第四讲 乳酸菌类型及生物活性次生代谢产物
- 格式:doc
- 大小:68.50 KB
- 文档页数:9

乳酸菌的功能·应用及大量增殖生产技术首先我们来认识一下乳酸菌是什么,以及它有什么特点及功能,这样就使我们能对乳酸菌的市场前景具有很好的认识:一. 乳酸菌简介什么是乳酸菌乳酸菌指发酵糖类主要产物为乳酸的一类无芽孢、革兰氏染色阳性细菌的总称。
凡是能从葡萄糖或乳糖的发酵过程中产生乳酸菌的细菌统称为乳酸菌。
乳酸菌存在于动物及植物中,在国外有时被称为植物性乳酸菌和动物性乳酸菌,从分类学来讲没有具体的分类,因为不管是以葡萄糖和乳糖为食的乳酸菌、他们的遗传因子是没有区别的。
乳酸菌是一群相当庞杂的细菌,目前至少可分为18个属,共有200多种, 从形状上来看有乳酸杆菌和乳酸球菌,单个细胞的大小约0.5-10μm 。
除极少数外,其中绝大部分都是人体内必不可少的且具有重要生理功能的菌群,目前已被国内外生物学家所证实,其广泛存在于人体及动物的肠道以及各种植物中。
而肠内乳酸菌与健康长寿有着非常密切的直接关系。
乳酸菌在有氧环境能够生存,但是喜欢在氧气很少的环境生存,是通性嫌气性菌。
中文学名:乳酸菌 界:细菌界 门:厚壁菌门Firmicutes 纲: 芽孢杆菌纲Bacilli在很久很久以前乳酸菌就和人类有着关联了,奶酪(是奶乳经乳酸菌发酵后的食品)在大约8000年前在格里斯河流和发拉底河流之间的狭长地区(现在的伊拉目: 乳杆菌目Lactobacillales 科: 乳杆菌科Lactobacillaceae 属: 乳杆克)就被生产了,这就是最早的乳酸菌食品。
在中国历史上和乳酸菌的渊源也很早,东汉末年,中国大约有数百万人死于霍乱病的肆虐传播。
公元200-205年,张仲景因而编撰成书具有划时代意义的《伤寒论》,在书中总结了艾蒿草的治疗方法。
现代科学的研究表明,起到神奇功效的其实是艾蒿草中富含的乳酸菌。
正是乳酸菌控制了瘟疫的蔓延,奇迹般地解救了中华民族。
在现代社会,乳酸菌的应用带来了一场乳酸菌的革命,中国科学院金锋教授的著书《乳酸菌革命》(日本株式会社评言杜2009年出版)中指出,研究表明:乳酸菌在工业、农业、畜牧业、医药及人们的日常生活中有着广泛效用。

初生代谢产物和次生代谢产物的概念初生代谢产物和次生代谢产物的概念1. 初生代谢产物和次生代谢产物的定义初生代谢产物和次生代谢产物是生物体内产生的两种不同类型的化合物。
初生代谢产物是在生物体内发育的早期阶段产生的化合物,主要用于维持生命和促进生长的基本代谢功能。
而次生代谢产物是在生物体内发育的后期阶段产生的化合物,不参与生物体的基本代谢,但具有一定的生理活性和适应性。
2. 初生代谢产物和次生代谢产物的生物功能初生代谢产物主要包括碳水化合物、蛋白质和脂类等生物大分子,以及氨基酸、酶、激素等生物小分子。
它们是维持生物体正常生长发育和代谢活动所必需的物质,是构成细胞、组织和器官的基本组成部分。
而次生代谢产物则包括生物碱、鞣质、挥发油、色素等化合物,具有抗菌、抗虫、抗氧化、防御等生理活性,在植物的适应环境和保护自身方面发挥着重要作用。
3. 初生代谢产物和次生代谢产物在生物体中的制备和调控初生代谢产物一般是通过生物体内的基础代谢途径合成的,如糖酵解、蛋白质合成、脂质代谢等。
它们的合成受到生物体内外环境的调控,如营养物质的供应、激素的调节等。
而次生代谢产物的合成一般是在特定的生物发育阶段或环境刺激下进行的,受到内在遗传和外部环境因素的影响,通常在生物体受到外界胁迫时产生。
4. 个人观点和理解在我看来,初生代谢产物和次生代谢产物在生物体内发挥着各自独特的作用。
初生代谢产物是维持生命的基础物质,是生物体正常生长和代谢活动不可或缺的。
而次生代谢产物则是植物为了适应环境和防御外界威胁而产生的重要物质,对于保护植物自身和与外界的相互作用至关重要。
总结回顾初生代谢产物和次生代谢产物作为生物体内重要的化合物,分别在维持生命和适应环境方面发挥着重要作用。
初生代谢产物是生物的基础代谢产物,次生代谢产物则是在特定条件下产生的具有生理活性的化合物。
这两种代谢产物相辅相成,共同维护着生物体的正常功能和适应性。
在撰写文章时,我尽力按照所提供的要求,以简单到复杂的方式全面评估了初生代谢产物和次生代谢产物的概念,希望这篇文章对您有所帮助。

乳酸菌有氧产物
乳酸菌是一类革兰氏阳性杆菌,通常在厌氧条件下发酵产生乳酸,这
是它们的主要产物。然而,在某些情况下,乳酸菌也可以在有氧条件
下进行代谢活动,产生一些其他有氧产物。
在有氧条件下,乳酸菌可能会产生以下一些有氧产物:
乳酸:即使在有氧条件下,乳酸菌仍然会产生少量乳酸。这是因为乳
酸菌的代谢途径中乳酸脱氢酶的活性会增加,导致乳酸的产生。
乙醇:在有氧条件下,乳酸菌有时也会产生少量乙醇,这是因为在氧
气存在的情况下,乳酸可以进一步代谢为乙醇和二氧化碳。
二氧化碳:无论在有氧还是厌氧条件下,乳酸菌都会产生二氧化碳作
为代谢产物之一。
需要注意的是,乳酸菌在有氧条件下产生这些有氧产物的能力相对较
弱,且产量较少。乳酸仍然是其主要的产物。这些有氧产物的生成受
到很多因素的影响,包括菌株的种类、培养条件、营养成分等。

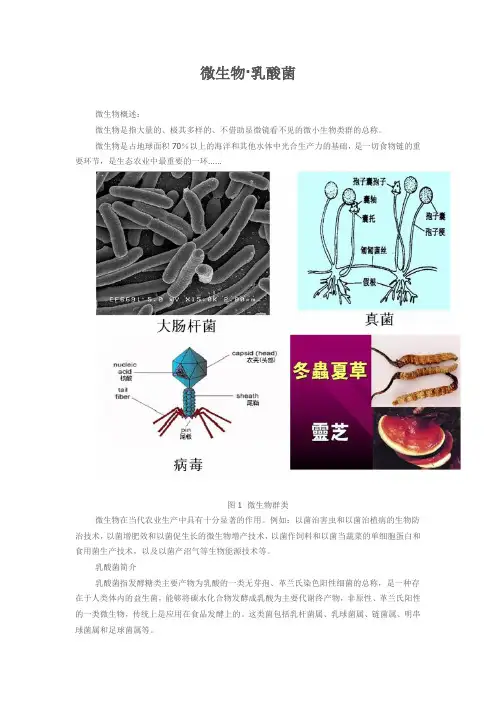
微生物·乳酸菌
微生物概述:
微生物是指大量的、极其多样的、不借助显微镜看不见的微小生物类群的总称。
微生物是占地球面积70%以上的海洋和其他水体中光合生产力的基础,是一切食物链的重要环节,是生态农业中最重要的一环……
图1 微生物群类
微生物在当代农业生产中具有十分显著的作用。
例如:以菌治害虫和以菌治植病的生物防治技术,以菌增肥效和以菌促生长的微生物增产技术,以菌作饲料和以菌当蔬菜的单细胞蛋白和食用菌生产技术,以及以菌产沼气等生物能源技术等。
乳酸菌简介
乳酸菌指发酵糖类主要产物为乳酸的一类无芽孢、革兰氏染色阳性细菌的总称,是一种存在于人类体内的益生菌,能够将碳水化合物发酵成乳酸为主要代谢终产物,非原性、革兰氏阳性的一类微生物,传统上是应用在食品发酵上的。
这类菌包括乳杆菌属、乳球菌属、链菌属、明串球菌属和足球菌属等。
图2 乳酸菌主要类群
大量研究资料表明,乳酸菌能促进动物生长,调节胃肠道正常菌群、维持微生态平衡,从而改善胃肠道功能;提高食物消化率和生物效价;降低血清胆固醇,控制内毒素;抑制肠道内腐败菌生长,提高机体免疫力等。
此外,乳酸菌的代谢产物乳酸是一种天然发酵酸,具有还原性,可增加食物的美味和产品风味,且有很强的防腐保鲜功效,能延长保质期。


引言概述:微生物乳酸菌是一类广泛存在于自然界和人体内的微生物,具有重要的生物学功能和应用价值。
它们是一类可产酸的革兰氏阳性菌,能够利用碳水化合物产生乳酸。
微生物乳酸菌在食品工业、医学和农业等领域有着广泛应用,具有调节肠道菌群、增强免疫力、抗菌和防腐等作用。
本文将从微生物乳酸菌的分类、生物学特性、功能和应用等方面进行详细阐述。
正文内容:一、微生物乳酸菌的分类1.乳酸菌的基本特征乳酸菌是一类革兰氏阳性菌,能够利用碳水化合物进行乳酸发酵。
其基本特征包括形态特征、生理特性和生物化学性质等方面。
2.微生物乳酸菌的分类与系统发育微生物乳酸菌是按不同的分类体系进行分类的,常见的分类体系有传统的形态分类和现代的分子分类等。
二、微生物乳酸菌的生物学特性1.微生物乳酸菌的形态及培养特性微生物乳酸菌的形态多样,包括球形、杆状和弯曲杆状等。
在不同的培养基和条件下,它们具有不同的培养特性和生长速度。
2.微生物乳酸菌的代谢途径微生物乳酸菌主要以碳水化合物为底物进行乳酸发酵代谢,同时还可以进行其它代谢途径,如产生酒精、乙酸和丙酸等。
三、微生物乳酸菌的功能1.调节肠道菌群微生物乳酸菌能够通过抑制有害菌的生长和增加有益菌的数量来调节肠道菌群,保持肠道微生物平衡,从而促进肠道健康。
2.增强免疫力微生物乳酸菌具有抗菌和免疫调节作用,能够增强人体免疫力,促进免疫细胞的活化和增殖,从而提高机体的抵抗力。
3.抗菌和防腐微生物乳酸菌能够产生酸和抗菌物质,具有抑制细菌和真菌生长的作用,可以用于食品工业中的防腐和保鲜。
四、微生物乳酸菌的应用1.食品工业中的应用微生物乳酸菌广泛应用于食品工业,如酸奶、乳酸发酵面包、酸奶饮料等。
它们能够增强食品的营养价值,改善口感,并有助于消化和吸收。
2.医学和药物领域中的应用微生物乳酸菌在医学和药物领域中也有重要应用,如用于治疗胃肠道疾病、预防和治疗感染性疾病、制备益生菌药品等。
3.农业领域中的应用微生物乳酸菌可用于农业领域,如作为土壤增效剂、生物防治剂和饲料添加剂,起到改善土壤质量、增强作物抗病能力和促进动物生长的作用。

风味发酵乳中乳酸菌代谢产物的生物活性研究乳酸菌是一类常见于乳制品中的益生菌,其发酵产物在食品行业中被广泛应用。
风味发酵乳是一种受欢迎的乳制品,它不仅具有丰富的口感和风味,还含有许多有益于健康的成分。
乳酸菌在发酵过程中生成的乳酸是主要的发酵产物,它除了为产品提供了特殊的风味外,还具有多种生物活性。
乳酸是一种有机酸,是乳酸菌发酵过程中生成的主要产物。
研究表明,乳酸具有抑制病原微生物生长的能力。
乳酸能够降低肠道pH值,创造不利于致病菌生存的环境。
此外,乳酸还可以提高肠道内有益菌的数量,进一步增强肠道的健康状况。
肠道是人体最大的免疫器官之一,乳酸通过改善肠道健康来增强免疫功能,减少患病的风险。
此外,风味发酵乳还含有其他乳酸菌代谢产物,如乳酸菌的胞外多糖、胞内多糖等。
这些多糖物质具有多种生物活性,如抗氧化、抗菌、降血压等。
胞外多糖经过适当的提取工艺后可以作为功能性食品添加剂,具有增强人体免疫力和调节血糖功能。
胞内多糖则可以作为纤维素摄入,对肠道有益。
此外,还有一些乳酸菌代谢产物在风味发酵乳中具有重要的生物活性。
例如,维生素B族是乳酸菌发酵过程中产生的重要物质之一。
维生素B族对人体的代谢具有重要的调节作用,缺乏维生素B族会导致一系列健康问题。
通过食用风味发酵乳,可以补充人体对维生素B族的需求,增强机体的新陈代谢,维持身体的正常运转。
另外,还有一些乳酸菌代谢产物具有抗炎和抗肥胖的作用。
乳酸菌的发酵产物中含有丰富的一种叫做肽的物质。
这些肽具有一定的抗炎作用,有助于减轻炎症反应,改善炎症相关的疾病。
同时,它们还可以抑制脂肪细胞的增殖和增加脂肪氧化,从而有助于控制体重和预防肥胖。
这些生物活性物质的存在,使得风味发酵乳成为一种有益于健康的食品选择。
通过对风味发酵乳中乳酸菌代谢产物的生物活性研究,可以深入了解其对人体的益处。
乳酸菌发酵产物的生物活性给人们带来了许多健康福利。
食用风味发酵乳可以改善肠道健康、增强免疫功能、调节血糖、抗炎和抗肥胖等。

乳酸菌知识点总结乳酸菌作为一种非常重要的微生物,其在人类的健康维护和食品工业中扮演着不可或缺的角色。
本文将从乳酸菌的定义、分类、生理功能、应用等各方面进行深入探讨,希望能够对读者更好地了解乳酸菌有所帮助。
一、乳酸菌的定义及分类乳酸菌是一类革兰氏阳性、非芽孢杆菌,产生乳酸为代谢产物,能够发酵多种碳水化合物,这使得其在食品发酵和生物医学领域中广泛应用。
根据乳酸生成产物和代谢途径的不同,乳酸菌可以分为两类:homofermentative乳酸菌和heterofermentative乳酸菌。
前者主要产生乳酸,而后者还会产生一定量的乙醛、二氧化碳等物质。
乳酸菌也可以根据表现形式、营养特性等方面进行分类,如Lactobacillus、Bifidobacterium、Streptococcus、Lactococcus等。
其中Lactobacillus是最具代表性的乳酸菌之一,其在多种食品中被广泛使用。
二、乳酸菌的生理功能1. 食品发酵乳酸菌适应性广泛,能够使用各种碳水化合物进行发酵,进而改善食品的口感和品质。
比如,Lactobacillus acidophilus可以在牛奶中产生乳酸和乳酸菌素,这有助于保持牛奶的新鲜和稳定。
2. 肠道正常菌群的维护乳酸菌在人类肠道中存在,并且作为一种益生菌,乳酸菌可以增强肠道的免疫力,维护肠道正常菌群的平衡。
此外,乳酸菌还可以促进肠道蠕动,减少便秘。
3. 抑制有害菌滋生乳酸菌可以通过产酸和抗菌素等机制,抑制肠道中有害菌的生长繁殖,维持肠道微生态系统的平衡,减轻或预防肠道疾病,如腹泻和炎症性肠病等。
4. 抗氧化作用乳酸菌中含有丰富的抗氧化物质,可以清除自由基,减缓细胞老化,同时增强机体的免疫力,预防各种疾病。
三、乳酸菌在食品和医药行业中的应用1. 食品工业乳酸菌在食品工业中被广泛应用,包括乳制品、酸奶、酸菜等。
比如,Lactobacillus reuteri可以用于生产乳清酸饮料,而Streptococcus thermophilus则常用于生产奶酪和酸奶等。

乳酸菌的简介乳酸菌的简介⼀、乳酸菌的起源根据圣经旧的书上记载,公元前4000多年,⼈类已经开始发酵乳酸⾁制品及蔬菜腌渍物。
直到公元1857年,巴斯德(Pasteur)发现酸乳(sour milk)中有微⼩⽣物体存在,将其定名为”levue lactique”,发酵乃微⽣物作⽤所致的秘密才⾸次得以揭露;这是发现乳酸菌的开端。
1873年李斯特(Lister)利⽤稀释法,由酸乳中纯化分离出Bacterium lactis,也就是⽬前的Lactococcus lactis,这是乳酸菌最早被分离出来的纪录(廖,1998;李,2000)。
⼆、乳酸菌的定义乳酸菌⼀般是指能将碳⽔化合物发酵分解为乳酸的细菌群(佐佐⽊隆,1998)。
乳酸菌群具有以下⼏点特性:1.为⾰兰⽒阳性(gram positive)球菌或杆菌。
2.不产⽣孢⼦(nonsporting)且⽆运动性(nonmotile)3.不具分泌催化酶(catalase negative)之能⼒。
4.可在有氧环境⽣长,但以⽆氧状态下⽣存较佳,亦有绝对厌氧者。
5.需有碳⽔化合物、胺基酸、维⽣素等多种⽣长因⼦⽅能⽣长之复合营养需求性(complex nutritional requirements)。
6.依代谢途径与最终产物的不同,可分为同质发酵(homofermentative)及异质发酵(heterofermentative)两种乳酸菌。
同质发酵性乳酸菌含有aldolase,最终产物90%-100%为乳酸;异质发酵性乳酸菌具有phosphoketolase,其最终产物除了40%-50%的乳酸外,还包括⼄醇、⼆氧化碳及醋酸等多项产物(Ingledew,1995)。
乳酸菌普遍存在动物体消化道中,因耐酸性佳且可分泌乳酸及抗菌物质等有利于⽣存的条件,故消化道三⼤菌群当中最占优势(Nemcova et al. 1997)三、乳酸菌的分类乳酸菌⼀般包括Lactobacillus (L)、Leuconostoc (Leuc.)、Streptococcus (S)及Pediococcus (Ped.)四属;⼴义的乳酸菌尚包括Bifidobacterium与Sporolactobacillus 两个属(Jay, 2000)。

乳酸杆菌代谢生
物及其在人体中的作用
乳酸杆菌是一种常见的益生菌,它们可以通过发酵代谢生产乳酸、醋酸、乙酸等有机酸,同时还能产生一些抗菌物质和酶类。
这些代谢产物在人体中具有多种作用。
首先,乳酸杆菌代谢产生的有机酸可以降低肠道pH值,抑制有害菌的生长,维护肠道微生态平衡。
此外,有机酸还可以促进肠道蠕动,增加粪便体积,预防便秘。
其次,乳酸杆菌代谢产生的抗菌物质可以直接抑制有害菌的生长,减少它们对人体的危害。
同时,这些抗菌物质还可以增强人体免疫力,提高身体抵抗力。
此外,乳酸杆菌代谢产生的酶类可以帮助人体消化吸收食物中的营养物质,促进肠道健康。
其中,乳酸杆菌产生的乳糖酶可以帮助人体消化乳糖,缓解乳糖不耐受症状。
最后,乳酸杆菌代谢产生的一些代谢产物还具有抗氧化、抗炎、降血压等作用,有助于预防多种疾病。
总之,乳酸杆菌代谢生产的有机酸、抗菌物质和酶类等代谢产物在人体中具有多种作用,可以维护肠道微生态平衡,增强人体免疫力,促进消化吸收,预防多种疾病。
因此,适当补充乳酸杆菌对于人体健康非常重要。
可以通过食用含有乳酸杆菌的食品,如酸奶、酸菜、酸奶菜等,或者选择乳酸杆菌制剂进行补充。
当然,在选择乳酸杆菌制剂时,应该选择质量可靠、来源清晰的产品,避免不必要的风险。
乳酸菌代谢途径的研究乳酸菌是一类重要的微生物,常被用于发酵食品,比如酸奶、酸酵素乳等。
它们在乳制品工业中起着关键作用,对人类的健康也有明显的益处。
然而,乳酸菌的代谢途径一直是研究的热点之一。
本文将探讨乳酸菌代谢途径方面的最新研究进展。
1、乳酸菌代谢途径的基本情况乳酸菌是一种革兰氏阳性杆菌,存在于自然界中的各个角落。
与酵母菌和细菌不同的是,乳酸菌的代谢途径比较简单,主要分为两种类型:异养代谢和自养代谢。
在自养代谢中,乳酸菌能够通过光合作用产生ATP,而异养代谢则依赖于外源性的有机物质。
2、乳酸菌代谢途径的研究进展近年来,随着分子生物学和代谢组学技术的发展,乳酸菌代谢途径的研究也取得了新的进展。
研究者通过对乳酸菌的基因组序列和代谢产物的分析,揭示了其代谢途径的调控机制和代谢产物的合成过程。
下面我们来详细讨论一下现有研究成果。
2.1 乳酸菌的葡萄糖代谢途径葡萄糖是乳酸菌生长的重要碳源之一,它通过代谢途径被分解为丙酮酸和乳酸。
在过去的研究中,乳酸菌的葡萄糖代谢途径被认为是通过Embden-Meyerhof途径进行的。
然而,最近的研究表明,乳酸菌的葡萄糖代谢途径可以是不同的。
一些乳酸菌可以通过Pentose Phosphate Pathway(PPP)来代谢葡萄糖。
PPP由糖醛酸途径和废物消化途径组成,能够产生更多的NADPH和核酸前体。
因此,通过PPP途径代谢葡萄糖的乳酸菌能够在缺氧的条件下生长,同时也能够合成足量的核酸前体。
2.2 乳酸菌产生乳酸的代谢途径乳酸是乳酸菌代谢产物中最为重要的一种,它在酸奶等乳制品生产中发挥着不可替代的作用。
目前已有一些乳酸菌的乳酸合成代谢途径被揭示。
指路糖酸代谢路径是一种在大多数乳酸菌中发现的代谢途径。
在这个途径中,葡萄糖经过葡萄糖激酶的磷酸化后进入到糖醇磷酸途径,在糖醇磷酸途径中被转化为L-乳酸。
虽然指路糖酸代谢路线是乳酸合成的主要途径之一,但仍有一些乳酸菌在此过程中合成出有机酸和二氧化碳等不同的产物。
第四讲乳酸菌及益生菌种类的功能及具有生物活性的次生代谢产物一、乳酸菌及益生菌1、定义及分类(1)乳酸菌(2)益生菌2.生物活性3.生长特性及应用、产品开发二、生物活性的次生代谢产物1、细菌素(1)定义及种类(2)特性Nisin是正常菌群中某些种类的乳酸乳球菌合成和分泌的一种对大多数G+菌有强大杀灭作用的细菌素。
Nisin对营养细胞的作用主要是在细胞膜上,它可以抑制细菌细胞壁中肽聚糖的生物合成,使细胞质膜和磷脂化合物的合成受阻,从而导致细胞内物质外泄,甚至引起细胞裂解。
作为一种天然的生物性食品防腐剂和抗菌添加剂,Nisin已广泛应用于乳制品、罐头食品、高蛋白食品及饲料工业中。
添加400IU/g的产品储存11天,其含菌数比对照组低4个对数期,产品的感官可接受性较高。
Nisin对乳酸菌株的影响也较大,如控制适宜的添加量(一般在5-100mg/kg),能对某些乳酸菌株有明显的抑制作用,造成菌株的活力下降、产酸受抑、生长不良,但乳酸菌总数变化不大。
nisin的研究现状乳链菌肽是由乳酸乳球菌产生的由基因编码的多肽抗生素,含有34个氨基酸残基,分子中含有5种稀有氨基酸,即氨基丁酸(ABA)、脱氢丙氨酸(DHA)、β-甲基脱氢丙氨酸(DHB)、羊毛硫氨基酸( ALA-S-ALA)和β-甲基羊毛硫氨基酸(ALA-ABA),它们通过硫醚键形成五个内环。
乳链菌肽的抑菌谱包括营养体和芽孢,对葡萄球菌、梭菌、芽孢杆菌和利斯特菌有强烈的抑制作用,对酵母和霉菌没有作用。
迄今为止,乳链菌肽已在全世界约60多个国家和地区被用作食品防腐剂]。
我国对乳链菌肽的研究始于1989年,中国科学院微生物研究所在国家自然科学基金项目和中国科学院“八五”重点科研基金项目资助下,完成了对乳链菌肽的基础研究,研究成果现已批量生产,投放市场。
1992年3月29日,我国卫生部食品监督部门签发了乳链菌肽在国内的使用合格证明,同时将乳链菌肽列入1992年10月1日实施的国标GB2760-86中的增补品种,可用于罐藏食品、植物蛋白食品、乳制品和肉制品的保藏中。
乳链菌肽用于食品中的优点是安全无毒,它的LD50约为7000Kg,允许使用的最大剂量是33000IU/Kg,乳链菌肽是一种短肽,可以被体内的α-胰凝乳蛋白酶降解;不影响食品的色香味;在食品加工中可以降低杀菌温度、减少热处理时间,改变食品的营养价值、风味和结构等性状。
国内外的研究者在对乳链菌肽的分子结构、分子形成过程、物理化学性质、抗菌机理和毒性等进行了广泛的研究产nisin菌株在发酵食品中的应用在干酪中的应用早在1951Hirsch等研究发现nisin产生菌乳酸乳球菌乳酸亚种在Swiss型干酪中抑制厌氧产气芽孢的形成。
nisin最早应用于干酪的防腐,也是目前乳制品应用中的一个主要方面。
在干酪加工的过程中,虽经过高温灭菌,但仍会有耐热性的革兰氏阳性菌孢子(如肉毒梭状菌和厌氧的梭菌芽孢)存在,乳酸链球菌肽可有效地阻止这些孢子的萌发和毒素的形成,有报道说,将nisin抗性株和nisin产生菌混合用于干酪发酵剂,可使优质产品率达90%以上,而用常规的方法仅有41%。
学者们在这方面进行了大量的研究,nisin产生菌在干酪中应用的主要限制是生长缓慢和产酸率低。
Exterkate等(1976)产酸率低主要原因是缺乏蛋白酶系统。
是这些nisin产生菌只能利用乳中的可溶性含氮化合物。
Roberts等(1992)用电穿空法将质粒pFG010引入乳酸乳球C2中使其产生nisin抗性和红霉素抗性,然后将其与nisin 产生菌混合发酵,以期改变干酪的产酸度,但并未取得成功,由于这个质粒译码的抗性新陈代谢导致nisin失活。
Roberts等(1992)通过筛选发酵蔗糖nisin产生菌菌株获得一株自发突变的蛋白酶阳性菌株NCDO1404,用1%的NCDO1404和0.5%JS102混合用于cheddar干酪发酵,使产酸率提高,达到干酪的酸度要求,干酪中大约700IU/g的nisin。
此后,Roberts 等(1993)又将用nisin产生菌制作的干酪用于巴氏杀菌的干酪酱的一种配料成份,结果低湿度在22和37℃,高湿度在22℃贮存时,延长了货架期。
Berg等(1998)对Manchego干酪在成熟期间产nisin的乳酸乳球菌乳酸亚种ESI515对Listeria Innocua的抑制性进行了研究,发现成熟60天后,产nisin的发酵剂发酵干酪中L.innocua比不产nisin发酵剂酵的干酪细菌总数少约104个。
Rodríguez等(2001)]等通过杂交的方法获得一株nisin产生菌菌株TAB50,并将其用于抑制半硬质干酪中单核增生性李斯特菌。
Maisnier-patin等(1992)也将其用于Camember干酪中。
Rodríguez等(1998)[13]将其用于Manchego干酪均证明对李斯特菌有良好抑制作用。
使用产乳酸链球菌的作为发酵剂用于干酪的生产能达到抑菌的目的,其风味并未受到影响。
Abdelghani等(2001)将nisin产生菌与其他乳酸菌用于Hispßânico干酪的发酵发现加入nisin产生菌与不加产nisin发酵剂的干酪的pH值没有差别,菌落总数低。
每1Kg接种1.0gnisin产生菌发酵的干酪在成熟45天后,其蛋白水解度和游离氨基酸的数量分别是不加产nisin发酵剂的干酪的1.8倍和2.7倍,疏水的肽降低,3-甲基-1-丁醛、丁二酮和乙酰甲基甲醇浓度增加,干酪风味更好。
在泡菜中的应用Harris等(1992)将nisin产生菌乳酸乳球菌NCK401和nisin抗性的Leuconostoc mesenteroides NCK293混合用于泡菜的发酵中。
泡菜在最初发酵阶段是异型发酵的L占主导地位,由于L对酸敏感,随着酸度的上升,L迅速死亡,在发酵的后期是植物乳杆菌占主导。
这种微生物生长顺序使泡菜具有特殊的风味和香味,然而,在大批量的生产中,泡菜的这种自然发酵主要信赖于原料的微生物种群。
微生物的生长顺序受到生产条件的影响。
因此,很难扩大生产并维持泡菜质量的稳定。
乳酸乳球菌NCK401产生的nisin可以在初期抑制L 以外的其它微生物的生长,而不影响LNCK293的生长繁殖。
在混合发酵剂中100AU /ml的nisin轻微抑制植物乳杆菌ATCC14917。
600AU/ml可以完全抑制株菌的发酵。
由于后期的同型发酵是泡菜发酵中必不可少的,因此从实践的观点来看nisin并不是消除植物乳杆菌的生长。
通过实验将乳酸乳球菌和L比例定为103:1。
这种泡菜在含有2%的NaCl在20℃发酵获得了较高质量。
在酸奶中应用与干酪相比,nisin产生菌用于酸奶的发酵的研究应用较少,只有Kalra等(1975)研究了nisin产生菌与其它菌株混合发酵来阻止酸奶中酸的增加,以后他又研究了乳酸菌对nisin 的敏感性,及nisin产生菌对其他普通发酵剂的影响。
nisin产生菌在干酪中的应用已取得了可喜的成功,在泡菜和酸奶的应用中也有了初步的研究。
到目前为止,还未见产nisin的乳酸乳球菌用于其他发酵食品中,如果将nisin产生菌与其他nisin抗性菌株用于发酵食品中,可以抑制其它不需要的微生物的生长,减少或不必添加防腐剂,改善品质,延长保质期。
国内将产nisin的菌株用于发酵食品中还未见报道。
随着对nisin产生菌的特性的进一步的研究,产nisin的菌株在发酵食品中的应用必将大放异彩。
2、乳酸菌胞外多糖的研究进展多糖是近年来兴起的一个研究热点,掀起了继蛋白质和核酸之后的又一生物大分子的研究热潮。
人们之所以如此重视它,是因为它在生物体内所起的复杂且重要的生理作用。
近年来,由于许多发达国家诸如美国、日本和欧洲等对多糖研究的重视,使得这方面的研究取得了不少进展,使得人们对它有了更深的了解(顾瑞霞,2000)。
多糖是高度水合的聚合物,由重复单元(由单糖组成)以糖苷键链接而成,分为同聚多糖和杂多糖,其分子上还可以带有无机分子和有机分子的取代基。
多糖是一些变化多端的生物大分子,因为不但组成它们的单糖单位可以变化,而且这些单糖单位的链接方式也不定。
单糖上存在一些羟基,可以形成糖苷键,这就意味着任何两个单糖就可以以多种位置相连。
与之相比,任何两个氨基酸只能以两种方式链接成二肽。
此外,多糖中还存在许多支链,以及无几分子和有机分子的取代基,致使其结构尤为复杂(Roberts,1995)。
由此可见,复杂的多糖必定与复杂的生命现象关系密切。
生物界中,动物、植物和微生物都是多糖的丰富来源。
微生物的胞外多糖尤为重要。
微生物多糖包括某些细菌、真菌和蓝藻类产生的多糖,主要以三种形式存在:粘附在细胞表面上;分泌到培养基中;构成细胞的成分。
微生物多糖,因其安全无毒、理化性质独特等优良性质而倍受关注。
微生物多糖包括胞内多糖、胞壁多糖和胞外多糖。
胞外多糖是由微生物大量产生的多糖,易与菌体分离,可通过深层发酵实现工业化生产。
据D.E.Eveleigh统计,已经发现49属76种微生物产生胞外多糖,但真正有应用价值并已进行或接近工业化生产的仅十几种。
近几年,随着对微生物多糖研究的深入,世界上微生物多糖的产量和年增长量均在 1 0 %以上。
而一些新型多糖年增长量在3 0 %以上。
到目前为止,已大量投产的微生物胞外多糖主要有黄原胶(Xanthangum)、结冷胶(Gellangum)、小核菌葡聚糖(Scleeroglucan)、短梗霉多糖(Pullulan)、热凝多糖(Curdlan)等。
微生物多糖具有植物多糖不具备的优良性质,它们生产周期短,不受季节、地域和病虫害条件限制,具有较强的市场竞争力和广阔的发展前景。
目前,许多微生物多糖已作为胶凝剂、成膜剂、保鲜剂、乳化剂等。
广泛应用于食品、制药、石油、化工等多个领域。
据估计,全世界微生物多糖年加工业产值可达50~1 0 0亿美元(魏培莲,2002)。
近年来,人们越来越关注乳酸菌产生的胞外多糖,因为乳酸菌是公认的有益菌,所以它们产生的胞外多糖也具有GRAS(普遍认为是安全的)地位(De Vuysthe 和Degeest 1999)。
用能产生胞外多糖的乳酸菌发酵乳,即使只产生少量的胞外多糖(60-400mg/L),也可使发酵物具有粘稠的质地(Cerning 1990)。
此外,胞外多糖还可以改善发酵乳的质地,使其质地象奶油一样光滑细腻(Macura 和Townsley 1984),并且还能减少酸奶的乳清析出和胶体断裂,从而成为酸奶稳定剂的替代品。
一些报道还称乳酸菌胞外多糖具有优良的生物学特性,如免疫活性(Oda et al. 1984)。
综上所述,乳酸菌胞外多糖具有很广阔的发展前景。