05 真核生物的遗传分析
- 格式:doc
- 大小:165.50 KB
- 文档页数:15
第七章真核生物的遗传分析重点:真核生物的基因组;真菌的遗传分析;真核生物重组的分子机制。
难点:顺序四分子分析。
第一节真核生物基因组一、C值悖论二、N值悖论三、真核生物基因组DNA的复杂度一、C值悖论基因组(genome):一个物种单倍体的染色体数目及其所携带的全部基因称为该物种的基因组。
genome -- The complete set of sequences inthe genetic material of an organism. Itincludes the sequence of each chromosomeplus any DNA in organelles.C值(C-value):是指生物体的单倍体基因组所含DNA总量。
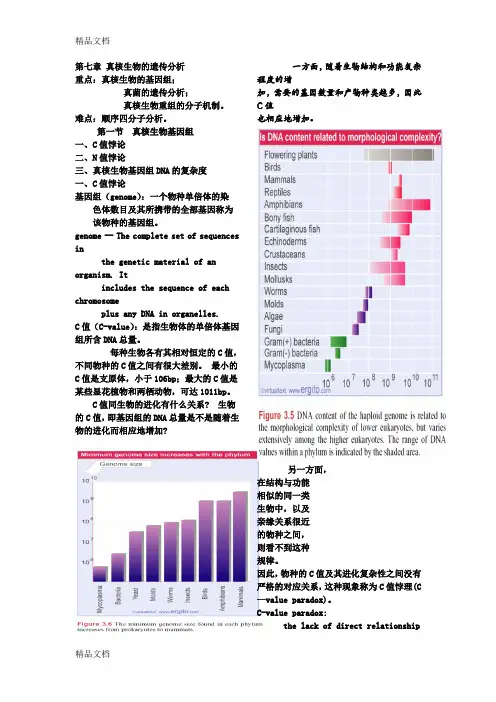
每种生物各有其相对恒定的C值,不同物种的C值之间有很大差别。
最小的C值是支原体,小于106bp;最大的C值是某些显花植物和两栖动物,可达1011bp。
C值同生物的进化有什么关系? 生物的C值,即基因组的DNA总量是不是随着生物的进化而相应地增加?一方面,随着生物结构和功能复杂程度的增加,需要的基因数量和产物种类越多,因此C值也相应地增加。
另一方面,在结构与功能相似的同一类生物中,以及亲缘关系很近的物种之间,则看不到这种规律。
因此,物种的C值及其进化复杂性之间没有严格的对应关系,这种现象称为C值悖理(C —value paradox)。
C-value paradox:the lack of direct relationshipbetween the C value and phylogenetic complex.人们对C值悖理已经提出许多解释:包括基因组的部分或完全加倍、转座、返座已加工假基因、DNA 复制滑动、不等交换和DNA扩增等。
Petrov等又提出一个解释是:各种生物基因组的大小是由于基因组中长期积累起来的过量的非编码DNA被清除的速率不同所造成的结果,即DNA丢失的速率愈慢,那么基因组DNA含量愈高。



第五章真核生物的遗传分析教学目的和要求:1. 理解四分子分析的原理和遗传作图的方法;2. 了解真核生物重组的机制3. 掌握各种遗传标记的特点及应用。
教学重点和难点:【教学重点】1. 四分子分析的原理和遗传作图的方法;2. 重要分子标记分析原理和方法。
【教学难点】应用四分子分析进行遗传作图的原理与方法;重要分子标记分析原理与具体实验环节之间的有机联系。
教学内容:第一节真菌类的染色体作图一.两个连锁基因的作图二.三个连锁基因的作图三.红色面包霉染色体的着丝粒作图第二节真核生物重组的分子机制一.同源重组的发生二.同源重组的分子模型三.有丝分裂分离与重组第三节基因转变及其分子机制一. 异常分离与基因转换二. 基因转变及其分子机制第四节真核生物遗传标记的特点及应用一. RFLP标记二. VNTR和STR标记三. SNP标记四. 分子标记遗传图谱的应用第一节真菌类的染色体作图除了二倍体的高等植物具有连锁和交换的遗传现象以外,单倍体的真菌也有连锁和交换的遗传现象。
对单倍体真菌的遗传分析和染色体作图通常采用四分子分析(tetrad analysis)的方法。
一次减数分裂的四个子细胞称为四分子,对四分子进行遗传学分析称为四分子分析。
根据减数分裂产物在子囊中排列是否有序将四分子分析可分为有序四分子分析和无序死分子分析两种。
有序四分子分析是指根据一个子囊中四个按严格顺序直线排列的四分子表现进行的遗传分析,也称为有序四分子分析。
无序四分子或八分子中孢子没有特殊的顺序,因此也就不能进行着丝粒作图,但无序四分子也可用于各种减数分裂分离和重组分析。
前者以对粗糙链孢霉(Neurospora crassa,2n=14)的遗传分析为代表,而后者主要是对酿酒酵母(Saccharomyces cerevisiae 2n=34)的遗传分析。
粗糙链孢霉,又称红色面包霉,在分类学上属于真菌中的子囊菌纲、球壳目、脉孢菌属,目前已知有4~5种。




第五章真核生物的遗传分析教学目的和要求:1. 理解四分子分析的原理和遗传作图的方法;2. 了解真核生物重组的机制3. 掌握各种遗传标记的特点及应用。
教学重点和难点:【教学重点】1. 四分子分析的原理和遗传作图的方法;2. 重要分子标记分析原理和方法。
【教学难点】应用四分子分析进行遗传作图的原理与方法;重要分子标记分析原理与具体实验环节之间的有机联系。
教学内容:第一节真菌类的染色体作图一.两个连锁基因的作图二.三个连锁基因的作图三.红色面包霉染色体的着丝粒作图第二节真核生物重组的分子机制一.同源重组的发生二.同源重组的分子模型三.有丝分裂分离与重组第三节基因转变及其分子机制一. 异常分离与基因转换二. 基因转变及其分子机制第四节真核生物遗传标记的特点及应用一. RFLP标记二. VNTR和STR标记三. SNP标记四. 分子标记遗传图谱的应用第一节真菌类的染色体作图除了二倍体的高等植物具有连锁和交换的遗传现象以外,单倍体的真菌也有连锁和交换的遗传现象。
对单倍体真菌的遗传分析和染色体作图通常采用四分子分析(tetrad analysis)的方法。
一次减数分裂的四个子细胞称为四分子,对四分子进行遗传学分析称为四分子分析。
根据减数分裂产物在子囊中排列是否有序将四分子分析可分为有序四分子分析和无序死分子分析两种。
有序四分子分析是指根据一个子囊中四个按严格顺序直线排列的四分子表现进行的遗传分析,也称为有序四分子分析。
无序四分子或八分子中孢子没有特殊的顺序,因此也就不能进行着丝粒作图,但无序四分子也可用于各种减数分裂分离和重组分析。
前者以对粗糙链孢霉(Neurospora crassa,2n=14)的遗传分析为代表,而后者主要是对酿酒酵母(Saccharomyces cerevisiae 2n=34)的遗传分析。
粗糙链孢霉,又称红色面包霉,在分类学上属于真菌中的子囊菌纲、球壳目、脉孢菌属,目前已知有4~5种。
利用粗糙链孢霉进行遗传学分析有如下优点:①个体小,生长快,容易培养;②既可进行有性繁殖,又可进行无性繁殖,一次杂交可产生大量后代;③染色体与高等生物一样,研究结果可广泛应用于遗传学上;④无性世代是单倍体,没有显隐性,基因型可以直接在表型上反映出来;⑤一次只需分析一个减数分裂的产物,就可以观测倒遗传结果,简单易行,而二倍体合子是两个不同减数分裂产生的配子相互结合的结果,需要通过测交实验才能分析减数分裂的结果,手续麻烦。
因此粗糙链孢霉是进行基因分离和连锁交换遗传分析的好材料。
粗糙链孢霉的营养体是由单倍体(n=7)的多细胞菌丝体和分生孢子所组成,生活方式由有性和无性两种。
菌丝经有丝分裂直接发育成菌丝体,称无性生殖。
而两种不同接合型细胞结合产生有性孢子的过程称有性生殖。
无性繁殖过程,由菌丝顶端断裂形成分生孢子。
分生孢子有两种,小型分生孢子中只含有一个核,大型分生孢子有几个核。
分生孢子萌发成菌丝,可以再生成分生孢子,周而复始。
酿酒酵母(n=17)的细胞有两种生活形态,单倍体和二倍体。
单倍体的生活史较简单,通过有丝分裂繁殖。
在环境压力较大时通常则死亡。
二倍体细胞(酵母的优势形态)也通过简单的有丝分裂繁殖,但在外界条件不佳时能够进入减数分裂,生成一系列单倍体的孢子。
单倍体可以交配,重新形成二倍体。
酵母有两种交配类型,称作a和α,是一种原始的性别分化,因此很有研究价值。
一.两个连锁基因的作图酿酒酵母、构巢曲霉和单细胞藻类中的衣藻的每一个子囊中的8个子囊孢子的排列是杂乱无序的。
这类真菌的遗传分析可采用非顺序四分子分析(unordeted tetrad analysis)方法。
以酿酒酵母为例,如果要研究A、B基因是否连锁,并计算图距。
首先要明了当AB×ab杂交时,无论有无连锁,只产生下列3种可能的无序四分子。
因为这些子囊是无序排列的,所以尽管第一种类型(亲二型)看起来似乎是两个座位都同MⅠ模式,实际上不是这样,这些孢子以任意顺序写出来都是等效的。
这些子囊仅仅是按照它们是包含二型(ditypes)还是四型(tetratypes)来划分的。
在二型中,两种基因型要么是亲二型(PD)要么是非亲二型(NPD),四型子囊则是由T所表示的。
我们可以试着写一些子囊,证明除了PD、NPD和T之外,没有别的子囊型。
观察这3种子囊类型(即四分子类型),可以发现只有NPD和T有重组型,故这两种四分子类型是决定重组频率的关键。
由于T只有巨1/2重组型,所以当我们已知NPD及T的百分数后,用下列公式求:若求出RF=0.5,可以断定A、B基因无连锁。
如果RF<0.5,则两基因座连锁。
并可以用这个百分数表征其图距。
如本章先前所述,这个RF值也可能低估了图距,其原因仍然是没有考虑双交换和多交换的缘故。
当然,PD、NPD和T的频率可用于校正双交换。
首先,我们要了解PD、NPD和T的四分子类型是如何在连锁的标记基因的杂交中形成的。
假设基因a、b连锁,如果在减数分裂过程中a、b间可以发生非交换(no crossovers,NCO)、单交换(single crossover, SCO)或双交换(double crossovers,DCO)。
用图表示减数分裂形成的这些无序子囊。
应注意到NPD是四线双交换产物。
如果假设双交换在4条染色单体间随机发生。
因此,可以推论4个DCO的频率是相同的。
这就意味着NPD类型包括了1/4的DCO,则可以表示为:DCO=4NPD。
单交换也可以用同样方法计算。
注意到T型四分子既可来自单交换(SCO),也可来自双交换(DCO)。
但我们可以估计T型中来自DCO减数分裂的部分是2NPD,故SCO的大小可以用下式表示:SCO=TT-2NPD。
最后,NCO的大小可依公式:NCO=1-(SCO+DCO)求得。
当我们已估算出这一区域上NCO、SCO、DCO的值之后,可以由这些数值来推导出m值,平均交换次数m=SCO+2DCO=(TT-2NPD)+2(4NPD) =TT+6NPD将m转换为图距单位图距=50×(TT+6NPD )二.三个连锁基因的作图(自学)1. 分类:确定由不交换、单交换、双交换产生的类型与频率;2. 确定基因顺序:根据三点测验中双交换类型频率确定基因顺序的原理,利用频率最小的包括两种亲本型孢子和两种重组型孢子的双线双交换类型确定基因在染色体中的顺序;3. 遗传距离估算:根据相邻基因间重组率计算遗传距离,绘制连锁图。
三.红色面包霉染色体的着丝粒作图1. 着丝粒作图①概念:以着丝粒作为一个座位,测定某一基因与着丝粒之间的距离,并进行基因在染色体上的位置作图。
②原理:A 如果着丝粒与某一对杂合基因之间未发生交换,则该基因与着丝粒同步分离。
此时,一对等位基因的分离为减数第一次分裂分离,即M1,形成非交换型子囊。
B 如果基因与着丝粒之间发生了交换,则该基因与着丝粒的分离不同步。
此时,一对等位基因的分离为减数第二次分裂分离,即M2,形成交换型子囊。
C 当两对以上等位基因分离时,有序四分子分析不仅可以确定它们与着丝粒的遗传距离,还可以分析它们之间是否连锁。
假设a 与b 两个位点,它们的关系仅有3种可能性:(1)a 与b 分别位于不同的染色体上(2)a 与b 位于同一染色体上着丝粒的两侧(3)a 与b 位于同一染色体着丝粒的同侧ab ×++杂交的子囊类型与数目分析:(1)PD>>NPD ,a 与b 连锁(2)若a 与b 在着丝粒两侧,则a MII 分离、b MI 与a MI 分离、b MII 分离两种情况均会发生,本例中无此情况,表明a 和b 在着丝粒的同侧。
(3)由于a MII 分离的同时,多数情况下b 也发生了MII 分离,可判断a 位于着丝粒与b 之间。
(4)a 与着丝粒之间的遗传距离=(1/2)×(60/400)=7.5%=7.5 cMa 与b 间的遗传距离=50×(125/400+6×5/400)=19.4 cM孢子对 子囊类型① ② ③ ④ 1-2ab a+ ab ab 3-4ab a+ a+ ++ 5-6++ +b +b ab 7-8++ +b ++ ++ 数目210 5 125 60 分类PD NPD TT PD a 分离MI MI MI MII b 分离MI MI MII MII 26.9第二节真核生物重组的分子机制重组(recombination)是已经存在的遗传物质产生新的组合的过程。
分子间或染色体间重组(真核染色体减数分裂时独立分配和病毒基因组片段的重新分配); 而分子内或染色体内重组是酶依赖的过程,新的遗传物质通过DNA的剪切和连接产生。
有五种类型的分子内重组,每种情况中分子反应——DNA 剪切、双链间的链交换、DNA修复和去除——是相似的。
重组和突变是遗传变异的两种截然不同的机制:重组是已经存在的信息重排,而突变是在基因组中导入新的信息。
突变和重组在分子水平上是相互关联的。
很多重组事件(特别是转座和不正常重组)会导致基因破坏,这称为突变。
相反的,一些重组是修复潜在突变或致死DNA损伤所必需。
重组的两种应用方式:对遗传座位作图和基因及基因组的操作。
遗传作图根据染色体上两个基因座距离越远越容易发生同源重组的原理进行。
因而一个特定杂交的重组产物的比例提供了对物理距离的估测。
同源重组可被用于基因寻靶、而位点专一性重组可以诱导缺失和染色体重排。
转座和不正常重组也被用来基因转移和整合。
分子内重组的不同类型:1 同源重组(Homologous recombination)重组对之间需要有同源性。
调节这一过程的蛋白(如大肠杆菌中的RecA)不是序列专一性的,而是同源依赖的。
经常涉及长的同源区域(如减数分裂中)。
2 位点特异性重组(Site-specific recombination)重组对之间不需要同源性。
调节这一过程的蛋白(位点特异性重组酶)在供体和受体分子中识别短的,特异DNA序列,这些蛋白之间的相互作用帮助重组。
供体和受体位点之间经常存在同源性,因为同样的重组酶蛋白可结合两者的识别位点。
3 转座重组(transposition recombination)重组对之间不需要同源性。
调节这一过程的蛋白(转座酶,整合酶)识别重组分子中的短特异序列,这样的序列称为转座因子(识别位点经常是转座因子和宿主DNA之间的接合处)。
受体位点一般在序列上相对非特异,重组将可转座因子整合到宿主DNA中(见第13章可移动的遗传因子)。
4 不正常重组(illegitimate recombination)重组对之间不需要同源性或少量同源性,是异常细胞作用的结果。
包括复制中不正常末端连接,链滑动或成环。
不等交换(参阅)经常被描述成不正常,尽管其机制是正常的(但在错误的地方发生)同源重组。