突触可塑性
- 格式:doc
- 大小:48.00 KB
- 文档页数:6

突触可塑性的生物物理机制与应用研究第一章突触可塑性的概念和分类突触是神经元之间传递信息的重要结构,突触可塑性是指突触自身或连接其他神经元的能力进行学习和记忆的过程。
突触可塑性的种类有很多种,本章将介绍其中的主要种类。
1.1 短期可塑性短期可塑性是指突触在一定时间内会发生改变,改变后可以恢复到之前的状态。
这种可塑性分为两种:早期和晚期。
早期短期可塑性通常在数秒到数分钟之内发生。
这种可塑性通常由神经递质的释放和突触后响应的改变引起。
例如,在兴奋性突触中,重复刺激会导致短暂的增强(俗称为"后向增强"),这种增强可以持续几秒钟,但不会导致长期更改。
晚期短期可塑性发生在几分钟到几小时之间。
这种可塑性是由于神经递质、受体密度、离子通道和突触传递的信号转导途径发生突触后效应的改变。
例如,在镁离子的存在下,高频刺激可以导致长时间的增强,这种增强可以持续几十分钟,但不会导致永久性的更改。
1.2 长期可塑性长期可塑性是指突触发生持久性改变,可能会持续数小时到数十年之久。
长期可塑性通常发生在大量的突触活动或体验后。
这种可塑性可分为两种主要类型:增强和抑制。
增强性长期可塑性发生在强烈的突触活动后,通常会增加神经递质的释放和突触传递的信号转导途径,以及增加附着于神经递质的受体密度。
增强性长期可塑性通常与学习和记忆有关。
抑制性长期可塑性通常发生在经验量的低活性后,通常会导致神经递质释放和突触传递的信号转导途径的下降。
抑制性长期可塑性通常与逆向学习和遗忘有关。
第二章突触可塑性的实验技术了解突触可塑性的机制需要使用一些实验技术。
下面介绍几种常用的实验技术。
2.1 突触切片实验突触切片实验可以对突触进行操作,提供用于观察其可塑性的条件。
突触切片实验通常是从大脑中取出一部分组织,在显微镜下进行操作,如切片、刺激和膜片钳技术。
这些切片可以用来研究突触前后细胞间连接及其对生理学活动的作用。
2.2 活体实验活体实验是在动物的大脑中进行的。

记忆形成过程中的突触可塑性机制记忆是人类思维和认知能力的核心之一,它使我们能够回忆过去的经历、获取新的知识和技能。
记忆的形成是一个复杂而精确的过程,其中突触可塑性机制发挥着重要作用。
本文将探讨记忆形成过程中突触可塑性的机制,帮助我们更好地理解记忆的神秘世界。
突触可塑性是指突触连接的强度可以改变的一种现象。
突触是神经元之间传递信息的联系点,在控制信息传递过程中起着关键作用。
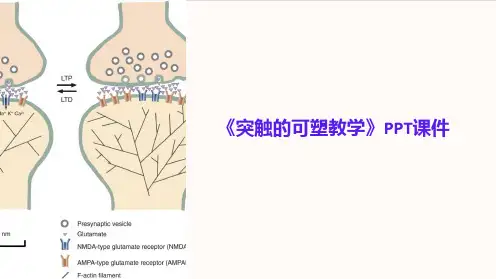
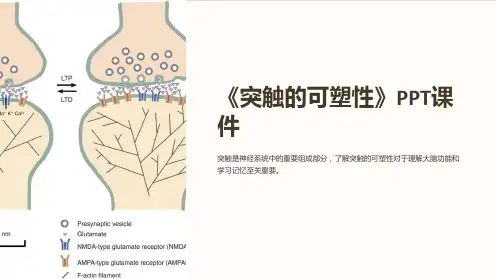
突触可塑性具有两种主要形式:长期增强(long-term potentiation,简称LTP)和长期抑制(long-term depression,简称LTD)。
LTP是指当突触反复受到刺激时,它的传递效率会增加,即突触连接的强度增强。
LTP通常包括高频刺激引起的短时LTP和持久LTP。
短时LTP主要通过改变突触前神经元释放神经递质的机制来实现,而持久LTP则涉及到突触后神经元中多种细胞内信号通路的激活。
LTP的发生依赖于突触前和突触后神经元之间的相互作用,包括神经递质的释放、突触后细胞内钙离子浓度的升高以及细胞内信号通路的激活。
LTD与LTP相反,是指当突触反复受到低频刺激时,突触传递效率会减弱,即突触连接的强度降低。
LTD的发生依赖于突触前和突触后神经元之间的相互作用,包括神经递质的释放调节、突触后细胞内钙离子浓度的变化以及细胞内信号通路的抑制。
LTP和LTD的相互作用是记忆形成过程中突触可塑性的基础。
LTP的产生可以加强突触的连接,增强特定的神经回路,有助于记忆的保持和巩固。
而LTD的发生则可以减弱某些突触的连接,削弱不必要的信息传递,从而提高记忆的选择性和灵活性。
突触可塑性的机制与神经递质、细胞内信号通路和突触结构的变化密切相关。
神经递质在突触传递过程中扮演着重要角色,它们通过突触前神经元释放,与突触后神经元上的受体结合,进而影响突触连接的强度。
细胞内信号通路则通过激活蛋白激酶和磷酸酶等酶的活性,调节突触连接的强度和稳定性。


神经科学中的突触可塑性及其意义神经科学是研究神经系统功能和结构的学科,突触可塑性则是神经科学中十分重要的研究领域。
突触可塑性是指突触连接的强度和性质随神经系统中的活动而变化的现象,被认为是神经信息传递和学习记忆形成的基础。
本文将阐述突触可塑性的特点、机制以及在神经系统功能、疾病和治疗方面的意义。
一、突触可塑性的特点突触可塑性是指突触连接的强度和性质会随着神经系统中的活动而变化。
这种变化可以持续很长时间,甚至是永久的。
其模式包括增强和抑制。
增强作用是指神经元之间的连接强化,从而使神经元被更容易激活。
抑制作用是指神经元之间的连接变弱,从而使其在之后更难以激活。
突触可塑性是一个高效的神经系统信息传递调节机制,在生物进化过程中具有重要的优势。
例如,大脑可以通过突触可塑性在很短的时间内改变神经元之间的通讯方式,从而对环境做出快速反应,这也是动物在生存和繁衍中不可或缺的生物学基础。
二、突触可塑性的机制突触可塑性的机制主要包括前突触和后突触两个方面。
前突触是指神经元释放到突触前端的神经递质的机制。
细胞内的电流脉冲会引发电压门控的钙通道开放,从而使细胞内的钙离子浓度升高,钙离子能够触发突触前端的囊泡释放神经递质。
神经递质在突触间隙内形成信号,激活相关的拮抗或增强通路。
后突触是指突触后面的神经元为了响应前突触释放的神经递质而释放的信号。
后突触释放的信号主要由第二信使和突触后细胞膜的离子通道控制。
神经递质的作用,通过与神经元中的受体结合而产生影响。
这些受体随时可以被激活或被抑制,从而通过调节神经递质在突触间隙内的浓度和后突触神经元的反应而产生前突触的调节。
三、突触可塑性在神经系统功能、疾病和治疗方面的意义突触可塑性在神经系统功能、疾病和治疗方面的意义具有十分重要的影响。
1. 神经系统功能的意义突触可塑性是神经系统最基本的适应性和学习能力的基础。
在大脑中,突触可塑性是记忆形成和学习的基础。
人们学习新知识时,原有的突触被加强或削弱,从而有效地存储和检索信息。

神经元可塑性的秘密神经元是构成神经系统的基本单位。
它们以极其复杂的方式相互连接,通过电信号进行信息传递。
然而,神经元并不是静止的实体,而是具有可塑性——即能够适应外界环境并改变其功能和结构的能力。
神经元可塑性的秘密一直是神经科学领域的重要研究方向之一。
可塑性的类型神经元的可塑性可以分为两大类:突触可塑性和细胞可塑性。
突触可塑性突触可塑性指的是突触前后元件之间连接强度发生改变的现象。
具体而言,突触可塑性包括增强突触传输和减弱突触传输两种形式。
增强突触传输(突触增强):当一个突触被高频激活时,突触传输效率会增加。
这种增强可以持续一段时间,并且可以在短时间内产生更大的电流响应。
减弱突触传输(突触抑制):与增强相反,当一个突触被低频激活或者一段时间内没有激活时,突触传输效率会减小。
细胞可塑性细胞可塑性指的是神经元自身特性发生改变的能力。
它涉及到细胞的发育、表达基因、形态结构等方面。
细胞发育:在神经系统发育过程中,神经元会经历不同的阶段。
例如,在婴儿期和青少年期,大脑将会进行快速的发育和重组,以适应新环境。
突触重建:当神经元受到损伤或环境改变时,它们可以通过重新组织、重建或产生新的连接来适应新情况。
表达基因:研究表明,外界刺激可以影响基因表达,并导致神经元特定蛋白质表达量的变化。
这些蛋白质对于调节突触连接和细胞功能至关重要。
神经元可塑性的机制神经元可塑性的机制非常复杂,目前还没有完全解析清楚。
然而,已有许多研究揭示了其中一些关键机制。
突触可塑性机制长时程增益(LTP):LTP是最著名和最广泛研究的突触可塑性机制之一。
当一个突触接收到高频激活时,钙离子通道会打开并导致二级信号路径的改变,进而引起强化效应。
长时程抑制(LTD):与LTP相对应,当一个突触接收到低频激活或一段时间内没有激活时,LTD被诱导并导致突触传输效率降低。
突触结构重构:部分研究表明,在突触可塑性发生时,部分突触可能会增加或减少其数目或大小,并且可以更改有关神经信号传播的部分结构。

突触可塑性的名词解释突触可塑性:是指突触前神经元释放的多巴胺或乙酰胆碱与突触后膜上的一种特异性的神经递质结合而引起的突触后电位,改变了传入冲动的形式和传播速度,进而影响学习和记忆的过程。
上皮神经元的递质是乙酰胆碱和多巴胺,轴突末梢的递质是乙酰胆碱和去甲肾上腺素。
突触传递作用主要是通过改变突触后膜上受体和离子通道对递质的敏感性而实现的,这些受体主要分布在突触后膜和胞浆中,包括许多与递质浓度和离子流动密切相关的离子通道,其中主要包括: h。
但并非全部的突触小泡都能脱颗粒,因为突触后膜上还存在着抑制递质的能力,叫做突触后膜的“门控”系统,其中最主要的是胆碱能受体,它们对于低级胆碱的摄取具有高度的选择性。
另外,突触后膜上还有许多对多巴胺具有高度亲和力的受体,它们与递质结合后可以直接引起膜对na的通透性增加。
由此可见,虽然突触后膜上没有“门”的概念,但当兴奋到达时,可导致整个细胞内ca2内流而致突触后膜对na的通透性增强。
根据突触形成的原理,突触传递可以被归纳为以下几个步骤: 1、递质与突触前膜上特异的受体相结合,引起膜对递质的通透性增强;2、通透性增强使离子内流,而后将突触后膜上的受体与膜上的特异性离子通道结合;3、离子通道将改变膜电位,从而激活下游的钙离子通道和钾离子通道,使后者打开;4、上述的突触后膜改变最终导致细胞外液中的钙离子大量内流,使突触后膜去极化;5、最终是细胞内钙离子大量内流导致动作电位的产生。
突触传递就是以上五个过程的循环反应。
2、抑制,是指细胞[gPARAGRAPH3]会因各种原因引起,一部分对特定离子或递质敏感的突触前膜上的受体的数目减少或功能减弱,从而引起递质与突触后膜上的受体结合减少或消失,甚至完全不能结合,这种状态称为突触的抑制。
比较常见的例子如下:(1)氨基酸类递质抑制( GABAA)当突触前膜上含有代谢性递质(谷氨酸等)时,突触后膜的运动性神经元中的GAM受体数目会减少,突触后膜对递质的敏感性降低,从而使突触后膜的电位改变,最终使突触后膜兴奋性降低,呈现不易兴奋状态,称为突触后膜的抑制。

神经科学中的突触可塑性研究神经科学是研究生物神经系统如何控制行为的科学。
突触是神经元之间连接的关键点,它是神经系统功能的基本单位。
突触可塑性研究是神经科学的一个重要分支,它探究了神经突触连接随时间和环境变化的能力。
突触可塑性的定义突触可塑性是指突触神经元之间的连接强度可以随时间和环境变化的能力。
这意味着突触可以通过一些刺激,在长时间的过程中来改变其传递信息的强度。
例如,长期强化(LTP)和长期抑制(LTD)就是突触可塑性的两种形式。
LTP和LTD简介LTP和LTD是突触可塑性的两种形式。
LTP会增强突触中的某种神经递质的传递效果,这样使得神经元之间的连接更强。
LTD 则会削弱突触中的某种神经递质的传递效果,这样使得神经元之间的连接更弱。
这种调整可以帮助神经元之间更加精细地互动,以达到正确的响应。
突触可塑性的意义突触可塑性的意义在于神经元能够通过学习和记忆来改变连接的强度。
例如,当我们学习一个新技能时,我们的神经元之间的连接可能会改变,这样我们就可以更好地完成任务。
同样的,当我们忘记一件事情时,我们神经元之间的连接也可能会改变,这样我们就可以更加容易地忘记这个记忆。
神经可塑性与神经疾病突触可塑性的研究对于神经疾病的治疗也有重要的意义。
例如,阿尔茨海默病(AD)就是突触可塑性受到损害的疾病。
研究人员通过研究病理性LTP和LTD的过程,已经开始着手研发用于治疗AD的新药物。
这些药物可能会改变神经可塑性的动力学或改善病人的神经可塑性,从而帮助病人恢复正常的神经功能。
突触可塑性研究的进展随着突触可塑性研究的发展,研究人员已经提出了许多对于突触可塑性的机制的分析方法。
其中一种方法是神经元的行为模拟。
通过模拟神经元的活动,研究人员可以更好地了解突触可塑性的机制,包括LTP、LTD和突触清理。
我们现在已经可以对神经系统的运作机制以及神经系统对人类行为的控制方式有更深入的了解。
随着未来神经科学的不断发展,我们可以期待更多突触可塑性相关的发现的出现,这些发现将推动我们更深入地了解大脑,并开发出更好的神经疾病治疗方法。

神经科学研究中的突触可塑性现象突触可塑性是指神经元之间的连接在学习和记忆过程中发生变化的现象。
这种现象是神经科学研究中的重要课题,对于我们理解大脑是如何存储和加工信息至关重要。
突触可塑性主要包括长时程增强(LTP)和长时程抑制(LTD)两种形式,这些变化可以持续从几分钟到几年不等。
突触可塑性的发现是在20世纪60年代由神经科学家Tim Bliss和Terje Lømo通过对海兔的突触进行电生理实验而得出的。
他们发现,通过高频刺激突触,可以使突触传递的电信号增强,这种增强可以持续数小时到数天。
这个发现开创了突触可塑性研究的先河,引发了全球范围内的科学家对于这一现象的关注和研究。
在突触可塑性的研究中,神经科学家们探索了多种机制和分子信号调节突触可塑性的过程。
其中一个重要的机制是NMDA受体介导的钙离子内流。
NMDA受体是一种离子通道,当发生突触传递时,需要同时存在刺激性神经递质的释放以及突触膜上的去极化,才能使NMDA受体打开。
这时,钙离子会进入突触细胞,激活一系列的信号转导通路,导致突触可塑性的发生。
此外,神经递质的释放和突触水平的可塑性也密切相关。
典型的神经递质包括谷氨酸、γ-氨基丁酸(GABA)和乙酰胆碱等。
这些神经递质的释放受到突触前和突触后神经元的相互作用和调节。
当兴奋性神经递质释放增加时,突触可塑性往往会增强,而当抑制性神经递质释放增加时,突触可塑性往往会减弱。
这种神经递质调节的平衡关系对于神经元网络的正常功能非常重要。
突触可塑性的研究不仅帮助我们理解学习和记忆的机制,也有助于揭示许多神经系统疾病的发生机制。
例如,突触可塑性的异常可能与阿尔茨海默病、帕金森病以及自闭症等疾病的发生和发展有关。
通过研究突触可塑性,我们可以寻找改善这些疾病的治疗方法,为神经系统疾病的治疗和预防提供新的思路和策略。
近年来,随着神经科学研究的不断深入,突触可塑性以及与之相关的机制和分子通路得到了更加详细和广泛的研究。

神经科学中的突触可塑性是一个十分重要的话题。
突触是神经元之间传递信息的地方,而突触可塑性则是指突触连接强度可以改变的能力,这对于学习记忆、行为调节等神经系统功能都至关重要。
接下来,让我们一起深入了解。
一、突触的类型突触分为化学突触和电子突触两种类型。
电子突触是指两个神经元之间通过直接联系的方式传递信息,而化学突触是指神经元之间通过释放化学信号物质来传递信息。
化学突触是人们所熟知的、常见的一种突触类型。
神经元通过突触前端分泌神经递质,被称为神经元前体,神经递质跨越突触裂隙,到达突触后膜上的神经元接受器。
然而,突触前体和接收器之间的距离少于20nm,这种短距离连接似乎不能通过扩散来实现,因此需要一段特殊的结构来实现化学信号传递。
这个特殊结构被称为活动区域。
二、突触可塑性神经系统的可塑性是指神经元及其突触的改变能力。
在学习和记忆过程中,人们的神经元和突触会发生可塑性变化,这就是突触可塑性。
突触可塑性的形成是通过调节突触前体释放的神经递质量和突触表面的受体数量来实现的。
神经元释放的神经递质可以增加或减少另一个神经元上的接收器数量,从而导致神经元之间的相互作用强度发生变化。
可塑性的促进因素包括物理或化学刺激、增加或减少神经递质分泌、调整活动区域等。
通常,突触可塑性存在两种形式:其中一种是称为长时程增强(LTP)。
“长时程增强”指的是有一种特定的刺激可以导致神经元之间拥有持续时间更长的连接。
这种可塑性常常是由于神经元放电和突触前体释放的增加造成的,并且这种放电可以持续很长时间。
另一种形式是称为长时程抑制(LTD)。
LTD是指神经元之间的连接弱化,它可以被不同的刺激所诱导,如当前两个神经元的同时放电。
这通常由于突触前体释放量的减少和突触内部多巴胺浓度的下降所导致。
三、突触可塑性的应用突触可塑性是储存在人类大脑中的信息的基础,因此,它在人工智能领域也有着极其丰富的应用。
我们可以通过构建基于神经元和突触的人工神经网络,从而实现“学习”的功能,而这一过程中突触的可塑性起到了关键的作用。
脑神经科学中的突触可塑性突触可塑性(synaptic plasticity)是指神经元之间的联系能够因为某种形式的输入而发生变化,包括长期增强(LTP)和长期抑制(LTD)等不同的形式。
在神经科学研究上,突触可塑性是一种被广泛研究的现象,因为能够展现出神经元之间间接的联系,从而可以推断出神经元之间的联系和信息传递机制。
一、突触可塑性的发现20世纪50年代,神经科学研究人员已经开始对神经元之间的连接良好地认识,但当时他们所认识的突触行为是一种非常固定的状态,似乎没有同时被激发而产生的增加或者减少。
但是这种信念在20世纪60年代被证明是错误的,斯塔姆巴赫教授发现了雄性大鼠很强的电刺激条件下能够增强杆状神经元(英语:Pyramidal neuron)之间的突触效能,也就是后来人们所研究的长期增强(LTP)现象。
同时,加拿大乔纳斯教授也探索了LTD的现象,并最终证明了LTD和LTP共存的情况。
二、突触可塑性的意义突触可塑性对于神经科学研究的意义很大,因为他可以让我们了解神经元之间的互动方式和信息传递机制。
LTP和LTD等突触可塑性的反应可以模拟现实情境,也会因为神经元之间的反应而影响我们平日从事的各种活动以至于我们的记忆并形成了我们的知识储备。
此外,突触可塑性有助于理解神经系统在各种情境下的适应性和能力,也可以帮助我们在神经元或者人工智能的研究方面提供思路。
三、突触可塑性的研究方法研究可塑性的方法有很多种,从早起的刺激方式到最近精细的电学和化学方法。
例如,可以用高强度电脉冲进行长期增强,或反之进行长期抑制。
还有一些化学方法如轻链蛋白(经过选择后)用于抑制KIF4A,可以通过阻止KIF4A促进突触可塑性来促进突触可塑性。
同时,神经科学包括化学、生理、形态、分子及形态等多学科的技术结合也可以加速研究进展。
四、突触可塑性的应用突触可塑性的研究也有一些具体的应用。
例如,根据神经元之间的联系变化可以推断出某些神经疾病的情况以及导致这些疾病的原因。
神经科学研究中的突触可塑性神经科学研究中的突触可塑性是指神经元之间连接的突触结构和功能在不同的刺激和环境下可以发生改变的过程。
突触可塑性是神经系统学习和记忆形成的基础,对理解大脑的信息处理和神经疾病的发生机制具有重要意义。
突触可塑性主要分为两种形式:突触增强和突触抑制。
突触增强即突触前神经元和突触后神经元之间的连接强度增加,这种情况下,刺激强度较小的突触前神经元也能引起突触后神经元的兴奋;而突触抑制则是连接强度减弱,导致突触前神经元的刺激难以引起突触后神经元的兴奋。
突触可塑性的机制主要涉及两个方面:突触前放电的强度和频率、以及突触后放电的强度和频率。
突触前放电的强度和频率越高,突触后神经元的兴奋程度越大,从而加强突触连接;而突触后放电的强度和频率越高,突触后神经元对突触前刺激的敏感程度越高。
神经科学研究中的突触可塑性的现象主要有两种:长期增强和长期抑制。
长期增强是指突触可塑性持续较长时间,增强了突触连接的强度,从而促进了学习和记忆的形成;而长期抑制则是指突触连接的强度持续较长时间减弱,导致学习和记忆的阻碍或遗忘。
突触可塑性的调控机制涉及到多种分子、细胞和网络水平的调节因素。
其中,神经递质和神经调节因子的释放、突触前和突触后的特定受体的激活、神经元膜电位改变以及突触突触结构的变化都能影响突触可塑性的发生。
突触可塑性在大脑的学习和记忆过程中起着关键作用。
通过突触可塑性,神经元之间的连接可以根据输入的模式进行改变,形成新的神经回路,从而记忆信息可以被长期储存。
突触可塑性的异常也与多种神经系统疾病的发生和发展密切相关,如帕金森病、阿尔茨海默病和自闭症等。
研究突触可塑性的手段包括行为学、电生理学、成像技术和分子生物学等。
行为学观察动物学习和记忆过程中的行为变化,电生理学记录神经元的电活动,成像技术可以实时观察突触活动的动态变化,分子生物学可以检测特定的分子机制参与突触可塑性的调控。
总结起来,神经科学研究中的突触可塑性是指神经元之间连接的结构和功能可以根据刺激和环境发生改变的过程。
精神疾病中突触可塑性的异常研究引言精神疾病是一类严重影响人类心理和行为功能的疾病,包括抑郁症、精神分裂症、焦虑障碍等。
过去的研究发现,精神疾病与突触可塑性异常之间存在密切关系。
突触可塑性指的是神经元之间连接强度和功能在不同条件下改变的能力,它对于学习记忆、情绪调节等神经功能至关重要。
本文将从细胞水平、大脑回路以及突触信号通路三个方面探讨精神疾病中突触可塑性异常的相关研究。
细胞水平上的突触可塑性异常1. 精神疾病与长时程增益型突触可塑性之间的联系长时程增益型突触可塑性(long-term potentiation, LTP)是指在重复刺激后,突触效应强度提高并持久一段时间。
多项实验证实了LTP在学习记忆等认知功能中的重要性。
研究发现,在某些精神疾病患者的脑组织样本中,LTP过程异常,导致神经信号传递受阻。
2. NMDA受体与突触可塑性异常的关系N-甲基-D-天冬氨酸(N-methyl-D-aspartate, NMDA)受体是突触可塑性的重要调控因子。
一些精神疾病患者显示出NMDA受体异常表达或功能失调。
研究发现NMDA受体缺陷可能导致突触可塑性异常从而参与了精神疾病的发生和进展。
大脑回路上的突触可塑性异常1. 杏仁核-前额叶皮质回路和情绪障碍之间的关系杏仁核和前额叶皮质形成了一个重要的情绪调节回路。
多项临床实证和动物模型研究表明这个回路在焦虑障碍、抑郁症等精神疾病中扮演了关键角色,与它们相关的行为和情绪缺陷可能部分归因于突触可塑性异常。
2. 海马体与记忆障碍之间的关系海马体是大脑中负责记忆和学习的关键结构。
在一些精神疾病患者中,海马体突触可塑性异常导致了记忆障碍的出现。
研究发现抑郁症患者海马体突触长时程抑制性可塑性增加,而精神分裂症患者则显示出减弱的长时程增益型可塑性。
突触信号通路上的异常1. 多巴胺系统在精神疾病突触可塑性异常中的作用多巴胺是调节情绪和奖赏机制的重要神经递质。
精神疾病如精神分裂症、成瘾等与多巴胺功能失调密切相关。
神经元突触可塑性生理学神经元突触可塑性是指神经元之间的连接强度和功能可以根据输入和输出之间的关系进行改变的能力。
它是神经系统学习和记忆的基础,也是了解神经退行性疾病和神经重塑的重要窗口。
本文将探讨神经元突触可塑性的生理学机制和影响因素。
一、突触可塑性的基本概念神经元之间的连接通过突触完成传递信息的功能。
突触可塑性指的是突触连接的强度和功能可以根据输入输出之间的关系进行调整和改变。
它可以分为两种类型:增强型突触可塑性和抑制型突触可塑性。
增强型突触可塑性,又称为长时程增强(LTP),是指当一个突触反复被激活时,其传递效率会增强,即连接的强度增加。
这种可塑性可以持续很长时间,并且对于学习和记忆等认知功能非常重要。
抑制型突触可塑性,又称为长时程抑制(LTD),是指当一个突触反复被抑制时,其传递效率会减弱,即连接的强度降低。
这种可塑性对于平衡和调节神经元之间的信息传递起到重要作用。
二、突触可塑性的生理机制突触可塑性的生理机制主要涉及突触前膜和突触后膜的信号传导和调节。
1. 突触前膜的机制突触前膜主要涉及神经递质释放机制。
当神经冲动到达突触前膜时,它会导致电压依赖性钙通道的打开,进而使得钙离子流入突触前膜。
这些钙离子的流入会促使神经递质囊泡与细胞膜融合,并释放神经递质到突触间隙,从而实现神经信号的传递。
2. 突触后膜的机制突触后膜主要涉及突触后受体的活性和信号转导。
神经递质在突触间隙释放后,会与突触后受体结合,触发细胞内的信号转导通路。
这些信号转导通路可以改变突触后膜的通透性,调节突触后细胞的兴奋性和抑制性,以及调控突触通道的开放和关闭,从而影响突触的可塑性。
三、神经元突触可塑性的调控因素神经元突触可塑性受到多种因素的调控,包括突触前膜因素、突触后膜因素和环境因素等。
1. 突触前膜因素突触前膜的标记和突触前膜蛋白的调节对于突触可塑性至关重要。
例如,突触前膜标记的变化可以指示突触连接的强度和状态,从而影响突触的可塑性。
神经生物学中的突触可塑性突触可塑性是神经生物学中一个复杂而又神秘的课题,也是神经科学家们长期以来研究的重点。
突触是指神经元之间的连接点,也是信息传递的重要通道。
我们的大脑拥有数百亿个神经元,它们之间的突触连接构成了极其复杂的网络结构。
神经科学家们早就发现,突触的连接不是一成不变的,而是具有可塑性。
当我们学习新知识时,大脑中的神经元之间的突触连接和传递信息的方式会发生改变。
这种突触可塑性是神经元能够响应环境变化、适应外界要求的基础,它是大脑能够学习、记忆的重要基础。
突触可塑性的形式有很多种,以下是常见的三种形式。
1. 长时程增强(LTP)在大脑中,当同一组神经元同时被多次激活时,它们之间的突触可塑性可能会增强。
这种增强过程叫做长时程增强(LTP)。
在LTP发生之后,当这组神经元再次激活时,其产生的效应可能会变得更加强烈。
LTP被认为是学习和记忆的基础之一。
2. 长时程抑制(LTD)与LTP相反,当同一组神经元被同时激活时,它们之间的突触可塑性也可能会减弱。
这种减弱过程叫做长时程抑制(LTD)。
在LTD发生之后,当这组神经元再次激活时,其产生的效应可能会变得更加弱化。
LTD也被认为是学习和记忆的重要基础之一。
3. 短时程增强(STP)短时程增强(STP)是指突触在短时间内(几百毫秒内)响应强度的暂时提高。
这种现象通常由于神经元之间短时突触的可塑性改变所引起。
与LTP和LTD不同的是,STP只持续很短的时间,但它在多个神经元之间的快速切换和协调起到了重要的作用。
以上三种突触可塑性在大脑的不同区域,不同神经回路中都会发生。
在谷氨酸型神经元(Glutamatergic Neuron)和主要抑制性神经元(GABAergic Neuron)之间的突触连接中,LTP和LTD被认为是最常见的可塑性形式。
LTP和LTD对神经元的突触连接进行了有针对性的调整,从而使得神经回路能够适应环境变化,实现外界刺激的有序响应。
突触可塑性机制与学习记忆一、引言突触可塑性作为神经系统最基本的学习记忆机制,是指当一个突触重复受到神经冲动刺激时,其表现出来的反应会随时间的推移而改变。
这种变化既可能是突触前神经元输入的改变,也可能是突触后神经元的改变。
这种可塑性机制是神经系统不断适应环境、学习记忆的基础。
二、突触可塑性的形式1. 短程突触可塑性短程突触可塑性指的是短暂的,持续时间短的突触可塑性,它通常涉及单个突触。
主要分为突触后感受性增强(Post-Synaptic Potentiation, PSP)和突触前抑制(Pre-Synaptic Inhibition, PSI)两种。
2. 长程突触可塑性长程突触可塑性指的是较长时间的,持续时间较长的突触可塑性,和大范围的神经元群体活动相关。
它通常涉及多个突触同时发生变化。
主要包括长时程增强(Long-term potentiation, LTP)和长时程抑制(Long-term depression, LTD)两种。
三、突触可塑性的生化机制1. LTP的生化机制LTP是指当一个突触反复被激活时,其相应效应会随时间的推移而变强,这种变化能够持续一段时间。
LTP可用于解释学习和记忆的整体过程。
LTP的生化机制主要包括以下三个环节:(1)突触前膜的NMDA受体活化当来自突触前神经元的信号通过AMPA受体进入突触后神经元时,会导致附着在突触前膜上的NMDA受体的通透性发生变化,使得钙离子进入细胞内部,启动下一步的LTP机制。
(2)蛋白质合成的启动钙离子的进入在复杂的信号转导过程中启动了神经元细胞内的蛋白质合成。
这种蛋白质的合成是LTP形成的必要条件之一。
(3)新突触的形成神经元上的新突触可以通过将新合成的突触蛋白传递到突触前膜来形成。
只要这个过程连续进行,就能形成新的突触,从而使得LTP持续触发。
2. LTD的生化机制LTD指触发某些突触的活动可能会在较长时间内下降,这种下降的过程就是长时程抑制。
突触可塑性:多种形式,功能和机制(紫色代表部分重点内容)摘要:不论是来自一堂课所学,一件很有压力的事情或者吃下精神药物,这些形成的感觉都会通过修饰神经的活性和组织特殊的神经回路来影响大脑。
一次经历形成的神经活性修饰大脑功能的主要机制就是通过修饰突触的传递;也就是说:突触可塑性。
这里,我们回顾最近对哺乳动物大脑兴奋性突触的突触可塑性的主要形式的产生机制。
我们也提供那些突触可塑性造成的可能促进的和行为上的功能的例子以及海马区域可塑性是如何导致神经精神紊乱的。
关键词: 长时程增强效应,长时程抑制效应,NMDA受体,AMPA受体,海马,成瘾介绍哺乳动物大脑的一个比较重要和引人注意的特性就是可塑性:通过一次经历修饰神经回路功能形成的神经活性的容量,从而导致接下来想法,感觉和行为的改变。
突触可塑性特指预先存在的突触的突触传递的强度和效能的活性依赖性的修饰,并且一个世纪以来突触可塑性都被认为是大脑将暂时性记忆转变为永久记忆的中心角色。
同时突触可塑性也被认为在神经回路的形成中起到重要作用,并且有证据累积表明突触可塑性的损伤和几种著名的神经精神紊乱相关。
这样子理顺几种任何脑区的突触可塑性的详细分子机制对于理解正常的和病理的脑功能的许多方面的神经基础都很有意义。
了解突触可塑性的多种功能,就会发现有许多种突触可塑性的形式和机制都被描述过了。
通过活动突触传导可以被增强或者抑制,并且这些改变会有一个时间上的延续,从微秒到小时,天或者可能更长的时间。
并且,基本上所有的哺乳动物脑的兴奋性突触都同时表现出一系列的不同突触可塑性的形式。
这里,我们尽量提供一个全面的关于哺乳动物脑兴奋性突触的突触可塑性的著名形式的机制。
简要的回顾突触可塑性的短时程形式后,我们会强调一下近来对细胞机制的理解和长时程增强(LTP)与长时程抑制(LTD)的可能的功能。
短时程突触可塑性:大量形式的短时程的突触可塑性,持续时间从微秒到几分钟,基本上在所观察的所有简单非脊椎动物到哺乳动物的每个突触都发现了(Zucker and Regehr, 2002)。
短时程突触可塑性被认为在下列现象中起主要作用: 感觉输入的短时程适应,行为状态的暂时性改变,短时的记忆形式。
大多形式的短时程突触可塑性都是由短暂的活性的爆发引起,从而引起在突触前膜神经终末Ca2+的暂时性累积。
这个突触前膜Ca2+的增加会反过来导致神经递质释放可能性(p)的改变——通过直接修饰突触小泡胞吐作用的生化过程。
成对刺激易化和抑制当两个刺激在比较短的时间内产生,那么第二个刺激的应答与第一个刺激相比可以被增强或者被抑制(Katz and miledi, 1968; Zucker and Regehr, 2002). 成对刺激抑制在短刺激间隔(小于20ms)时,在所有突触都能被普遍观察到,这最有可能是因为电压依赖Na+或者Ca2+通道的失活,或者突触前终末小泡准备释放池的暂时耗竭。
许多突触在较长的刺激间隔(20-500ms)中表现出成对刺激易化。
对于这种现象的一个简单的解释就是,第一个动作电位时进入细胞内的残留的Ca2+会导致第二次刺激时小泡的额外的释放,但是似乎这里面还涉及到其他的机制。
这些可能涉及到调节突触前膜磷蛋白活性的蛋白激酶的激活。
例如,突触前膜磷蛋白突触蛋白基因被敲除的老鼠会表现出短时程突触可塑性的异常。
突触表现出成对刺激易化还是抑制取决于突触激活的最近的历史。
由于这些形式的突触可塑性很大程度上取决于递质释放可能性(p)的改变,因此以比较高的p起始的突触倾向于抑制第二个刺激的应答(Dobrunz and Stevens, 1997)。
反过来,起始p比较低的突触一般就会表现出第二个刺激应答时p的增加。
坚信这个观点,人为降低p基本上总会引起成对刺激易化的放大甚至成对刺激抑制翻转为易化。
因此,根据突触激活和修饰的最近的历史,同一个突触可以表现出易化或者抑制。
成串刺激的易化和抑制高频(10-200Hz)的成串持续性(200ms-5s)重复或者强直刺激之后可以观察到长时程突触可塑性(Zucker and Regehr, 2002)。
强化和强直后增强(PTP)就描述了递质释放时间从几秒到几分钟的增强过程。
由于刺激时突触前膜内Ca2+的增加,神经递质释放的概率p 也增加。
残留的Ca2+合并接下来单刺激内流的Ca2+直接增强了神经递质的释放,或者导致了突触前终末内蛋白质的生化修饰(Magleby and Zengel,1982; Zucker and Regehr,2002)。
在一些突触内,重复的激活可以导致持续几秒到几分钟的抑制(Betz,1970; Zucker and Regehr,2002)。
在成对脉冲抑制中,抑制一般发生在p比较高的突触中,这被人们解释为或者部分解释为:由于突触小泡准备释放池的暂时耗竭。
突触传递的效能的降低也可以由以下原因造成: 被激活的突触前终末、突触后细胞或者邻近的细胞释放的调节物质,从而导致一个抑制突触前膜释放的信号瀑布。
最后,短时程可塑性突触后膜机制可能与配体门控受体的脱敏有关,从而导致靶神经元对神经递质的敏感性降低。
许多突触抑制中的一个重要特点就是使用依赖性。
高水平的传递和更大的抑制有关,并且基线传递的降低(例如,通过降低胞外Ca2+浓度)可以减轻抑制。
通过突触前受体的传递的修饰大多数突触前终末都拥有很多不同种类的促代谢型G蛋白耦联受体和促离子型受体(MacDermott et al, 1999)。
递质释放的概率p就是断定突触传递效能的一个重要因素,而p 值部分决定于这些受体被结合的程度,而后者则取决于胞外相关受体激动剂的浓度。
虽然如此,通过暂时提高各种突触前神经调质的浓度,突触活性可以进一步增加受体的占用(Thompson et al, 1993;Miller,1998)。
根据它们的特性,这些受体的激活可以增强或者抑制突触的传递。
通过释放一系列不同的神经调质,突触后细胞也可以影响突触前终末递质的释放。
一个普遍的现象就是,当突触后较强去极化时,树突就会释放逆行性信使作用于突触前膜的G 蛋白耦联受体,从而影响递质的释放。
在一些特定的细胞类型中已经被确定的逆行性信使有:多巴胺,强啡肽,谷氨酰胺,GABA,NO,脑衍生神经营养因子(BDNF),催产素。
尽管突触后膜Ca2+通道依赖小泡的融合是逆行性信使释放的一个普遍的机制,逆行性信使也可以通过非小泡的机制释放。
例如,关于介导逆行性信使信号的系统涉及到突触后膜内源性大麻素类的释放,例如:----------------,它们根据需求,通过切割磷脂来合成,并且可以被突触前膜终末的CB1受体识别。
内源性大麻素类释放的机制还不明确,并且这可能还涉及到一个能够易化穿膜的转运体。
通过突触后膜释放大麻素类,这样的逆行性信号传导可以被突触后膜较强的去极化或者促代谢型受体的激活而引发。
同时,已经表明,逆行性信号传导可以在几个脑区暂时性抑制抑制型和兴奋性突触。
重要的是,在纹状体,有一种可以被内源性大麻素类释放所引发的长时程抑制(LTD)的形式。
神经胶质细胞相关的短时程突触可塑性越来越多证据表明神经胶质细胞可能与一些短时程突触可塑性的形式相关。
由于它们和突触很近,星形胶质细胞和突触旁施万细胞可以在位置上很好的调节突触的活动。
它们有确定的清除神经递质的功能,并且很多通过控制这种清除的速度和广度来参与突触的可塑性。
这可以反过来影响突触后膜被激活或者脱敏的程度。
胶质细胞影响突触可塑性的另外一种途径就是:通过感受细胞外的信使,然后释放一些物质反过来直接影响突触的传递效能。
例如,胶质细胞可以表达许多不同的神经递质的受体(例如,谷氨酰胺的受体),当这些受体被激活时,可以释放物质(例如ATP)作用于突触前膜终末从而调节神经递质的释放。
短时程突触可塑性的功能通过对简单有机体(例如,雨虎属)的研究,短时程突触可塑性最初被认为在行为上起到重要作用。
在哺乳动物的大脑,短时程突触可塑性一个重要的结果就是影响突触的信息加工的功能,使这些突触可以表现出有很多特性的过滤器的功能。
例如,起始p比较低的突触在功能上可以作为高通过滤器,这是由于当高频动作电位爆发时它们会易化,而低频的动作电位的爆发将不能以同样的效能传导。
与此相反,起始p比较高的突触在功能上可以作为低通过滤器,这是由于当高频动作电位的爆发时它们会抑制传导,却可以确实地转播低频动作电位的爆发。
突触的滤过特性可以通过改变p来调节。
由于神经调质的释放通过激活突触前膜受体来降低p,而产生过滤功能。
并且它改变了突触的滤过特性,导致易化大于抑制。
这样一来,突触前膜的抑制可以将一个突触从低通过滤器变为高通过滤器。
长时程突触可塑性人们普遍认为,任何类型的经历都会至少部分地修饰后续的行为,通过突触效能的活性依赖的长时程修饰。
大脑将内部和外部发生的事情作为一个复合体进行编码,大量神经元总体的空间-时间模式的活动可以被定义为:神经回路。
定义一个特定神经回路的重要特点就是:构成回路的突触中连接个体神经元的突触比重的模式。
这个假设的推论就是:当一个神经回路中的活动导致一个突触比重模式的长时程的改变时,新的信息(记忆就形成了)就产生了。
这个观点是100年前西班牙诺贝尔获得者Saniago Ramon y Cajal提出的,后来在1940s末被Donald Hebb所发展,他认为脑中相关记忆的形成是通过一个突触修饰的过程,当突触前膜活动与后膜激动相关时,突触修饰加强了联系。
这个提出的关于突触可塑性的功能,形成了发现两个一致性事件的记忆线索,提示了行为现象(例如Pavlovian经典条件反射)的细胞基础。
这种长时程,活性依赖的突触传导效能的改变存在性的实验支持一直是缺乏的,直到1970s早期Bliss和他的同事报道了海马区兴奋性突触的连续性刺激会导致突触传导效能的增强,这种增强可以持续几个小时甚至几天。
在过去三十年,这种现象,最终被定义为长时程增强(LTP),并且开始被人们深入的研究,因为人们普遍认为LTP对于理解记忆形成的细胞和分子机制有重要帮助。
尽管仍然被认为具有代表性,但是现在人们了解到海马区的LTP只是存在于哺乳动物脑中特定回路中几种不同形式长时程突触可塑性的其中一种。
重要的是,明确了,大多数表现出LTP的突触也表现出了一种或几种形式的LTDs。
因此,一个重要的概念就是:兴奋性突触的传递效能可以通过不同的活动形式进行双向修饰。
并且,人们清楚的认识到名词LTP 和LTD描述了这样一个现象,它的机制根据它们作用的神经回路而改变。
最近确定的其他的突触可塑性的形式包括:homeostatic plasticity和metaplasticity. homeostatic plasticity的主要形式就是突触缩放(synaptic scaling), 它描述了:一个给定细胞上的所有突触的传递效能都被活性的长时间改变调节而产生应答。