第3章 原核生物转录及调控
- 格式:ppt
- 大小:677.00 KB
- 文档页数:25

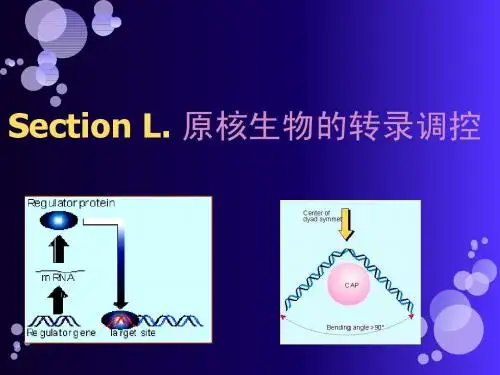
(武汉大学)分子18.※原核生物的转录调控●本章概要●不同基因启动子的强度不同,基因表达水平不同●受调控的基因(管家基因与组织特异性基因)●持家基因所有细胞中均要表达的一类基因●控制细胞基本生命过程●可诱导基因当被诱导物或细胞因子激活时表达●常常由胞外信号的改变而表达●基因调控在转录起始时最为常见●转录调控的原理●调控蛋白DNA结合蛋白,识别受其控制的基因上或基因附近的特异位点●activator 活化子(正调控蛋白)增强基因转录●repressor 抑制子(负调控蛋白)降低或关闭基因转录●一般在转录起始作用的原因●在起始阶段抑制能够避免能量和资源的浪费●比起翻译起始(调控数个mRNA),转录起始(调控一个启动子)更容易调控●在转录起始之外阶段调控的优势●可以有更多信号参与调控,具有更多检查点●对信号有更快的反应●recruitment 募集调控靶点——RNA聚合酶与启动子结合●basal level 本底水平表达组成型低水平表达●通常情况下RNA聚合酶微弱地结合在启动子上●activator 活化子同时结合DNA和RNA聚合酶,增加两者间的亲和力●有两个结合位点:DNA和RNA聚合酶●增加转录水平●repressor 抑制子结合到DNA与RNA聚合酶结合区相重叠的位点●阻止RNA聚合酶的结合,从而抑制或阻止转录●operator 操作子(注意区别operon 操纵子)DNA上抑制子结合的位点●allostery 变构调控靶点——从closed complex向open complex的转变●activator 活化子closed complex: 聚合酶与启动子结合,DNA保持双链状态 open complex: 在起始位点周围约14bp处解链形成转录泡●使RNA聚合酶从closed complex转变为open complex,继而开始转录●repressor 抑制子●抑制闭合式到开放式复合物的转变●抑制启动子逃离●远程激活和DNA环化●DNA结合蛋白互作调控蛋白可以结合在离启动子区较远的位点并发挥作用●DNA-弯曲蛋白协助弯曲DNA●转录起始外的调控●其他调控点●转录的延伸和终止●核糖体蛋白基因的翻译●由RNA进行的转录和翻译的调控●attenuation 衰减作用●核糖开关●小干扰RNA●原核生物转录调控实例●乳糖操纵子 (lac operon) [recruitment 募集调控]细菌利用乳糖作为碳源时需要lacZYA编码的酶●启动子结构组成●3个结构基因受一个启动子P_{lac}调控,转录为一条lacZYA mRNA●lacZ●编码β-半乳糖苷酶将乳糖切割成半乳糖和葡萄糖●lacY●编码乳糖渗透酶插入细胞膜,将乳糖转运到细胞内●lacA●编码硫代半乳糖苷乙酰转移酶消除lacY转入的硫代半乳糖,防止毒伤细胞●2个控制元件●CAP site●结合活化子CAP (cAMP receptor protein, CRP)●招募RNA聚合酶指导转录起始●启动子P_{lac}的-35区缺乏UP-element,不适合RNA聚合酶结合暗示了活化子结合位点的存在●O_{lac} (lac operator)●结合lac 抑制子●抑制RNA聚合酶的结合●O_{lac}与P_{lac}有重叠,阻止RNA聚合酶与启动子结合●发生机制CAP和抑制子的联合作用保证在乳糖存在且缺少葡萄糖的环境中,lac基因得以显著表达●同时存在葡萄糖和乳糖时,优先使用葡萄糖,lac基因低水平转录●抑制子感应乳糖信号●当乳糖含量低时,抑制子激活,lac基因不转录●活化子CAP感应葡萄糖信号与RNA聚合酶的αCTD直接相互作用(αCTD:原结合UP-element的位点)|帮助RNA聚合酶与P_{lac}结合和稳定●当葡萄糖含量低时,活化子激活,lac基因高水平转录●信号控制二级信号:异乳糖 / cAMP●异乳糖← lactose乳糖由β-半乳糖苷酶介导转变为异乳糖,能关闭抑制子,抑制其与O_{lac}的结合●乳糖水平低,异乳糖水平低,大部分lacZYA被抑制子抑制●乳糖水平高,(细胞内原有的)乳糖渗透酶吸收乳糖转变为异乳糖,与抑制子结合使其失活,lacZYA得到转录●cAMP← glucose葡萄糖降低cAMP浓度,抑制CAP-cAMP与CAP site的结合●葡萄糖水平低,cAMP浓度高,CAP-cAMP可以与lacZYA结合并转录●葡萄糖水平高,cAMP浓度低,CAP缺少cAMP而无法与lacZYA结合●不同的σ factor不同的σ因子结合RNA聚合酶,与不同的启动子亲和力增强,从而转录不同的启动子●σ^{70}——最常见的σ factor●热休克σ^{32}(heat shock 热休克反应)●当环境温度>37℃,参与转录蛋白质●有其特有的启动子共有序列●感染B. subtilis的噬菌体SPO1的级联表达上一时期基因的转录产物中包含这一时期所需的σ factor●两个活化子NtrC和MerR[allostery 变构调控]●NtrC——参与控制氮(N)代谢的基因表达●具有ATPase活性为诱导聚合酶产生构象变化而提供能量●作用于离基因较远处●具有独立的激活域和DNA结合域,只有在低氮时与DNA结合,使RNA聚合酶由closed complex向open complex转变低氮水平(信号)→ NtrC磷酸化发生构象改变→ 暴露NtrC的DNA结合区,发生结合→ NtrC与σ^{54}直接相互作用→ 激活NtrC的ATPase活性,为RNA聚合酶构象改变提供能量→ 转录起始●MerR——控制merT基因编码抗汞(Hg)酶●通过改变DNA构象激活merT的表达●Hg离子不存在MerR结合启动子,但其构象不利于转录(-35元件与-10元件间的距离是19bp)●Hg离子存在MerR-Hg^{2+}构象改变→ 启动子中心扭曲→ -35与-10 元件间距缩短(易于σ^{70}结合)→ 转录起始●抑制作用的其他变构机制●E.coli Gal抑制子抑制closed complex向open complex的转变●噬菌体Φ29的P_4蛋白抑制启动子的逃离(在不同启动子中作用不同)●结合强启动子P_{A2c}——抑制聚合酶和启动子之间的强相互作用加上活化子相互作用所提供的总结合能太强,将聚合酶固定在启动子上无法移动●结合弱启动子P_{A3}——激活活化子的额外结合能协助招募聚合酶●阿拉伯糖操纵子 (araBAD operon)●启动子在有阿拉伯糖而没有葡萄糖时启动激活转录,编码代谢阿拉伯糖所需的酶(类似于lac operon)●两个活化子●AraC●有阿拉伯糖时阿拉伯糖结合AraC,AraC结合araI_1和araI_2位点,I_2位点上的AraC激活RNA聚合酶起始转录●无阿拉伯糖时AraC结合araI_1和araO_2位点而发生DNA环化,araI_2位点未被激活,转录不能起始●CAP在无葡萄糖时结合到CAP site协助转录●阿拉伯糖转录启动子常被用作expression vector 表达载体●阿拉伯糖诱导araBAD启动子的数量级非常大●阿拉伯糖对araBAD启动子的控制力很强●λ噬菌体的调控●基因组结构●50Kb,含有61个基因●启动子●强启动子:P_R, P_L(P_R:向右转录,P_L:向左转录)●弱启动子:P_{RM} (Repressor Maintance), P_{RE} (Repressor Establishment)●两种生长模式●lytic growth 裂解生长●噬菌体DNA的复制●新外壳蛋白的合成●通过宿主细胞裂解释放●lysogenic growth 溶原生长●噬菌体DNA整合进入细菌染色体●在每次细胞分裂的时候主动复制●非常稳定●lysogenic induction 溶原性诱导——从溶原生长到裂解生长的转变●生长模式的维持机制由不同的启动子和调控蛋白决定●cro(control of repressor and other thing)(启动裂解生长)抑制转录●由P_R起始转录单个Cro二聚体结合O_{R3}(占据了弱启动子P_{RM}的位点)→ 强启动子P_R自身进行Cro的转录→ 不断产生Cro蛋白强化过程进行●cⅠ→λ repressor(启动溶原生长)既可以激活也可以抑制转录●正自我调控●由P_{RM}起始转录cⅠ所编码产生的λ repressor二聚化后于O_{R1}结合→ 四聚化使之与O_{R2}结合(占据了强启动子P_R的位点)→ O_{R2}上的λ repressor作为活化子招募RNA聚合酶→ 激活P_{RM}的转录进而转录cⅠ不断产生λ repressor ●生长模式的建立——cⅡ蛋白●裂解生长——通过P_R启动●P_R是强启动子,一旦感染发生,它就启动转录,产生cro蛋白●cro蛋白水平达到一定量时结合O_{R3},关闭P_{RM}(导致溶原生长无法进行)●溶原生长——通过P_{RM}启动为建立溶原生长,cⅡ 必须在cro抑制P_{RM}前激活之由P_{RE}建立,由P_{RM}维持●P_R转录的产物除了cro外还有cⅡ●cⅡ结合DNA,激活启动子P_{RE},产生cⅠ蛋白●当cⅠ达到一定量时结合在O_{R1}和O_{R2}上,启动P_{RM}的转录●cⅡ 稳定性受E. coli生长条件控制——FtsH 蛋白cⅡ在E. coli中不稳定,被蛋白酶FtsH降解●E. coli生长条件良好●FtsH活性高→ cⅡ被其降解→ 发生裂解生长●E. coli生长条件恶劣●FtsH活性低→ cⅡ积累→ 发生溶原生长●本章名词●转录调控的原理●activator●repressor●basal level (contitutive expression)●operator●recruitment●allostery●原核生物转录调控实例●Lac repressor●lac operator●lactose●glucose●expression vector●λ噬菌体的调控●lytically●lysogenically●lysogenic induction●重点知识点●什么是调控蛋白?什么是受调控基因?转录调控的原理是什么?调控蛋白可分为两类:正调控蛋白或活化子、负调控蛋白或抑制子。

原核生物基因的转录和翻译调控的细节探究在生物学研究中,原核生物的基因转录和翻译调控一直是一个热门话题。
原核生物的基因转录和翻译调控涉及的分子机制非常复杂,从转录和翻译的起始和终止位置到转录和翻译的调控因素都是需要我们深入探究的。
本文将通过细节探究原核生物基因的转录和翻译调控。
1. 转录调控的细节探究原核生物的转录调控是指在基因转录过程中,通过多种分子机制影响RNA聚合酶在DNA上的结合和进一步转录过程的调控。
在转录调控过程中,最常见的调控方式包括转录因子和各种转录调控元件的作用。
转录因子是一种特殊类型的调节蛋白,它们可以结合到DNA上的转录调控元件上,从而直接或间接地控制RNA聚合酶的下降。
在转录因子的分类中,存在两种不同类型的因子:激活因子和抑制因子。
激活因子主要通过与RNA聚合酶的结合及与与RNA聚合酶结合的DNA序列互作,来增加转录水平;相反,抑制因子则抑制RNA聚合酶与DNA序列的结合,从而降低转录水平。
除了转录因子,各种各样的转录调控元件也直接影响RNA聚合酶的结合和转录。
转录调控元件在DNA序列中特定位置上存在,它们包括操作子、增强子及隐汉子等不同类型。
操作子在DNA序列中的位置是在与RNA聚合酶结合点相对的位置上,它们通常是在编码DNA范围内的一个小的DNA片段。
增强子通常远离操作子位置,但对于RNA聚合酶的结合和转录也有影响。
最后是隐汉子,它们是特定长度的DNA序列,它们不能直接影响RNA聚合酶的结合和转录,但可以通过一些其他机制来影响整个转录大过程。
2. 翻译调控的细节探究原核生物的翻译是在转录期间直接进行的,这是因为原核生物没有核内RNA 加工的机制,而mRNA的转录过程与翻译过程之间是重叠的。
在原核生物翻译调控过程中,翻译调控元件(诸如启动子、终止子等)和核苷酸序列上的一些结构也起到了重要作用。
在原核生物的细胞质中,环状RNA(环状RNA,环C RNA)是一种能够一度大量影响原核生物翻译过程的物质。

原核生物的基因转录与翻译调控机制原核生物是指生物体细胞中缺乏真核细胞核以及其它特有的细胞器的一类生物,象细菌、放线菌等都属于这类生物。
在原核生物营养代谢产物再生的反应过程中,基因转录和翻译机制起着关键性的作用。
虽然原核生物在结构和功能上与真核生物存在差异,但其基因转录和翻译调控机制的探究仍然是一个备受关注和重要的研究领域。
一、原核基因转录的发生原核基因转录由细菌RNA聚合酶完成,RNA聚合酶由细菌DNA依据序列特异性与序列不一致性识别,准确并高效地转录所需RNA。
细菌基因起始位点(promoter)的序列组合不尽相同,其中最重要的是绝大多数细菌由主要调控元件σ—70识别维持其顺畅的生理需求,这种元件由基因-35与-10区域和过渡段区也就是-10-35之间区域识别决定。
并由帮助透过无DNA序列特异性转录激活蛋白(transcription activator)的序列(最多存在于4簇)进行辅助。
不过也有一定数量的细菌被由基因σ—54识别的类R元件助其转录,该元件由基因1-2个高度保守的区域以及管充区(38-40bp)组成。
二、原核基因转录的调控原核生物的基因在转录时可受到大量调解,使基因转录速率可被生物正式需求所有效调整。
最常见的反馈控制是会抑制在产物中需要的一个或几个酶的基因转录,这是通过由这些酶产物反馈调控于RNA聚合酶进行的。
在此基础上,产物的积累往往会对酶的活性、协同性以及数量等进行影响调解,以实现生物正式需求。
此外,细菌一般存在一套广泛而复杂的信号传导体系,称为二次信使系统(secondary messenger system),该系统是在感知环境等信号后透过一连串的化学反应激活底物转移酶(substrate transferase)透过底物磷酸化等化学改变对转录起调节作用,通常还包括一套由环状形体标记产生的传导导致产物合成的启动通路(quorumsensing pathway,简称QS),可能是底物释放或者捕获的Q信号分子,能在大多数情况下发挥合成起始因子(activator)和抑制蛋白(repressor)的作用。


原核生物基因转录调控机制的研究随着生物化学及分子生物学的不断发展,人们对生命的理解更加深入,其过程中研究原核生物基因转录调控机制不断深入,本文将对原核生物基因转录调控机制做一简单的介绍。
1. 原核生物基因转录在原核生物中,基因转录的过程通常发生在单一的细胞内,是一种高度协调的过程。
基因转录是DNA到RNA的信息传递过程,需要发生在特定的DNA区域上,该区域被称为启动子。
启动子的序列决定了哪些转录因子可以结合,从而决定哪些基因会被转录。
酵母菌的启动子大多是由一个核心启动子和多个活性装置组成,核心启动子上存在有一个区域包含与RNA聚合酶结合的TATA盒。
活性装置是指由不同的转录调控因子组合而成的调节模块。
在一些原核生物中,还有依赖微小RNA (miRNA) 和 RNA干扰 (RNAi) 的调节。
2. 原核生物基因转录调控机制原核生物通过改变转录因子的结合,加入或减少调节模块从而调节基因的表达。
转录因子是通过与启动子上相应的序列结合来激活或抑制基因转录。
在一些原核生物中,有一些具有负反馈调节的自稳定回路,这种回路位置通常在一些关键基因的下游,通过影响关键组分的表达及活动,维持系统的稳定性。
尽管转录因子是通过与启动子上的特定序列结合来调节基因转录,但原核中有一些特定序列同时可以被单个外来分子或其他细胞内分子结合。
这种情况下,转录因子与这些分子间的相互交织可能具有决定性的意义,可以大幅影响合成转录因子的能力、转录速度及准确性。
3. 原核生物是研究转录因子、mRNA的初步阶段,它们的简化数量谱系使人们能够更容易地研究它们的转录过程。
原核生物中经常被使用的实验室菌株是大肠杆菌,在它上面研究库的演化携带了丰富的基本性质关于基因转录调控的知识。
分子选择进化(SELEX)是一种常见的实验技术,用于筛选出匹配具体转录因子序列的短链DNA分子。
通过SELEX技术,人们已经挖掘了丰富的转录因子的基本性质,这一技术在原核生物中的使用更加流行。





原核生物的转录及调控课件 (一)原核生物是一类简单的单细胞生物,其转录和调控机制相对于真核生物来说要简单许多。
本文将会从原核生物转录和调控的基本知识入手,分别对其进行相关的讲解。
一、原核生物的转录机制原核生物的转录和真核生物相比较于简单,它的基因位置并没有核膜的保护,直接暴露在细胞浆中,基因间没有间隔,这意味着原核生物较容易完成基因转录任务并且速度快。
原核生物的转录分为三个阶段:启动、延伸和终止。
启动阶段是通过RNA聚合酶(enzyme)与启动子(promoter)的结合来完成的。
当RNA聚合酶与启动子结合后,它会在SO基因(即甲基接收酶基因)上找到对应的开始密码子(subunit)。
发生开放读取框架(ORF),这时RNA聚合酶就能够开始向下一个框架(ORF)复制。
与真核生物不同的是,原核生物只有一个RNA聚合酶,大部分基因的转录都是由这种聚合酶完成的。
二、原核生物的调控机制原核生物的基因调控是非常重要的一部分。
它们使用许多方法来对其基因表达进行调控,以适应环境变化和其它外部信号的影响。
原核生物的基因调控主要分为两种方式:正调控和负调控。
1. 正调控:这种方式是使得信号物质(Messenger)能够结合到转录因子上,进而使其能够附着到启动子(promoter)上面。
这些信号物质可以直接或间接地影响基因的表达,以达到对细胞的调节效果。
在真核生物中,常见的正调控因子有启动子结合蛋白(TBP)。
2. 负调控:在负调控中,信号物质通过阻止转录因子的结合,来预防其结合到启动子上来。
这将使得该基因的表达能够被阻止,因此,其效果可能与正调控相反。
在原核生物中这种现象是非常常见的。
总结在原核生物中,转录和调控的机制相对比较简单。
它们基因转录的速度较快,在基因调控时使用正调控和负调控来达到有目的地外部信号的调节效果。
虽然其调控机制相对单一,但是作为生物学的基础研究,其在基因转录和调控方面的应用可能带来重要突破。