山大学GDSL酯酶的定向进化与分子改造研究丁俊美云南师范大学整合宏组学方法研究番茄秸秆堆肥生境中的关键微生物及其功能张小梅青岛农业大学假单胞菌CNB-12分解代谢3-氯硝基苯的机理研究闵 军中国科学院烟台海岸带研究所假单胞菌LY1降解3-吲哚乙酸上游途径分子机理研究于 浩青岛农业大学一种二苯醚类污染物直接开环角度双加氧酶的分子机制研究蔡 舒江苏省农业科学院根际促生解淀粉芽孢杆菌SQR9组氨酸激酶KinA-E响应的根系分泌物信号鉴定刘云鹏中国农业科学院农业资源与农业区划研究所膜囊泡介导Geobacter sulfurreducens胞外电子传递过程及机制刘 星福建农林大学短短芽孢杆菌Spo0A蛋白介导的生物膜调控通路中阻遏基因的功能分析侯启会山东农业大学高盐环境下盐单胞菌(Halomonas)降解偶氮染料的机制研究郭 光南京工程学院纳米氧化铝促海洋枯草芽孢杆菌抗菌物质合成机理研究于秀霞山东大学红树林湿地生态系统中MCG古菌mcrA基因的时空变化规律和环境效应研究潘 杰深圳大学基于秀丽隐杆线虫模型的抗菌性海洋益生菌筛选及其机理研究李英秀山东大学滇池水华中控藻菌的杀藻相关基因及其功能研究杨彩云西南大学太湖蓝藻群体颗粒附生细菌的宏基因组学研究张军毅东南大学稻田藻-菌生物膜中胞外聚合物对水体营养水平变化的响应机制刘俊琢中国科学院南京土壤研究所高原藏族人特异性皮肤菌群与紫外辐射适应关系研究曾 博四川农业大学学龄前儿童“口腔菌群年龄”的遗传基础和过程机制滕 飞中国科学院青岛生物能源与过程研究所地衣共生系统中地衣细菌群落结构及其功能研究司红丽山东师范大学广东省凡口铅锌矿矿山酸性废水中古菌的多样性与功能研究陈林兴中山大学鸡肠道微生物代谢黄曲霉毒素B1的分子机制研究汪玲玲华南农业大学西藏热泉环境栖热菌类群多样性及其生理生态功能研究周恩民中山大学中华蟾蜍蝌蚪肠道菌群的结构及其影响因素研究宋晓威信阳师范学院转录激活因子PilR在σ54调控水稻白叶枯病菌毒性中的功能研究余 超中国农业科学院植物保护研究所TLR介导的ATP释放在副猪嗜血杆菌感染上调IL-1β中的作用研究于 江山东省农业科学院分枝杆菌aceE基因影响细胞壁合成代谢的机制研究陈素婷首都医科大学碳青霉烯耐药鲍曼不动杆菌新优势克隆ST208毒力因子鉴定及功能研究陈 燕浙江中医药大学沙门氏菌毒素效应蛋白SifA C-端结构域毒力功能机制的研究赵伟栋新乡医学院MexS调控铜绿假单胞菌III型分泌系统分子机制的研究靳永新南开大学基于炭疽芽孢杆菌S-层蛋白自组装的双抗高灵敏检测纳米材料的研究王旭颖武汉血液中心c-di-AMP信号通路在炭疽杆菌致病机制中的功能研究胡 葭中国科学院武汉病毒研究所生物被膜在沙门菌逃逸肠黏膜树突状细胞免疫监视中的作用和机制阴银燕扬州大学结核分枝杆菌广谱胁迫蛋白Rv1996介导的异烟肼耐药机制研究胡新玲中国科学院微生物研究所一种肺炎链球菌磷壁酸合成相关蛋白的功能鉴定吴凯峰遵义医学院NO调控鲍曼不动杆菌多重耐药的机制研究邓珊珊成都医学院禾谷镰刀菌FgPrp6调控剪接体激活的作用机制研究金巧军西北农林科技大学高渗胁迫下应激活性蛋白激酶afSakA对黄曲霉毒素合成的调控机制研究袁 军福建农林大学低铁环境下白色念珠菌核质转运受体Nmd5调控转录因子Sef1异常核输出的分子机制研究黄新华中国科学院上海巴斯德研究所Fasciclin-1结构蛋白在红色毛癣菌生长发育和侵袭宿主细胞中的作用机制研究占 萍南昌大学生命科学第28卷1496白念珠菌灰菌细胞的菌丝生长调控机制研究管国波中国科学院微生物研究所水稻条纹病毒Pc2蛋白在病毒侵染介体中的功能研究赵淑玲扬州大学南方水稻黑条矮缩病毒(SRBSDV) P7-1形成的管状结构进入细胞壁的机制及其在病毒运动网络中的功能谢 礼浙江省农业科学院反向长链非编码RNA调控NIA基因响应CMV侵染的机制研究赵建华中国科学院微生物研究所植物液泡脂质调控CMV病毒基因组复制的机制顾周杭浙江理工大学Dicer-2调控抗病毒Toll免疫通路的作用机制研究王赵玮武汉大学利用MDV载体递呈靶向ALV-J的CRISPR/Cas9系统预防MDV和ALV-J感染的研究李 凯中国农业科学院哈尔滨兽医研究所NS1蛋白与人核仁磷酸化蛋白1互作在A型流感病毒感染过程中的作用研究朱春玉辽宁大学乙型肝炎病毒HBx类泛素化Neddylation修饰的功能及生物学意义刘宁宁中国科学院微生物研究所Nrf2/ARE信号通路对RHDV感染中肝脏氧化应激的作用胡 波江苏省农业科学院汉滩病毒核衣壳蛋白核质转运信号的鉴定及其在病毒感染中功能的研究叶 伟中国人民解放军第四军医大学免疫缺陷病毒Vpx蛋白识别宿主CRL4 (DCAF1) E3泛素连接酶的作用机制研究魏 伟吉林大学新型EV71病毒抑制剂靶向病毒3D蛋白的作用机制及其抗病毒功能研究张 伟苏州大学乙型肝炎病毒C蛋白促进Src/PI3K/Akt通路介导的肝细胞周期进程及机制刘 伟三峡大学基于"HCV core-ZEB相互作用促进EMT"探讨HCV感染相关性肝癌的发生机制张利军重庆医科大学抗肿瘤化合物tyrphostin AG490抑制丙型肝炎病毒NS5B聚合酶活性的分子机理研究杨 娜中国科学院海洋研究所GADD45G蛋白通过组蛋白修饰调控HSV-1感染的分子机理研究陈晓庆广州医科大学新型H3N2人流感的进化及受体结合特性研究路希山中国科学院天津工业生物技术研究所用VLP表达HPIV3的HN和F蛋白及其介导的免疫应答对小鼠的保护作用研究张光媛重庆医科大学CCHFV囊膜蛋白Gc结构域III与病毒入侵相关位点研究张怀东中国科学院武汉病毒研究所乙型脑炎病毒RNA元件的宿主特异性研究刘思情中国科学院武汉病毒研究所HIV-1利用CypA躲避TRIM11加速脱衣壳作用的机制研究袁 婷中国科学院武汉病毒研究所ISlncRNA-23促进HIV-1复制的机制研究董银慧中国科学院生物物理研究所发热伴血小板减少综合征病毒对IL-10和IL-11抗炎信号的抑制机制宁云佳中国科学院武汉病毒研究所HCMV通过IE1调控神经干细胞中Hes1表达及节律的机制李小军中国科学院武汉病毒研究所柠檬酸铁的抗病毒作用与机制研究王洪斌中国科学院上海巴斯德研究所长链非编码RNA A VIRL在抗流感病毒感染中的作用机制研究陈玉海中国科学院微生物研究所噬菌体SWU1杀菌过程中ROS的作用及产生机制研究樊祥宇济南大学gp38蛋白决定大肠杆菌噬菌体Bp7宽宿主谱的机制张 灿青岛农业大学鹦鹉热嗜衣原体两个TMH家族蛋白结构与功能研究伍海英南华大学黑龙江立克次体表面蛋白与血管内皮细胞(宿主细胞)表面分子相互作用的研究齐 永中国人民解放军南京军区军事医学研究所2植物学NAC转录因子调控荔枝果柄离区细胞凋亡的机理研究李彩琴华南农业大学细胞壁组分与复合多层结构对小麦籽粒水分及品质调控机理的研究应瑞峰南京林业大学水稻极度矮化基因STD1的图位克隆与功能分析房静静中国农业科学院作物科学研究所复苏植物牛耳草DnaJ蛋白参与水分胁迫下叶绿体保护的作用机理刘 杰中国科学院植物研究所PRSL1通过蛋白磷酸酶PP1调控拟南芥植株形态的分子机理秦倩倩兰州大学拟南芥WUSCHEL互作因子WIC1调控茎端分生组织功能的机理研究周 超山东农业大学虎耳草科虎耳草属石荷叶组的分类学研究张卓欣华南农业大学国家自然科学基金委员会生命科学部2016年度资助项目第12期1497中国景天科山景天组的分类学研究孟世勇北京大学日本蛇根草和广州蛇根草及其近缘类群的系统分类学研究吴 磊中南林业科技大学万寿竹属的分类学和系统学研究朱鑫鑫信阳师范学院豆科甘草属分子系统发育与生物地理学研究段 磊中国科学院华南植物园苍山冷杉复合体的物种划分与气候响应模式研究邵毅贞河南农业大学泛喜马拉雅地区鼠尾草属的分类修订胡国雄贵州大学豆科长柄山蚂蝗属的分子系统学研究宋柱秋中国科学院华南植物园中国长篦藻属硅藻的分类修订及分子系统学研究刘 琪山西大学平叶多褶苔和变异多褶苔的物种界定以及洲际间断分布格局的形成原因师雪芹安徽师范大学绿藻门橘色藻目的系统分类学研究朱 欢中国科学院水生生物研究所北半球间断分布植物珊瑚菜的亲缘地理学和保护遗传学研究李密密江苏省中国科学院植物研究所东亚植物区系空间分化的分子机制——棣棠花和粉花绣线菊复合群的谱系地理学研究罗 冬中国科学院昆明植物研究所热带亚洲鞭苔属的系统分类学研究董珊珊华南农业大学溯祖方法在系统发育基因组学分析中的可靠性席祯翔四川大学基于ddRAD-seq技术的悬竹属时空演化格局研究张宪智西北农林科技大学银莲花属西南银莲花组和鹅掌草组的分子系统学研究张 煜湖南科技大学双盖蕨属(蹄盖蕨科)的系统发育和物种多样性分化研究卫 然中国科学院植物研究所小檗科(Berberidaceae)系统基因组学及叶绿体基因组进化研究孙延霞中国科学院武汉植物园鬼灯檠属的进化历史研究马祥光中国科学院昆明植物研究所浅苞橐吾、云南橐吾和大黄橐吾之间的自然杂交和基因渗入研究余姣君中国科学院昆明植物研究所芸薹属油菜类作物细胞质基因组单倍型的精细解析及其协同进化分析乔江伟中国农业科学院油料作物研究所荨麻科叶绿体系统发育基因组学研究吴增源中国科学院昆明植物研究所大豆脂肪酸脱氢酶FAD2家族酶活性差异的进化与功能研究赵 嫚浙江工业大学印度-西太平洋地区红树基因组渐渗与物种分化机制研究何子文中山大学青藏高原—蒙古高原—中亚地区砾玄参复合群的亲缘地理学研究王瑞红浙江理工大学小麦杀配子现象中存在的转录组变异及miRNAs在其中的调控作用研究白 琰哈尔滨师范大学水玉簪属(Burmannia L.)植物与丛枝菌根真菌协同进化研究赵中涛中国科学院华南植物园结合几何形态学方法探索泽泻科植物心皮的发育与进化黄岚杰湖北大学荠属种内叶形自然变异的研究杨 丽中国科学院植物研究所姜科唇瓣的发生、发育及其分子机理研究李秀梅广东省农业科学院农业生物基因研究中心同域分布老鹳草属植物花冠开口方向的分化机制王 慧华中农业大学Hlips在保护蓝藻光系统II免受氨损伤中的机制研究戴国政华中师范大学管藻黄素型LHCII参与的假根羽藻非光化学淬灭机制研究王文达中国科学院植物研究所D1蛋白周转过程的调控机制研究-以短命植物为例涂文凤中国科学院植物研究所糖基化调控水稻乙醇酸氧化酶与过氧化氢酶互作及过氧化氢信号发生的机理研究张智胜湖南农业大学水稻镉吸收和积累相关基因的发掘和功能鉴定杨 猛华中农业大学硫-TOR信号通路介导拟南芥生长的分子机理研究徐 萍中国科学院上海生命科学研究院木薯碱性/中性转化酶MeNINV1的酶活性调节机制研究姚 远中国热带农业科学院热带生物技术研究所拟南芥转录因子NAC103在逆境胁迫响应中的生物学功能及其调控机理孙 玲江苏大学基于ssRNA-seq的木薯抗旱lncRNA的挖掘及相关基因调控网络的研究丁泽红中国热带农业科学院热带生物技术研究所BHLH转录因子HBI1调控植物生长和免疫抗病动态平衡的分子机制研究樊 敏山东大学转录因子基因LbCPC参与二色补血草盐腺分化的功能研究袁 芳山东师范大学GSK3类蛋白激酶SGK1通过SOS2调节植物耐盐性的分子机制研究周华鹏四川大学生命科学第28卷1498拟南芥内质网膜蛋白ROOT HAIR DEFECTIVE 3 (RHD3)调控花青素代谢分子机理王 静北方民族大学大豆转录因子SNAC的无序序列区对耐盐相关基因表达调控的分子基础刘国宝深圳大学光周期下拟南芥CAT2蛋白降解的分子机制研究苏 彤山东师范大学拟南芥类受体蛋白激酶CRKN1在脱落酸信号转导中的功能研究梁 杉清华大学一氧化氮调控拟南芥体内硼含量稳态的分子机制研究夏金婵河南中医学院植物高温响应的表观记忆机制刘军钟中国科学院上海生命科学研究院拟南芥光信号转录因子FAR1与EDS1互作调控植物免疫的机理研究王晚晴北京联合大学非传统G蛋白及其激活蛋白的晶体结构与水稻应答盐胁迫机制的关系解析苗 锐福建农林大学拟南芥MYB102通过调控细胞壁扩展蛋白的表达增强植物耐旱的分子机理研究周 成安徽科技学院大豆高盐响应蛋白GmOSM的调控机理研究万 群江苏省农业科学院拟南芥新型液泡阴离子通道(VSAC1和VSAC2)介导细胞水势调控的分子机制研究张海纹北京市农林科学院野生番茄响应昆虫唾液中FAC诱导物的遗传基础研究申国境中国科学院昆明植物研究所膜结合转录因子NAC091调控植物内质网胁迫应答的分子机理研究杨正婷贵州师范大学拟南芥丝氨酸羧肽酶SCPL41基因在干旱胁迫中的作用及机制贾艳霞中国科学院昆明植物研究所小麦类钙调素调节植物耐盐性的功能研究周 硕河北省农林科学院遗传生理研究所拟南芥RopGEF7的互作蛋白eIF4E1参与生长素介导的植物发育的分子机制刘太波华南农业大学拟南芥CKRW1调节内源生长素水平稳态平衡的分子机理研究武 磊兰州大学拟南芥乙酰转移酶HLS1在BR与Auxin协同调控植物生长发育中的功能研究刘晓磊中国科学院上海生命科学研究院GA与BR共同调控拟南芥纤维素合成的分子机制研究王 昕沈阳大学独脚金内酯信号通路D53-like SMXLs下游转录因子的鉴定与功能分析王 冰中国科学院遗传与发育生物学研究所一氧化氮与细胞分裂素信号通路互作调节植物适应性生长的分子机制张燕香中国科学院遗传与发育生物学研究所转录因子GhKNOX1-1调控棉花叶片形态建成的分子机制研究肖光辉陕西师范大学拟南芥WRKY71转录因子调控叶片衰老的分子机制研究于延冲青岛农业大学RLF1亚硝基化介导生长素调控水稻侧根发育的分子机制孙爱珍中国科学院上海生命科学研究院油菜生长素合成相关基因BnaA.YUCCA6调控分枝角度的机理研究成洪涛中国农业科学院油料作物研究所类受体激酶SIT1调节水稻叶片衰老的分子机制王 耕河北师范大学拟南芥隐花色素CRY2在介导蓝光依赖的生物钟调控中的作用机理研究曹世江福建农林大学周质微丝在植物网格蛋白介导内吞中的功能研究范路生福建农林大学IDD5的O-GlcNAc糖基化修饰在赤霉素信号转导通路中的作用KihyeShin福建农林大学甲基茉莉酸响应的bHLH类转录因子在青蒿素生物合成中的调控机制研究沈 乾上海交通大学糖基转移酶UGT78H2的活性鉴定及在黑莓类黄酮代谢中的功能分析陈 清四川农业大学玉米糖基转移酶UFGT2调节黄酮合成与耐逆的功能研究李燕洁山东大学FtMYB2对荞麦类黄酮生物合成的代谢调控机制及抗逆功能研究李晓华华中农业大学金柑类黄酮糖基转移酶基因功能分析及其调控网络构建刘小刚西南大学何首乌中芪合酶、白藜芦醇羟化酶基因的功能研究生书晶广东第二师范学院拟南芥LMBD2基因突变回复mur3-3表型的分子机制丁安明中国农业科学院烟草研究所博落回根中苄基异喹啉生物碱合成、转运和积累的细胞类型特异性定位及分子机制研究郑亚杰湖南农业大学拟南芥核质反向信号参与表观遗传调控的分子机理沈 杰中国科学院植物研究所灵芝三萜酸下游合成路径关键CYP450s基因挖掘与功能分析陈方方中国科学院武汉植物园文冠果性别分化的内源激素与microRNA调控机制敖 妍北京林业大学一个CCCH锌指蛋白调控水稻雄性生殖发育中胼胝质代谢的研究方瑞秋华南农业大学转录因子TDF1对拟南芥分泌型绒毡层发育与功能的转录调控楼 悦上海师范大学。

收稿日期:2004-08-10;修订日期:2004-12-15基金项目:中国科学院知识创新工程重要方向项目(KZCX3-S W -332)、国家自然科学基金项目(90211003)资助。
作者简介:孙志高(1979-),男,山东烟台人,博士研究生,主要从事环境生态与生物地球化学研究。
E -mail:zhigaosun@yahoo .com.cn15N 示踪技术在湿地氮素生物地球化学过程研究中的应用进展孙志高1,2,刘景双1,于君宝1,王金达1(1.中国科学院东北地理与农业生态研究所,吉林长春13001;2.中国科学院研究生院,北京100039)摘要:稳定性同位素技术是现代生态学研究中的一门应用技术,它几乎在生态学研究的各个领域都有着广泛的应用。
其中15N 技术由于具有示踪和区分氮素物质的源与去向等优越性而在生态系统氮循环研究中发挥了极为重要的作用。
文章主要从湿地氮素的输入过程、转化过程以及归趋过程三方面综述了该技术在当前国内外湿地氮素生物地球化学过程研究中的应用进展,特别指出当前基于该技术的湿地氮素生物地球化学过程研究尚缺乏一定的系统性、深入性和广泛性。
最后,文章就该技术在湿地氮素生物地球化学过程研究中的应用前景进行了展望研究。
关 键 词:生物地球化学过程;同位素分馏;15N 示踪技术;湿地中图分类号:X142 文献标识码:A 文章编号:1000-0690(2005)06-0762-07 湿地氮素生物地球化学过程是指氮素在湿地环境中进行的各种迁移转化和能量交换过程。
它以生物过程、化学过程和物理过程为主线,主要研究湿地系统内(湿地中各种沉积物以及湿地内生物新陈代谢所进行的氮素物质交换和转化过程)和系统外(湿地与毗邻生态系统之间进行的氮素物质交换过程)两种过程[1]。
U rey 关于同位素物质热力学性质的深入研究和N ier 研制的比值质谱计为稳定同位素地球化学奠定了基础[2]。
McKin 2ney [3]等对N ier 型质谱计的改进最终使稳定性同位素成为一种行之有效的分析方法。

第53卷第3期 辽 宁 化 工 Vol.53,No. 3 2024年3月 Liaoning Chemical Industry March,2024收稿日期: 2023-02-21 Feammox工艺反应性能 及其功能微生物研究王静雅,王宇佳,胡雪松,傅梓铖(沈阳建筑大学,辽宁 沈阳 110168)摘 要: 采用CSTR反应器对厌氧铁氨氧化(Feammox)反应性能和功能微生物进行了研究。
通过对厌氧消化污泥驯化启动Feammox反应,反应器运行温度为32 ℃,进水pH为7.0±0.1。
培养130天后,NH4+-N去除率可达40.97%,Fe3+转化率可达31.86%。
通过分析NH4+-N和Fe3+的转化率,考察不同pH和温度条件对Feammox反应的影响。
结果表明,最佳反应pH为7.0、温度为30 ℃。
高通量测序表明,反应器运行各阶段污泥中优势菌门为变形菌门、酸杆菌门及浮霉菌门,核心菌群为Desuifoprunum、GP10和GP7。
关 键 词:厌氧铁氨氧化;工艺启动与运行;温度;pH;氮素转化中图分类号:X703 文献标识码: A 文章编号: 1004-0935(2024)03-0370-05Feammox是NH4+直接氧化为N2、NO2-或NO3-,Fe(III)作为末端电子受体的厌氧铵氧化与微生物异相还原Fe(III)相结合的反应过程。
近年来已在水田、高原土壤、河岸湿地等富含氨氮和铁的厌氧环境中发现[1-3]。
这一现象的发现,为自然界氮元素与铁元素的转化提供了新的认识,被研究者广泛关注。
几种可能的Feammox反应式如表1所示。
表1 研究者推测的 Feammox 反应方程式研究者 预测方程式CLÉMENT[1]等 NH4++6FeOOH+10H+→NO2-+6Fe2++10H2OΔG= -30.9 kJ·mol-1YANG[4]等 3Fe(OH)3+5H++NH4+→3 Fe2++9H2O+0.5N2ΔG= -245 kJ·mol-16Fe(OH)3+10H++NH4+→6Fe2++16H2O+NO2-ΔG= -164 kJ·mol-18Fe(OH)3+14H++NH4+→8Fe2++21H2O+NO3-ΔG= -207 kJ·mol-1pH是影响Feammox的重要因素之一。


202丨年第49卷第3期 V〇1.49. No. 3,2021地球与环境EARTH AND ENVIRONMENT325磷酸盐氧同位素示踪环境中磷的来源与转化:原理、方法与应用杨小红,张瑞雪1,王敬富2’3〃,贺康康U2,陈敬安2’3,刘勇2’3(1.贵州大学资源与环境丁.程学院,贵阳550025:2.中国科学院地球化学研究所环境地球化学国家重点实验室,贵阳550081 ;3.中国科学院大学,北京100049)摘要:磷是地表环境中重要的生命元素。
由于研究手段的限制,目前有关磷生物地球化学循环过程及机制的认识仍然匮 乏。
作为一种新兴示踪剂,磷酸盐氧同位素(s l80p)成为当前研究磷生物地球化学循环的潜在有效工具。
本文综述了 &80p 示踪环境中磷来源与循环的基础原理,不同环境介质中s l80p的分离纯化及测试方法的研究进展,梳理了近年来Sl80p在环境 科学领域的应用,重点是土壤、沉积物和水生生态系统(河流、湖泊和海洋)。
在此基础上,提出了以8O p在未来工作上的展望:进一步扩展V8O p分析测试技术(如低磷、高溶解有机质的样品),在藻类、微生物体系中分馏效应的探索。
关键词:磷酸盐氧同位素(Sl8Op);来源示踪;生物地球化学循环;土壤和沉积物;水生生态系统中图分类号:X142文献标识码:A文章编号:1672-9250(2021)03-0325-11d o i:10. 14050/j. cnki. 1672-9250.2021.49.066磷被认为是所有已知生命形式的必要营养素,参与着生物的代谢过程,是核酸(〇〜六、1^~六)、三磷 酸腺苷(ATP)和磷脂等生物大分子结构和功能的重 要组成部分:1]。
在自然界,磷主要以正磷酸盐的形式广泛分布在土壤、岩石、海洋、湖泊中。
随着全球 人口的增加和工业的快速发展,人类对全球磷循环产生了巨大影响,使磷在地表环境中的流动大大增强[2]。
人为活动会释放出大量的磷,比如磷矿开采、化肥施用、牲畜养殖、粮食生产、人类消费和排泄物等[3_4]。

甲烷(CH4)和氨(NH3)是两种重要的生物体内催化剂,它们可以分别被甲烷氧化微生物和氨氧化微生物利用。
甲烷氧化微生物可以将甲烷氧化为甲醛(HCHO),而氨氧化微生物则可以将氨氧化为亚硝酸盐。
这两种微生物在生态系统中发挥着重要的作用,对于环境的生物地球化学过程和氮素循环具有重要影响。
1. 甲烷氧化微生物甲烷氧化微生物是一类嗜甲烷的微生物,它们可以在缺氧或微缺氧的环境中利用甲烷进行生长和能量获取。
甲烷氧化微生物广泛存在于土壤、水体和湿地等环境中,对减缓大气中甲烷含量具有重要意义。
甲烷氧化微生物的代表物种是甲烷单胞菌(Methylococcus),它们通过甲烷单加氧酶(MMO)和甲醛脱氢酶(ALDH)两个酶催化酶反应,将甲烷氧化为甲醛,再将甲醛氧化为二氧化碳和水。
2. 氨氧化微生物氨氧化微生物是一类能利用氨为唯一氮源的微生物,它们可以将氨氧化为亚硝酸盐,这是土壤中氮素循环的重要环节。
氨氧化微生物广泛存在于土壤和水体中,对于维持生态系统中的氮素平衡具有重要意义。
氨氧化微生物的代表物种是亚硝基细菌(Nitrosomonas),它们通过氨氧化酶(AMO)这一酶催化氨的氧化反应,将氨氧化为亚硝酸盐。
3. 甲烷氧化和氨氧化微生物的耦合功能除了各自单独发挥作用外,甲烷氧化和氨氧化微生物还可以发挥耦合功能,对于生态系统中的生物地球化学过程和氮素循环产生重要影响。
在一些沼气发酵产氨系统中,甲烷氧化和氨氧化微生物能够共同作用,将甲烷和氨转化为有机物和硝酸盐,这对于提高沼气发酵产氨系统的氮素利用效率具有重要意义。
甲烷氧化和氨氧化微生物的耦合功能还可以用于生物修复领域,由于甲烷和氨在环境中广泛存在,因此利用这两类微生物进行生物修复的研究已经引起了人们的重视。
甲烷氧化和氨氧化微生物及其耦合功能对于生态系统中的生物地球化学过程和氮素循环具有重要意义,不仅在减缓大气中甲烷含量、维持氮素平衡方面发挥作用,还可以应用于生物修复领域,具有重要的应用价值和研究前景。

第 35 卷 第 1 期环 境 科 学 研 究Vol.35,No.1 2022 年 1 月Research of Environmental Sciences Jan.,2022碳氮氧稳定同位素技术在水生态环境中的应用杨 蓉1,2,李 垒2*1. 北京市南水北调水质监测中心, 北京 1000972. 北京市水科学技术研究院, 北京 100048摘要:稳定同位素技术是研究环境和生态系统中元素循环途径的重要方法. 稳定同位素的丰度变化反映了自然界和生物体内混合、分馏双重作用的结果,因此可作为指标计算混合物的来源贡献,或研究造成分馏的化学反应和生物代谢路径. 从20世纪中期确立稳定同位素的基础原理,70年间该技术在地球化学、环境科学、生态学、微生物学、食品科学等领域获得了大量有价值的成果. 其中,水体作为自然环境的重要组成和人类社会的重要资源,已有诸多研究涉及稳定同位素在水环境中污染物溯源、水生态系统元素迁移转化、水生生物营养来源和营养关系等方面的应用. 通过梳理常见的碳、氮、氧稳定同位素在水环境和水生态领域的研究进展,发现污染物和食物来源分析已不局限于定性识别,基于数学模型的混合物组分定量评估方法正得到越来越多的应用;同时,为开展水体脱氮强度和通量估算、水生生物营养级计算及食物网分析,精确测量18O、15N和13C的富集程度以及通过试验和调研获取运算所需的基础参数都是关键步骤. 虽然在实际应用中存在待完善之处,但稳定同位素技术的前沿研究仍昭示了其整体化、精细化的发展方向. 未来与计算科学的方法学进步相结合,将为水科学研究提供更有力的技术支撑.关键词:稳定同位素;分馏;多元混合模型;硝酸盐溯源;食性分析;营养级中图分类号:X52文章编号:1001-6929(2022)01-0191-11文献标志码:A DOI:10.13198/j.issn.1001-6929.2021.07.03Applications of Stable Carbon, Nitrogen, and Oxygen Isotope Techniques in Aquatic Environment and EcologyYANG Rong1,2,LI Lei2*1. Beijing Water Quality Monitoring Center for South-to-North Water Diversion, Beijing 100097, China2. Beijing Water Science and Technology Institute, Beijing 100048, ChinaAbstract:Stable isotope technique is an important method to study element cycle in the environment and ecosystems. The change in the abundance of stable isotopes reflects the results of mixing and fractionation in nature and organisms. Therefore, it can be used as an indicator to calculate the contribution of mixture sources, or study the chemical reactions and biological pathways that cause fractionation. Since the establishment of the basic principles of stable isotopes in the middle of the last century, this technology has obtained many valuable achievements in geochemistry, environmental science, ecology, microbiology, food science and other fields in the past 70 years. Among them, there have been many studies on the application of stable isotopes to evaluate the pollutant sources, element migration and transformation, food sources and nutritional relations in the aquatic environment that is an essential component of nature and a significant resource of human society. By summarizing the research progress of common carbon, nitrogen and oxygen stable isotopes in the field of water environment and ecology, it is found that the analysis of pollutants and food sources is not limited to qualitative identification. In fact, quantitative assessment methods of mixture components based on mathematical models have been increasingly used. Meanwhile, to evaluate the intensity and flux of denitrification, or perform trophic level calculation and food-web analysis, it is important to measure the enrichment degree of 18O, 15N and 13C accurately, and obtain appropriate basic parameters for model through experiments and survey. Although some aspects still need to be optimized in practical applications, the research frontier of stable isotope technology shows its integrated and refined development direction. Combined with methodological advances in computational science, stable isotopes will provide more powerful technical support for water science research.收稿日期:2021-04-24 修订日期:2021-06-19作者简介:杨蓉(1987-),女,山西大同人,高级工程师,博士,主要从事水环境监测及评价研究,ygrg@.*责任作者,李垒(1980-),男,辽宁朝阳人,教授级高级工程师,博士,硕导,主要从事水生态环境评估和污染生态修复研究,ll@基金项目:国家自然科学基金项目(No.41301540)Supported by National Natural Science Foundation of China (No.41301540)Keywords:stable isotope;fractionation;multivariate mixed model;nitrate source identification;diet analysis;trophic level稳定同位素指核结构稳定、不发生或不易发生放射性衰变以及半衰期极长的一系列核素. 1912年,英国物理学家John Thomson发现天然氖由20和22两种不同质量数的原子组成,首次证明了常态元素稳定同位素的存在. 该技术的发展始于20世纪中期的稳定同位素地球化学领域,自然界常见的氢、碳、氮、氧、硫等元素都有超过一个的稳定同位素,研究者测定各类天然物质中元素的同位素组成,分析变化规律并推测原因所在.近年来,稳定同位素技术在地球化学、环境科学、生态学、微生物学、食品科学等领域得到了广泛应用[1-3]. 应用重点并非探究同位素丰度的分布特征,而是将其作为观测手段研究过程、开展溯源. 物理、化学和生物过程都可造成稳定同位素的丰度差异,反之稳定同位素化学性质相同,其变化不影响过程,因此可作为示踪剂为回溯环境、生物等多种因素对元素的共同作用提供科学有效的依据.水是生物不可或缺的基础物质,水生态环境不仅是环境科学和生态学的研究对象,其污染状况、物种分布等信息也是水资源管理的重要支撑. 随着色谱质谱联用和傅里叶变换核磁共振技术的诞生,同位素的检测手段愈加丰富,研究成本大幅降低,已有大量研究涉及稳定同位素在水体污染物溯源、水生生物食物网研究等不同方向的应用. 聚焦研究中最常见的碳、氮、氧稳定同位素,介绍了分馏和混合的基础理论及模型. 水质分析方面,以氮素和硝酸盐为例探讨了氮氧稳定同位素以及以IsoSource和SIAR为代表的混合模型在污染物迁移转化和溯源中的应用;水生态方面,基于碳氧稳定同位素特征、浓缩系数和基线生物选择综述了该技术在营养级、生态位和食性分析中的应用.1 稳定同位素分馏和混合的基础理论1.1改变同位素丰度变化的驱动力−分馏作用1.1.1分馏作用的定义同位素中子数的差异使其质量和原子间结合力不同,也导致了地球化学及生物化学过程中扩散和反应速率的差异.最终表现为同位素分馏(isotopic fractionation)效应,即一种元素的同位素以不同比例在不同物相中分配. 通常而言,轻同位素优先参与化学反应,造成动力学非平衡分馏[4]. 而当反应已达到化学平衡,不同化合物对重同位素的竞争导致了热力学平衡分馏.分馏效应形成了具有不同同位素丰度的物质库. 2H、13C、15N和18O等重同位素在环境中的绝对丰度低,一般接近或低于1%. 为了放大细微变化,便于比较,一般用样品和国际公认标准物质同位素比值的千分差描述稳定同位素含量,记作δ. δ值越大(越正)表示重同位素越多,越小(越负)表示轻同位素越多[5].1.1.2自然分馏自然界的物理过程和化学反应均会造成同位素分馏. 以氮素为例,挥发、扩散、水解、固氮、硝化、反硝化等反应体系具有不同的同位素分馏特征. 氨化作用造成的分馏仅为±1‰左右. 但硝化作用会在反应物里富集15N,浓缩系数在−29‰~−12‰之间;氨挥发也会导致NH4+中15N含量上升,氮的轻同位素更易于以NH3的形式挥发[6-7]. 李荣富等[8]总结了15N在氮循环过程各环节中的分馏系数,并指出并不是所有环节的同位素分馏都得到了清晰阐述,如硝酸盐异化还原、厌氧氨氧化的分馏效应就暂未明确.1.1.3营养分馏生物的新陈代谢会导致同位素分馏. 呼吸和排泄优先排出营养元素的轻同位素,造成食物中的重同位素在捕食者体内富集. 研究[9-10]表明,13C富集系数通常为0~1‰,而15N的浓缩系数通常为3‰~4‰,并随着食物链进一步放大. 因此,与食物组成相近的δ13C 常被用于追溯水体中捕食者的食物组成和贡献,富集程度更高的δ15N则用于开展水生态系统的营养水平研究[11-14].1.2计算源贡献的理论及方法−多源混合模型分馏和混合是稳定同位素研究中的两个重要概念. 分馏作用导致物质库之间的同位素丰度差异,而多个物质库的相互混合进一步造成环境中稳定同位素的复杂分布. 混合模型用数学方法推断混合物中各物质库的贡献率,被广泛应用于污染物定量溯源和捕食者食性分析.作为最基础的混合模型,质量平衡模型(mass-balance model)可用于n种同位素和n+1个来源的溯源计算. 最常用的是2种同位素和3个来源的定量评估,可以避免单一同位素的区分度不够(如NO3−中的15N). 但也有研究[15]指出质量平衡模型未考虑不确定性的影响,在混合物具有多种来源时不能使用. 为解决多来源的问题,2003年,Phillips等[16]通过对质量平衡方程组的反复迭代提出了IsoSource模型. IsoSource模型适用于n种同位素和>n+1个源,获得192环 境 科 学 研 究第 35 卷的不是点估计,而是分布. 2008年,Parnell等开发的SIAR(stable isotope analysis in R)模型引入了不确定性和同位素分馏的考虑,其基于狄利克雷分布和贝叶斯算法,结合似然函数和逻辑先验分布计算贡献率.吴文欢等[17]对质量平衡模型、IsoSource模型和SIAR 模型的适用情境和使用技巧进行了详细论述.除质量平衡模型、IsoSource模型和SIAR模型外,IsoError、IsoEconc、IsotopeR、MixSIR等模型也在污染物溯源中有所应用,Hopkins[18]等和冯建祥等[19]均比较了它们的原理、特征和优缺点. 但在实际使用中为避免误用,还需仔细评估背景资料是否充分,科学问题、试验设计和模型选择是否合理[20].2 氮氧双同位素在水体氮素迁移转化及溯源中的应用2.1反硝化脱氮氮循环是生物地球化学循环中的重要环节,生态系统中氮的输入、转化、利用和消除都是被长期探讨的科学课题. 氮氧双同位素用于脱氮过程研究,提供了用稳定同位素探讨元素迁移转化的范例.厌氧条件下反硝化菌有偏好地利用轻同位素转化成N2和N2O,造成15N和18O富集在剩余的NO3−中,二者的同位素分馏系数比值约为2,即δ15N升高1‰,δ18O相应上升0.5‰[6]. Xue等[7]认为,实际情况下15N 和18O以1.3∶1~2.1∶1的比例富集都可以推论反硝化的发生. 水体脱氮过程中氮氧同位素分馏系数如表1所示. Panno等[26]、Houlton等[28]分别试算了森林和密西西比河中因反硝化损失的氮量,为研究氮素迁移转化提供了新方法.2.2硝酸盐溯源我国许多地方的地表水和地下水正在面临硝酸盐污染[29-30]. 六成分图、派珀图等是硝酸盐污染分析的经典方法[31],但随着同位素方法的建立和推广,氮氧双同位素被广泛用于氮源定性识别. 1998年Kendall等[6]根据大量试验结果提出了利用氮氧双同位素示踪不同来源硝酸盐的经验方法,利用δ15N的差异区分来自肥料和降水中NH4+硝化、土壤氮素、有机肥与污水的硝酸盐,再通过δ18O的差异区分来自硝酸盐肥料和大气降水的硝酸盐. 该方法得到了后续研究的证实[7,32-33].氮氧同位素溯源研究通常需要结合水化学和其他同位素数据一起进行[34]. 例如,Min等[35]采用δ15N 联合NO3−浓度解释了生活污水和化肥对韩国洛东江流域地下水硝酸盐的贡献;Nyilitya等[36]利用NO3−、Cl−、硼(boron, B)浓度和δ11B探讨了肯尼亚基苏木城和Kano平原地下水硝酸盐来源及转化;Wang等[37]比较了Cl−浓度与NO3−浓度、NO3−/Cl−的相关性,以讨论云南程海氮素的来源和反硝化的发生;Wang等[38]在赣江流域分析了δ15N与NO3−浓度、NO3−/Cl−的关系. 此外,研究地点的背景信息(如土地使用情况)[31,38]、排放负荷和入水负荷[39]等也常被用作综合分析.在氮源定性识别的基础上,量化污染源贡献率的定量评估方法也逐渐受到青睐. 研究者将一系列数学模型用于地表水和地下水的硝酸盐来源探讨,得到氮源的相对贡献. 近年来,IsoSource和SIAR模型在我国各地得到了越来越多的应用(见表2).表 1 水体脱氮过程中氮氧同位素分馏系数对比Table 1 Fractionation factors of nitrogen and oxygen stableisotope in aquatic denitrification采样区域水体类型氮氧同位素分馏系数比值数据来源溪流岸边带地下水 1.5:1文献[21]河流岸边带地下水 1.3:1文献[22]城市含水层地下水 1.9:1文献[23]湖泊深水层地表水 1.8:1文献[24]脱氮菌培养系统模拟淡水/海水1:1(兼性厌氧反硝化菌)、1.6:1(光合异养菌)文献[25]森林地区土壤溶液 1.5:1文献[26]森林地区土壤溶液 1.7:1 ~ 3.6:1文献[27]表 2 稳定同位素在硝酸盐定量溯源中的应用Table 2 Application of stable isotopes in nitrate source identification研究区域水体类型模型名称主要结论数据来源江苏滆湖地表水质量平衡模型临近城市的河流工业废水对硝态氮贡献率为76%~82%;远离工业区的样点硝酸盐来源主要是农业面源和生活污水,二者之和在60%~80%之间文献[39]德国瓦尔诺河地表水质量平衡模型灌溉用水、地下水和大气降水对瓦尔诺河水中硝态氮的贡献率分别为86%、11%和3%文献[40]山西汾河地表水IsoSource模型丰水期临汾段和M5采样点污染源主要是粪便和生活污水,占比为40%~72%,其余采样点化肥贡献率高,为45%~62%;枯水期粪便和污水是所有采样点硝态氮的主要来源,占比为40%~73%文献[41]第 1 期杨 蓉等:碳氮氧稳定同位素技术在水生态环境中的应用1933 碳氮双同位素在水生生物营养谱构建和食性研究中的应用3.1营养级估算稳定同位素在营养生态学的应用始于20世纪90年代中期. 以营养分馏为基础,聚焦水生生物体内的15N含量可反映生态系统中不同物种的营养水平. Zanden等[59]发现传统食物分析和稳定同位素法得到的342种鱼类的营养级无显著性差异,证明了新方法的可靠性. 稳定同位素计算结果用连续数值标示生物在食物网中所处位置,与传统方法获得的正整数相比更能反映其在生态系统中的真实状况[60].捕食者和食物之间较确定的δ15N差值是稳定同位素用于营养级关系计算的基础,根据实际需要选用有针对性的浓缩系数和基线生物是开展研究的前提. Minagawa等[10]的试验表明15N浓缩系数的平均值±标准差为3.4‰±1.1‰,Post[61]和Mccutchan等[62]基于文献调研的结果分别为3.4‰±0.98‰和2.2‰±0.18‰,相差不大. 多种原因造成了浓缩系数的差异,如Mccutchan等[62]发现,针对不同的食物结构和捕食者部位,15N呈现出不同的富集模式;Zanden等[63]按照生物分类(鱼类和无脊椎动物)、栖息地(海水和淡水)、调查方式(实验室和野外)和食物类型(肉食和 续表 2研究区域水体类型模型名称主要结论数据来源内蒙古乌梁素海地表水IsoSource模型化肥和土壤氮相关的农业活动对春季湖水硝酸盐的贡献率为43.7%;生产生活污水是其他3个季节硝酸盐氮的主要来源,贡献率分别为51.3%、38.8%和40.2%文献[42]山东王河地下水库地下水IsoSource模型库区中硝酸盐主要来源于化肥和生活污水,二者的平均贡献率分别为61.44%和33%文献[43]广西寨底地下河地下水IsoSource模型硝酸盐来源以化肥、动物粪便和生活污水为主,且距离居民区较远时化肥贡献比例较高,距居民区较近时动物粪便和生活污水贡献比例更高文献[44]重庆中梁山槽谷区地下水IsoSource模型雨季土壤氮、粪便及污水、降水和化肥中的氨氮对硝酸盐的平均贡献率分别为26%、21%和45%,旱季分别为20%、47%和19%文献[45]重庆青木关地下河地下水IsoSource模型不同采样点的硝酸盐污染特征不同,临近居民区和养殖区的采样点主要污染来源分别为土壤有机氮(48.3%)和粪便污水(59.6%)文献[46]山东桓台地下水IsoSource模型和SIAR模型硝酸盐主要来源于化肥(45%~46.9%)和污水(40.7%~51.7%),畜禽养殖也有一定贡献(2.6%~10.7%)文献[47]河北洋河地表水SIAR模型硝酸盐来源贡献表现为粪便生活污水(45.37%)>土壤氮(41.39%)>降水和化肥中的氨氮(13.24%)文献[48]陕西浐河灞河地表水SIAR模型硝酸盐来源贡献表现为污水及粪便>土壤有机氮>化肥>降雨,其中污水及粪便在浐河灞河的贡献率分别为30%和36%文献[49]安徽滁州地表水SIAR模型无论雨季和旱季,粪便和生活污水(28%~36%)及土壤氮(24%~27%)都是硝酸盐的主要来源文献[50]浙江长兴地表水SIAR模型12月硝酸盐来源以粪便和生活污水为主,占比为52%;5月以化肥为主,占比为37%文献[51]山东潘庄引黄灌区地下水SIAR模型粪便和生活污水的贡献率最高,为35.1%~80.5%,其次是化肥,占比为4.9%~31.7%文献[52]河北洋河地下水SIAR模型硝酸盐氮来源分别为土壤氮(44.36%)、粪便及污水(43.35%)、无机化肥及工业废水(9.24%)文献[53]贵州会仙湿地地下水SIAR模型动物粪便及生活污水、化肥、土壤氮是主要的硝态氮来源,贡献率差别不大,平均值分别为39.1%、32.2%和28.5%文献[54]比利时弗兰德斯地表水SIAR模型粪便和生活污水是主要的硝态氮来源,冬季贡献率为9%~85%,夏季贡献率为8%~59%文献[55]美国匹兹堡地表水SIAR模型94%的硝酸盐来自污水;暴雨情况下34%的硝酸盐来自大气降水文献[56]希腊阿索波斯盆地地表水和地下水SIAR模型离城市和工业区越远,城市和工业废水对硝酸盐贡献越大,反之化肥和粪便贡献越大文献[57]韩国京畿道地下水SIAR模型农业用地的硝酸盐主要来源于化肥(贡献率为35%~71%),其次是有机肥料(包括堆肥,贡献率为39%~49%)文献[58]194环 境 科 学 研 究第 35 卷植食)比较了15N的浓缩系数,发现后二者有显著性差异(P<0.05),但整体在3‰左右;万祎等[64]和蔡德陵等[65]分别通过渤海湾水生生物网调查和鳀鱼养殖试验得出了3.8‰和2.5‰的15N浓缩系数并得到应用[64,66],但很多研究还是采用3.4‰[67-69].通常情况下浮游植物的数量变动剧烈,不适合反映水域时间和空间的平均信息,因此基线生物多选用研究水域中常年存在、食性单一的浮游动物或底栖动物[70],如魏虎进[66]等、李红燕[71]均以水体浮游动物优势种中华哲水蚤、太平洋纺锤水蚤、针刺拟哲水蚤作为基线生物,以其δ15N值为基线值. 也有研究[61]认为,浮游动物易受外界干扰、季节波动明显,应选择个体较大、生活周期长的螺类和双壳类,如已有实际应用的福寿螺[68]、铜锈环棱螺[72]、珠蚌[73]、栉孔扇贝[69]、翡翠贻贝[70]等. 在国内,稳定同位素技术曾被用于三峡库区、太湖等淡水水域以及象山港、胶州湾、流沙湾等海域的连续营养谱构建,所调查的生物、使用的基线生物和浓缩系数等如表3所示.表 3 稳定同位素在国内水域营养谱构建中的应用Table 3 Application of stable isotopes in domestic trophic position calculation 采样区域水域类型受试生物基线生物浓缩系数/‰数据来源小江库湾淡水POM、固着藻类、软体动物、水生植物、鱼类不详 3.4文献[67]东太湖淡水苦草、浮游生物、底栖动物、鱼类铜锈环棱螺、河蚬 3.4文献[72]象山港海水浮游生物、SOM、POM、底栖生物、鱼类浮游动物 2.5文献[66]东营、烟台潮间带海水海草、底栖动物、鱼类青蛤、异白樱蛤 3.4文献[74]胶州湾海水浮游生物、底栖动物、鱼类中型浮游动物 3.4文献[75]大连近岸海水底栖动物、鱼类栉孔扇贝 3.4文献[69]枸杞岛海水浮游生物、SOM、POM、底栖生物、鱼类小型浮游桡足类 3.4文献[76]流沙湾海水浮游生物、底栖大型海藻、贝类、虾蟹、头足类、鱼类翡翠贻贝 2.5文献[70]中国澳门海水软体动物、虾蟹、鱼类、鸟类颗粒有机物 3.4文献[77]上海市崇明区海水底栖动物绿螂 2.9文献[78]3.2生态位研究利用δ13C-δ15N散点图量化营养结构和生态位的方法为食物网研究提供了新思路. 在用δ13C-δ15N二维坐标关联同位素含量和营养功能群的基础上[79],提出δ15N差值(δ15N range, NR),δ13C差值(δ13C range, CR)、总面积(total area, TA)、平均离心距离(mean distance to centroid, CD)、平均最邻近距离(mean nearest neighbor distance, NND)和平均最邻近距离标准差(standard deviation of nearest neighbor distance, SDNND)6个指标,分别表示营养层次、食源多样性、占据生态位或食物网中营养多样性的总量、营养多样性平均水平、群落的整体密度和营养生态位分布的均匀程度[80]. 该方法量化了食物网的营养结构多样性程度和冗余度,有利于评价生态系统中每个物种的功能及生态位变化.张文博等[81]和谢斌等[82]使用δ13C-δ15N散点图研究了海洋渔场中小型消费者的碳氮稳定同位素比值,分别分析了华南海陵湾、陵水湾两个水域和连云港海州湾不同季节的水生生物NR、CR、TA、CD、NND、SDNND的时空变化. 盖珊珊等[83]和俞雅文等[84]将该方法用于不同生物的生态位研究,分别探讨了两种鱼类和两种蟹类生态位的宽度和重叠程度. 其中,盖珊珊等[83]除NR、CR和TA外,还计算了标准椭圆面积(standard ellipse area, SEA)和营养生态位重叠面积(overlap area, OA). SEA被认为比TA更好,且更不易受样本数量干扰[85]. OA量化了生态位的重叠程度,能更好地评价不同物种利用食物资源的竞争强弱.3.3食物源分析胃含物分析是开展食性研究的经典方法,通过直接解剖动物胃肠道分析残留食物的组成,以此了解其食物来源. 实际应用中发现其具有一系列局限性,包括样本量小时偶然性强、不适于难以解剖的小型动物、只能反映较短时间内的摄食状况、食物残渣偏向难消化的食物类型等[86]. 与之相比,生物体内某些组织的稳定同位素具有较长的周转时间,可用来研究长生命周期内消费者对食物的代谢和吸收[87-88].与定性判断氮源类似,可通过对比捕食者和不同来源食物的同位素值分析食物组成. 例如,当几种潜在食源的δ13C值差异显著,消费者的δ13C落在颗粒有机物和固着藻类之间,可认为这二者是主要食物[67]. 张波等[89]根据生物的δ13C值由栖息水层加深逐渐增大,提出了崂山湾鳅虎鱼不同生命阶段摄食不同深度水生生物的变化规律;崔莹等[90]结合中华绒第 1 期杨 蓉等:碳氮氧稳定同位素技术在水生态环境中的应用195螯蟹各发育阶段特点和碳氮同位素比值分析其洄游期食物组成,强调了结合稳定同位素与生活习性开展分析的必要性. 定量分析方面,混合模型同样被用于食物贡献率计算,稳定同位素在水生生物食性研究中的应用如表4所示. 此时要注意通过胃含物、文献查阅等传统方法确定食源范围,避免取回的样品缺失重要的食物来源;另外,尤其针对15N等营养分馏较明显的同位素,需要选取合适的分馏系数[97].表 4 稳定同位素在水生生物食性研究中的应用Table 4 Application of stable isotopes in energy sources analysis同位素模型名称主要内容数据来源C、N质量平衡模型计算了5种捕食者2~3个食物来源的贡献,C和N的营养分馏系数分别取1.3‰和3.3‰文献[91]C、N质量平衡模型计算了陆源植物碎屑、丝状藻类和水生昆虫对两种淡水螯虾的贡献. C和N的营养分馏系数分别取0.8‰和3.4‰文献[92]C IsoSource模型分析了沉积有机物(SOM)、悬浮颗粒有机物(POM)和浮游植物3种食源对海州湾牧场28种消费者的贡献文献[82]C IsoSource模型分析了石莼、浒苔、海带、POM、SOM和浮游植物6种食源对象山港牧场34种消费者的贡献文献[66]C IsoSource模型分析了C3植物、C4植物和微型藻类3种食源对3种鱼类和1种虾类的贡献文献[93]C IsoSource模型分析了底层鱼类、中下层鱼类、中上层鱼类、虾类、蟹类和头足类共6类28种食源对江豚的食物贡献比例文献[94]C、N SIAR模型分析了8类19种潜在食源在精养和共作两种养殖模式下对克氏原螯虾的贡献.C和N采用甲壳类的分馏系数,分别为1.3‰和3.9‰文献[68]C、N SIAR模型分析了POM、浮游动物、鱼类和甲壳类4种食源对闽江口凤鲚的贡献.C和N的营养富集因子分别为0.4‰和3.4‰文献[95]C、N、S SIMMR模型26种食源被分成无脊椎动物4组和鱼类3组,考察了其对半干涸河口3种主要鱼类的食物贡献. C、N和S的营养分馏系数分别为1.0‰、3.3‰和0文献[96]4 其他特定化合物稳定同位素技术利用氮氧同位素进行硝酸盐溯源是针对化合物(或离子)中某元素开展的稳定同位素分析(compound-specific isotope analysis, CSIA)应用之一. 在环境科学领域,CSIA也被用于卤代烃、多环芳烃等有机污染物的溯源[98]. 而在微生物系统结构和生物化学过程解析方面,Ohkochi等[3]分离纯化了日本Kaiike湖中的光合色素并测定其碳氮稳定同位素组成,解释了与色素相关的自养微生物的生活深度、同化途径和生物功能. Isaji等[99]测定了意大利某盐场中营养盐、叶绿素的δ15N变化,以此探讨底栖微生物中氮素的转化途径及铵态氮在初级生产中的循环利用.上文所述碳氮同位素分析食源和营养级的研究都是将个体或组织(如肌肉)作为对象,检测同位素的整体丰度. 目前已知15N浓缩的主要原因是氨基酸代谢中的脱氨基反应有较明显的同位素分馏,导致排泄出更轻的氮同位素[10]. 因此有研究者分离作为生物标记物的氨基酸并测定其δ15N,从中获得与食物链相关的信息,其理论基础是:生物体内以谷氨酸为代表的一类氨基酸可显著富集15N(约8‰),而苯丙氨酸等的15N含量随食物链富集程度较弱(约0.4‰),二者的δ15N值携带了浓缩系数和基线的双重信息[100]. 另有研究[101]认为,利用谷氨酸、脯氨酸、天冬氨酸、赖氨酸等多种氨基酸的δ15N计算营养级可能比仅采用谷氨酸和苯丙氨酸更为准确. 基于氨基酸氮元素的CSIA方法已在食物网生态学中有所应用[102];与此类似,脂肪酸碳元素的CSIA为海洋食物网的食源分析研究、尤其是海底热液系统中能量来源等方面提供了诸多信息[103].随着研究的进一步深入,还出现了针对化合物特定位置元素开展的稳定同位素分析(position-specific isotope analysis, PSIA),以获得更详细的分子内同位素分布信息[104]. 例如,NO2中氮素的PSIA方法可被用于研究硝化和反硝化过程,且在一定程度上推算二者发生的比例[105]. PSIA可以解答CSIA提出的问题,加深人们对元素在环境中归趋与去向的理解.5 结论与展望稳定同位素技术为研究自然和生物过程中的元素迁移转化提供了新手段,其在水环境和水生态领域得到快速发展,在模型方法逐渐完善的同时积累了大量有价值的数据. 但在实际应用中,还存在以下需要深入解决的问题.a) 磷是水生态系统中的重要元素,与水域富营养化关系密切. 然而除31P外,磷的其他同位素均具有放196环 境 科 学 研 究第 35 卷。
亚硝酸盐氧化菌及其生态位的研究进展邢丽贞;程成;孔进【摘要】亚硝酸盐氧化菌(NOB)是氮素转化中重要的一环,对其分类及生态位进行研究,有利于深入了解其生存竞争力和环境适应能力,充分发挥NOB在污水处理领域中的作用.文章阐述了已知NOB的种类及分布,剖析了反应过程中的关键酶NXR,探讨了其不同的代谢方式及其与其他微生物间多样的种间关系,介绍了近年来硝化螺旋菌属中发现的全程硝化菌的研究进展及其对于NOB生态位多样性以及环境适应性的意义,对其研究方向和生态潜力进行了展望.【期刊名称】《山东建筑大学学报》【年(卷),期】2019(034)003【总页数】6页(P62-67)【关键词】亚硝酸盐氧化菌;生态位;代谢方式;种间关系;全程硝化菌【作者】邢丽贞;程成;孔进【作者单位】山东建筑大学市政与环境工程学院,山东济南250101;山东建筑大学市政与环境工程学院,山东济南250101;山东建筑大学市政与环境工程学院,山东济南250101【正文语种】中文【中图分类】X1720 引言氮循环是生态系统中物质能量循环转化的重要组成部分,与生物代谢息息相关,其中亚硝酸盐氧化菌NOB(Nitrite Oxidizing Bacteria)是氮转化中重要的一环,其基本功能是将亚硝氮转化为硝氮,提供自身所需能量的同时推动氮循环的进行,是生态系统必不可少的基本微生物,传统上,认为NOB是生态位相对单一固定的硝化菌。
在经典的硝化二段理论中,NOB与氨氧化菌共同作用,将氨氮转化为亚硝氮,并进一步转化为硝氮,这也是水处理脱氮中的基础步骤和基本理论[1] 。
目前脱氮生物的研究多集中于氨氧化菌AOB(Ammonia Oxidizing Bacteria)或氨氧化古菌AOA(Ammonia Oxidizing Archaea)[2] 与厌氧氨氧化菌[3] ,对亚硝酸盐氧化菌的研究较少,主要有:(1)由于AOB或AOA所作用的氨氧化步骤对于新的脱氮技术尤为重要,成为了脱氮效率的主要限制条件;(2)NOB的生长速度较AOB更加缓慢[4] ,难以得到用于研究的足够的生物量。