蛋白质化学2-3
- 格式:pdf
- 大小:5.59 MB
- 文档页数:220
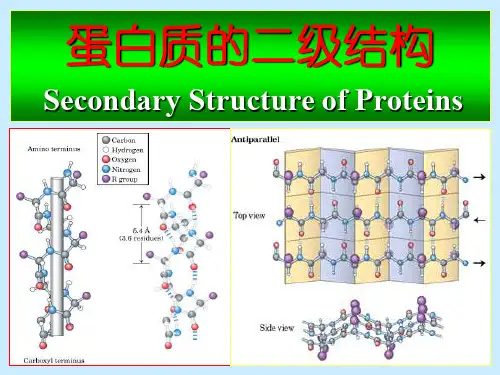
两条或多条几乎完全伸展的多肽链侧 向聚集在一起,相邻肽链主链上的氨 基和羰基形成有规则的氢键,这样的 多肽构象就是-折叠片。
更反 稳平 定行
的 折 叠 片
β-
↓↓
0.35nm
平 行 的
β-
折
↑0.325nm↑
叠
片
β-
折 叠 片
侧链基团交替发布在折叠片的两侧
疏水相互作用
极性侧链 疏水侧链
用作横坐标, 作纵坐标, 作图表示哪 些成对二面角所规定的两个相邻肽单位的 构象是允许的,哪些是不允许的,这个 - 图,就是Ramachandran 构象图(拉氏 构象图)。
从该图可以看出,肽链的折叠具有相当大 的局限性。
(二)二级结构的主要类型
α-螺旋 β-折叠 β-转角
无规则卷曲
β-
转 角
-转角(-turn)
多肽链在折叠成球状蛋白质构象时,第一(n) 个氨基酸残基的羰基和第四(n+3)个残基的氨 基之间形成氢键,形成一种紧密的环,使主链结 构本身以180 °角回折,即-转角。
特点:
多存在于球状蛋白质分子表面 ;
Gly, Pro常存在于-转角处。
H O
-转角
Ⅱ型
Arg-X Lys-X
胰凝乳蛋白酶/糜蛋白酶(chymotrypsin)
断裂Phe、Trp、Tyr等疏水氨基酸残基的羧 基端肽键: Phe-X、Trp-X、Tyr-X优先
化学裂解法
用溴化氰断裂 只断裂由Met残基的羧基参加形成的肽键。 (Met-X)
用羟胺断裂 在pH9下能专一地断裂Asn-Gly之间的肽键, 但Asn-Leu及Asn-Ala键也能部分裂解。
还原法
C-末端AA+硼氢化锂
还原 α- 氨基醇
(肽链完全水解后 用层析法鉴定)
Sanger早期采用此法鉴定胰岛素A、B链的C-末端残基
羧肽酶法
羧肽酶(carboxypeptidase):肽链外切酶, 专一地从肽链的C-末端开始逐个降解,释 放出游离氨基酸。
⑹裂解多肽链成较小的片段
两种或几种不同的断裂方法(断裂点不一样) 将每条多肽链样品降解成两套或几套肽段;
多肽链(亚基)由非共价键缔合: 变性剂如8mol/L尿素、6mol/L盐酸胍、 高浓度盐
多肽链通过二硫桥(S-S)交联: 氧化剂或还原剂将二硫桥断裂
打开后单个多肽链可根据它们的大小或 /和电荷的不同进行分离
不能再重新形成二硫键; Trp被破坏,Met氧化为亚砜
封闭巯基
⑶断开多肽链内的二硫桥
溴化氰作用于Met-X
⑺测定各肽段的氨基酸序列
Edman降解法(自动序列分析仪) 酶降解法 质谱法 根据核苷酸序列的推定法:
推定法与直接氨基酸序列分析结合:
测定蛋白质的N-末端和C-末端,以找到编 码序列(结构基因)
部分氨基酸序列测定,以找出正确的密码阅 读框(reading frame)
② 每圈螺旋沿中心轴方向上升0.54nm, 每个氨基酸残基上升0.15nm;
③特征二面角:=-57 °; =-47°;
④ a-螺旋靠氢键维持,氢键的取向几 乎与中心轴平行;
⑤由一个氢键闭合的环包含13个原子;
⑥a-螺旋中氨基酸残基侧链伸向外侧。
右
手
左
螺
手
旋
螺
旋
亲水的氨基酸残基 疏水的氨基酸残基
N
d+
C
d-
α -螺旋的形成受多种因素制约
某些氨基酸如脯氨酸和甘氨酸可以破坏螺旋; 大侧链氨基酸影响螺旋的形成; 离子间的作用影响螺旋形成; 肽键的平面性和反式构象的影响。
其他螺旋结构
310-螺旋 π -螺旋(4.416-螺旋) 左手α-螺旋
β-折叠
β-折叠股(β- strand) β-折叠片(β-pleated sheet)
全α-结构域
全β -结构域
α/β结构域
α+β结构域
结 构 域 的 一 些 类 型
六.蛋白质的三级结构
(一)概念:
多肽链借助各种次级键和二硫键,通过 盘绕折叠,形成具有特定肽链走向的紧密球 状构象,称为三级结构。
蛋白质的三级结构指蛋白质分子或亚基 内所有原子的空间排布,但是不包括亚基间 或不同分子间的空间排列关系。
-折叠片特点:
1. 所有的肽键都参与链间氢键的交联,氢键与 肽链的长轴接近垂直;
2. 多肽主链取锯齿状折叠构象;
3. 侧链交替分布在片层平面的两侧;
4. -折叠片有平行式和反平行式;平行式的 -折叠片中,两个氨基酸残基之间的距离为 0.325nm,反平行式中为0.35nm。
β-转角(β-turn)
测定每个小肽段的序列
多肽链的结构重建
确定完整多肽链的序列 确定二硫键的位置、酰胺的位置
蛋白质序列测定的一般策略:
⑴测定蛋白质分子中多肽链的数目 ⑵拆分蛋白质分子的多肽链 ⑶断开多肽链内的二硫桥 ⑷分析每一多肽链的氨基酸组成 ⑸鉴定多肽链的N-末端和C-末端残基 ⑹裂解多肽链成较小的片段 ⑺测定各肽段的氨基酸序列 ⑻重建完整多肽链的一级结构 ⑼确定半胱氨酸残基间形成的S-S交联桥的位置
N末端是Glu(E)
T-2位于氨基端(E开头); T-3 位于羧基端(末尾不是 R或K)
C-3 与T-1 及T-4 有重叠, 可确定序列
确定二硫键位置等
(三)蛋白质一级结构举例
• 1953年 Sanger确定
• 51个氨基酸残基(21+30)
• 分子量为 5700Da
(胰岛素)
•二硫键
二硫键的形成
"for their studies of the structures of globular proteins"
Hb
(1937-1960)
Perutz et al. (1960) Nature 185:416
Mb
(1947- 1960)
Kendrew et al. (1960)
⑻重建完整多肽链的一级结构
利用两套或多套肽段的氨基酸序列彼此间交 错重叠拼凑出完整序列
⑼确定半胱氨酸残基间形成的S-S交联桥 的位置
胃蛋白酶水解 对角线电泳分离 注意:书上26页图2-12,“第一向”与“第
二向”应对换
共38个氨基酸残基; 1 Arg (R),2 Lys(K) 2 Met (M)
(二)蛋白质一级结构的测定
1.蛋白质序列测定的一般步骤
测序前的准备工作 序列测定 多肽链的结构重建
测序前的准备工作
纯化蛋白质 测定蛋白质的分子量 确定蛋白质的肽链数目或亚基数目 测定其氨基酸组成 配基的确定 末端氨基酸残基的分析
序列测定
用至少两种不同方法断裂多肽链并分离小肽 段
α-螺旋 (α-helix)
N C
肽链由于氨基酸残基的氨基氢和它后面第四个残基上 的羰基氧之间形成氢键,呈周期性规则的螺旋状构象, 即a-螺旋。
第n个氨基酸残基的氨基氢与第n+4个残基上的羰基氧
α -螺旋氢键的形成
a-螺旋(3.613-螺旋)的基本特征: ① a-螺旋为右手螺旋,每圈螺旋含3.6个 氨基酸残基;
C-末端测定的方法
肼解法 还原法 羧肽酶法
肼解法
蛋白质/多肽
无水肼 100℃
(n-1)个氨基酸酰肼+C-末端氨基酸
+苯甲醛
二苯基衍生物(沉淀)
上清中游离C-末端氨基 酸可用FDNB法或DNS 法、层析技术进行鉴定
肼解过程中,谷氨酰胺、天冬酰胺、半胱氨酸 等被破坏不易测出
C-末端的精氨酸变为鸟氨酸
第四节 蛋白质的分子结构
一、蛋白质的结构层次
蛋白质的一级结构 蛋白质的二级结构 蛋白质的超二级结构 蛋白质的结构域与三级结构 蛋白质的四级结构
二、蛋白质的一级结构
(一)蛋白质一级结构的定义: 指氨基酸在肽链中的排列顺序。
注意蛋白质一级结构和共价结构的区别:
蛋白质的共价结构包括肽链的数目、末端氨 基酸残基组成、氨基酸排列顺序及二硫键位 置等内容。
(发夹):两个反平行的β折叠股由转角或无规 卷曲相连。
多个发夹结构可形成β迂回或希腊钥匙结构。 a:两段平行β折叠股由一段a-螺旋连接。
Rossman折叠 fold: aa
The structure of hair and hair a- keratin
aa-structure
⑴测定蛋白质分子中多肽链的数目
根据蛋白质N-末端或C-末端残基的 摩尔数和蛋白质的分子量:
若蛋白质只含一条多肽链,则蛋白质的摩尔数= 末端残基的摩尔数
若末端残基的摩尔数是蛋白质的摩尔数的倍数, 则该蛋白质由多条多肽链组成
若末端残基多于一种,则蛋白质由两条或多条不 同的多肽链组成
⑵拆分蛋白质分子的多肽链 (寡聚蛋白质)
牛
胰
核
糖
核
酸
酶
的
氨
ห้องสมุดไป่ตู้
基
酸
顺
序
•50年代末Moore等完成;
•一条多肽链,124个氨基酸;
•分子量为12600Da;
•分子内含有4个二硫键
三、蛋白质的二级结构
二级结构的概念:
指多肽链主链有规律的折叠和盘绕,是多 肽链主链局部的空间排列。维系二级结构的 主要作用力是氢键。
(一)多肽链主链构象的空间限制
一对二面角决定了相邻两个肽单位的相对位 置。
a-C 原子为肽平面间的连接点
F= 0° = 0°
多肽链折叠的空间限制
二 面
F:
Ca
-N
角 Y: Ca - C
2.二面角F, 对主链构象的限制
F= 180° = 180°