真核生物遗传规律分析
- 格式:ppt
- 大小:854.00 KB
- 文档页数:15
第七章真核生物的遗传分析重点:真核生物的基因组;真菌的遗传分析;真核生物重组的分子机制。
难点:顺序四分子分析。
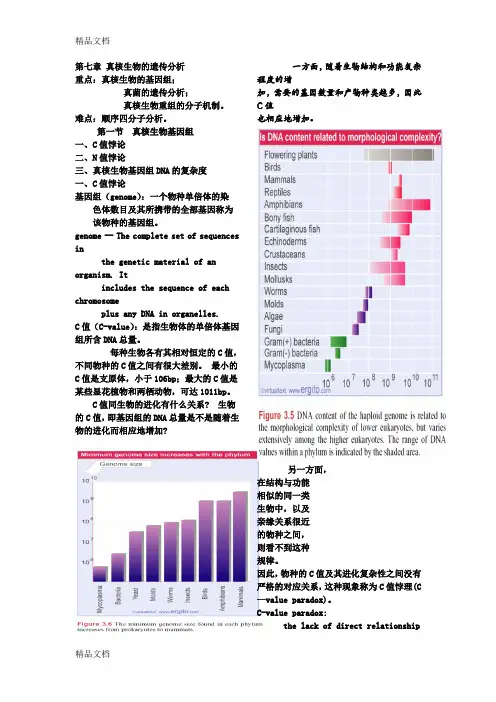
第一节真核生物基因组一、C值悖论二、N值悖论三、真核生物基因组DNA的复杂度一、C值悖论基因组(genome):一个物种单倍体的染色体数目及其所携带的全部基因称为该物种的基因组。
genome -- The complete set of sequences inthe genetic material of an organism. Itincludes the sequence of each chromosomeplus any DNA in organelles.C值(C-value):是指生物体的单倍体基因组所含DNA总量。
每种生物各有其相对恒定的C值,不同物种的C值之间有很大差别。
最小的C值是支原体,小于106bp;最大的C值是某些显花植物和两栖动物,可达1011bp。
C值同生物的进化有什么关系? 生物的C值,即基因组的DNA总量是不是随着生物的进化而相应地增加?一方面,随着生物结构和功能复杂程度的增加,需要的基因数量和产物种类越多,因此C值也相应地增加。
另一方面,在结构与功能相似的同一类生物中,以及亲缘关系很近的物种之间,则看不到这种规律。
因此,物种的C值及其进化复杂性之间没有严格的对应关系,这种现象称为C值悖理(C —value paradox)。
C-value paradox:the lack of direct relationshipbetween the C value and phylogenetic complex.人们对C值悖理已经提出许多解释:包括基因组的部分或完全加倍、转座、返座已加工假基因、DNA 复制滑动、不等交换和DNA扩增等。
Petrov等又提出一个解释是:各种生物基因组的大小是由于基因组中长期积累起来的过量的非编码DNA被清除的速率不同所造成的结果,即DNA丢失的速率愈慢,那么基因组DNA含量愈高。

真核生物的基因组结构与功能分析真核生物是指在生命进化过程中逐渐形成的一类生物,其基本特征之一是存在真核细胞核。
真核生物的基因组结构较为复杂,包含多个线性染色体和一些质粒。
对基因组结构的分析与理解,对于揭示其生物功能和进化机制是至关重要的。
一、真核生物的基因组结构真核生物的基因组大小较大,同一物种不同个体之间的基因组大小存在较大的差异。
基因组大小与细胞大小和复杂度之间存在着类似关联性。
人类基因组大小约为3亿个碱基对,其中蛋白编码基因仅占大约2%。
真核生物的基因组在基本结构上与细菌大相径庭,主要包括以下几个方面。
1. 染色体染色体是真核生物中最重要、最基本的遗传物质,是基因在生物体内的物质传递介质,是遗传信息的载体。
在精细结构上,真核细胞中存在很多复杂的染色体结构,如核小体、类固醇激素受体、平衡染色体等。
2. 基因组复制真核生物的基因组复制主要包括原核生物和真核生物的不同模式,其中原核生物中存在着DNA单线复制机制,而真核生物则采用DNA复制机器进行自我复制。
与原核生物不同的是,真核生物的DNA复制机器必须满足染色体的线性特性和复杂的三维结构,包括多个酶和蛋白质。
3. 基因只读基因只读是指通过读取基因组中的基因序列,进而达到生物高效功能表达和调节的过程。
真核生物基因组的序列阅读具有高度异质性,不同物种、不同个体之间存在大量的序列差异,这在一定程度上阻碍了对真核生物的功能研究。
二、真核生物的基因组功能分析真核生物的基因组分析主要包括以下几个方面。
1. 蛋白编码基因预测蛋白编码基因是真核生物基因组的重要组成部分,对真核生物的基因组进行蛋白编码基因预测,可以揭示其生物功能和进化机制。
目前,已经建立了多种基于序列、结构、相对位置等的蛋白编码基因预测算法与工具,如Glimmer、InterProScan、Pfam等。
2. 生物信息分析真核生物的基因组分析需要大量的计算资源和分析工具,这就需要借助生物信息学的手段来实现。


二RNA的生物合成(转录)生物体以DNA中的一条单链为模板,NTP为原料,在DNA依赖的RNA聚合酶催化下合成RNA链的过程称为转录。
真核生物一个mRNA分子一般只含有一个基因,编码产物为单顺反子。
原核生物的一个mRNA分子通常含有多个基因,编码产物为多顺反子。
(一)转录体系:DNA模板、4种NTP、RNA聚合酶某些蛋白因子和必要的无机离子。
(二)转录DNA模板RNA转录模板并非DNA的全部基因,而是DNA链上区段结构基因。
发生转录的链成为模板链,相对应的另一条链为编码链,模板链并不是总在同一条链上。
(三)转录特点:不对称转录,边转录边翻译(原核生物)(四)RNA聚合酶:原核生物和真核生物RNA聚合酶种类不同,原核生物中RNA聚合酶可以直接起始转录合成RNA,真核生物则不能。
在真核生物中,三种RNA聚合酶都必须在蛋白质转录因子的协助下才能进行RNA的转录。
主要见表3(五)原核生物以操纵子为一个转录单位。
表3原核生物和真核生物RNA聚合酶的特点原核生物RNA聚合酶真核生物RNA聚合酶1种(RNA-pol),5个亚基(α2ββ'ζ)3种(RNA-polⅠ、Ⅱ、Ⅲ)RNA-pol具有合成mRNA,rRNA,tRNA的功能,没有校对功能,缺乏3'→5'外切酶活性α2 位于启动子上游,决定哪些基因被转录β与底物NTP结合,形成磷酸二酯键β'酶与模板结合的主要部位ζ辨认起始点(无催化活性)、Ⅱ、Ⅲ由于识别不同的启动因子而分别识别不同的基因。
转录RNA-polⅠ定位核仁,转录45S- rRNARNA-polⅡ定位核浆,转录产生hnRNARNA-polⅢ定位核浆,转录产生tRNA,5S- rRNA,snRNA.(三)DNA复制的过程原核生物和真核生物DNA的过程大致可分为:起始+延长+终止三个阶段。
1、起始阶段表2(1)解链/旋,解链/旋酶催化。
(2)起始点识别。
(3)原核生物形成复制叉。



原核生物与真核生物的遗传物质与基因组织结构的差异按照细胞的结构和遗传物质在细胞内的分布可将生命有机体划分为原核生物和真核生物两大类。
噬菌体和病毒既不是原核生物也不是真核生物它们是一种超分子的亚细胞生命形式它们的遗传物质是DNA或RNA。
特征原核生物真核生物核膜无有不同染色体数目11核小体结构无有核仁无有遗传交换质粒介导单向配子融合DNA是原核生物染色体的主要组成成分含量占染色体的80以上其余为RNA和蛋白质。
原核生物的遗传物质一般为环状DNADNA存在于细胞内相对集中的区域一般称为拟核nucleoid但并无核膜包裹。
拟核当中的DNA只以裸露的核酸分子存在虽与少量蛋白质结合但不形成染色体结构。
当然它还有一些位于拟核之外的遗传物质——质粒和转座因子。
真核生物中也含有转座因子原核生物一般只有一条染色体即一个核酸分子DNA或RNA而且染色体DNA大多数以双链、共价闭和、环状的形式存在。
多少年来一直以为原核生物的单一环状染色体是区别于真核生物中的多条线状染色体的最好标志。
然而越来越多的研究证明除单一环状的染色体外有些细菌具有多条环状染色体还有些细菌具有线状染色体。
如根癌土壤杆菌含有2条染色体其中一条是长度为3.0Mb的环状染色体另一条是长度为2.1Mb的线状染色体原核细胞中含有一些DNA结合蛋白它们与DNA结合后帮助DNA进行高度折叠。
这些参与DNA折叠的蛋白质称为类组蛋白histone-like protein。
除类蛋白外DNA还与其他蛋白质相结合如与复制、转录和加工有关的蛋白质结合在一起这样其环状染色体DNA以紧密缠绕的、致密的、不规则小体形式存在该小体即是拟核。
真核生物基因组与原核生物基因组有很大的差异真核生物基因的结构、基因表达的过程、表达调控等方面都远比原核生物复杂。
真核生物和原核生物的最大差异之一是遗传物质的分布和存在状态。
原核细胞的遗传物质是以裸露DNA或RNA的形式位于拟核之中而真核细胞的遗传物质是以与组蛋白和非组蛋白相结合缠绕成多条染色体的形式集中于细胞核中。


原核生物与真核生物的差异原核生物指的是一大类细胞核无核膜包裹,只存在称作核区的裸露DNA的原始单细胞生物,包括真细菌和古生菌两大类群。
而真核生物是一大类细胞核具有核膜,能进行有丝分裂,细胞质中存在线粒体或同时存在叶绿体等多种细胞器的生物。
并且真核生物种类特别多,小到微生物,大到动植物,都是真核生物。
原核生物与真核生物(此处仅指微生物)的差异可从下面四个方面进行讨论。
一、细胞结构方面:原核生物没有细胞骨架,而真核生物有细胞骨架。
原核生物的鞭毛属于亚显微结构,细而简单;而真核生物的鞭毛属于显微结构,有微管组成,并且粗而复杂。
原核生物的细胞壁中多糖的主要成分是肽聚糖,而真核生物的主要成分是纤维素和几丁质。
原核生物的细胞膜也不含甾醇和胆固醇,但有呼吸或光合的组分,真核生物与之相反。
原核生物的细胞质膜也常缺少固醇,细胞质结构也简单,没有微管结构,没有流动性,只有核糖体这一种细胞器,呼吸系统位于质膜或中体。
相比之下真核生物的细胞质结构就复杂得多。
另外,原核生物的核糖体是70S的,而真核生物是80S的。
原核生物没有真正的细胞核,只有无核膜包裹的拟核。
原核生物的拟核中没有核仁,没有组蛋白,仅有由环状DNA分子构成的单个染色体,但其DNA含量高。
在真核生物中染色体是由线状DNA和组蛋白组成的。
原核生物基因的重复顺序、重复次数极少,所占基因组的比率很低,同时重复片段小,没有高、中重复顺序之分。
而在在真核生物基因组中重复顺序DNA 普遍存在,不仅重复次数多,而且所占基因组比率高,并具多样性。
不仅如此,基因组内基因的排列在原核生物中功能相关的基因有高度集中的表现, 且愈是简单的生物愈是集中,而真核生物基因组中功能相关基因的排列更多的是处于分散状态,在一条染色体上常被一些不编码的顺序所隔开。
二、个体形态学方面原核生物的细胞体积一般较小,只有1~10微米,而真核生物体积较大,一般是10~100微米。
原核生物的个体形态也简单,且形状较少,。
真核生物基因特征基因是生物体内控制遗传性状的基本单位,是物种遗传多样性的基础。
真核生物是指细胞核内有真正的染色体和细胞器的生物,包括动物、植物、真菌等。
它们的基因具有以下特征。
1. 基因的物理结构真核生物的基因通常是由DNA序列编码的,而不是RNA。
基因通常位于染色体上,由起始密码子和终止密码子围成。
基因的长度和复杂性因物种而异,可以从几百个碱基对到数百万个碱基对不等。
基因的物理结构决定了它的功能和表达方式。
2. 基因的剪接真核生物的基因通常具有剪接功能,即一个基因可以编码多种不同的蛋白质。
这是因为基因的内含子和外显子不是一一对应的,而是可以组合成不同的剪接方式。
这种剪接方式可以增加基因的多样性和复杂性,使得一个基因可以编码多种不同的蛋白质,从而实现更为复杂的生命功能。
3. 基因的表达基因的表达是指基因转录为RNA,然后再翻译为蛋白质的过程。
真核生物的基因表达受到多种因素的调控,包括启动子、转录因子、RNA剪接、RNA稳定性、翻译后修饰等。
这些调控机制可以使得基因表达产生差异,从而实现细胞和组织的不同功能。
4. 基因的遗传方式真核生物的基因遗传方式包括显性遗传和隐性遗传。
显性遗传是指基因表现出来的遗传特征可以直接观察到,而隐性遗传则是指基因表现出来的遗传特征需要通过基因型分析才能确定。
真核生物的基因遗传方式是复杂的,受到多种因素的影响,包括基因型、环境、表观遗传等。
5. 基因的突变基因突变是指由于DNA序列发生变化而导致基因表达发生改变的现象。
真核生物的基因突变通常是由于DNA复制错误、DNA损伤、DNA重组等原因引起的。
基因突变可以导致基因表达的改变,从而影响生物的生长、发育和适应性。
总之,真核生物的基因具有多样性和复杂性,是生物体内控制遗传性状的基本单位。
对基因的研究可以帮助我们更好地理解生命的本质和多样性,为人类健康和可持续发展提供基础支撑。
真核基因组结构特点
真核基因组的结构特点主要体现在以下几个方面:
1.真核基因组结构庞大且复杂,含有大量非编码区,其DNA与蛋白
质结合形成染色体,这些染色体被储存在细胞核内,且体细胞内的基因组是双份的,即双倍体。
2.真核生物的基因转录产物为单顺反子,即一个结构基因转录后翻
译成一个mRNA分子,并对应一条多肽链。
3.真核生物基因组中存在大量的重复序列,这些重复序列的长度不
一,短的只有几个核苷酸,长的可达数百甚至上千个核苷酸。
4.真核生物基因具有不连续性,其结构基因的内部存在许多不编码
蛋白质的间隔序列,这些间隔序列被称为内含子,而编码区则被称为外显子。
此外,基因的非编码区较多,多于编码序列。
5.真核基因组远大于原核生物的基因组,并且具有许多复制起点,
而每个复制子的长度较小。
总的来说,真核基因组的结构特点使其能够适应更复杂的生物过程和更高的生物功能需求。
第六章真核生物遗传分析一、名词解释1、C值悖理(C—value paradox):生物基因组的大小同生物在进化上所处的地位及复杂性之间无严格的对应关系,这种现象称为C值悖理。
2、N值悖理(N value paradox):物种的基因数目与生物进化程度或生物复杂性的不对应性,这被称之为N值悖理。
3、同源重组(homologous recombination):同源重组的发生依赖于大范围的DNA 同源序列的联会,主要利用两个DNA分子序列的同源性识别重组对象,负责DNA配对和重组的蛋白质因子没有碱基序列的特异性,只要两条DNA序列相同或接近,重组就可以在此序列中的任何一点发生,重组中两DNA分子相互交换对等的结构部分。
4、位点专一性重组(site-specific recombination):这类重组依赖于小范围的同源序列联会;重组事件只涉及特定位置的短同源序列,或仅涉及特定的碱基序列,重组时发生精确的切割和连接反应,DNA不丢失、不合成,重组发生在特殊位点上,此位点含有短同源序列,有位点专一性的蛋白因子催化。
两个DNA分子并不交换对等的序列部分,而往往是一个DNA分子整合到另一DNA分子中,因此这种重组又叫做整合式重组5、基因共转变(coconversion):基因转变不仅是专一的,而且是有方向的,它不仅涉及单个位点(或基因座),而且涉及染色体的一个区段,如一对含有两个基因差异的突变型杂交时,在某些子囊中可以发生几个基因同时发生转变的现象,称为基因共转变6、末端连接(end-joining):是指断裂的DNA末端彼此相连。
7、链滑动(strand-slippage):是指DNA复制时,由一个模板跳跃到另一个模板所引起的重组。
8、异源双链DNA(heteroduplex DNA):发生重组时,在重组处每个双链都有一段区域是由亲本DNA分子的各一条链组成,该区域称为异源双链DNA。
9、基因转变(gene conversion):由于异源双链DNA区段不对称碱基对的不同修复过程导致一个基因转变为它的等位基因,称为基因转变。
原核和真核生物的遗传机制生命起源于原核细胞,经历了几亿年的演化后才出现了真核细胞。
这两种细胞在遗传机制上有很多不同之处。
本文将介绍原核和真核生物的遗传机制,包括基因的结构、DNA的复制、转录和翻译等方面。
一、原核细胞的遗传机制原核细胞是指没有真核细胞核的细胞,典型的原核细胞包括细菌和蓝藻等。
原核细胞不同于真核细胞的一个主要特点是其遗传物质不在细胞核内,而是自由地分布在细胞质中的一个称为核区的地方。
原核细胞的遗传信息主要由环状DNA分子组成,因此称为“环状DNA分子基因组”。
1、基因的结构原核细胞的基因比真核细胞的基因要简单得多,其基因大小很小,一般仅有几千个碱基对,编码的蛋白质通常为单一的。
原核细胞的基因一般由连续的区域组成,这些区域中编码蛋白质的部分被称为“开放阅读框架”(ORF),ORF中间被一些间隔序列分开。
另外,有些基因只包含RNA,被称为rRNA(核糖体RNA)和tRNA(转移RNA)。
2、DNA的复制原核细胞的DNA是如何复制的呢?细胞开始时,DNA的双链被酶解开形成复制起始点,然后通过招募一系列蛋白质和酶,实现DNA的复制。
DNA复制时,某一段DNA上的双链解开形成复制泡,随后逐渐向两侧扩散,形成两条新的DNA分子。
3、转录和翻译在原核细胞中,RNA的合成和蛋白质的合成没有分开进行,称为“同步转录和翻译”。
RNA合成需要一些促进因子的作用,使得RNA聚合酶能够与DNA结合形成复合物,这个复合物可以在DNA分子上扫描一段开放的DNA后,在RNA合成时从3'端向5'端正向扫描。
RNA合成完成后,mRNA(信使RNA)能够直接进行翻译,将RNA扫描出来的信息转化为氨基酸序列,而形成蛋白质。
二、真核细胞的遗传机制真核细胞是指在细胞核内含有染色体的细胞。
其遗传物质需要绕过一些障碍才能到达细胞质中进行翻译过程。
真核细胞的遗传信息主要由线性DNA分子组成,形成大量的染色体。
1、基因的结构真核细胞的基因由外显子和内含子组成,外显子直接编码氨基酸序列,而内含子虽然位于基因组内,但并不参与氨基酸序列的编码,它们的存在会妨碍蛋白质的合成。
真核生物的染色体结构与表观遗传调控染色体是存在于细胞核内的遗传物质,是遗传信息的基本单位。
在真核生物中,染色体通常以线性结构存在,其中包含了DNA、蛋白质以及其他辅助蛋白质。
在染色体层面,真核生物可以分为两类:一是单一染色体类型,如人类的23对染色体;二是多倍体染色体类型,如植物和一些无脊椎动物的体内存在多份同源染色体。
真核生物染色体的结构通常分为两个部分:核小体和染色质纤维。
核小体由多个小球状蛋白质组成,而染色质纤维有两种状态:萎缩态和膨胀态。
萎缩态的染色质纤维非常致密,难以转录,而膨胀态的染色质纤维则相对松散,转录起来相对容易。
为了更好地理解染色体的结构特点,我们可以将其比喻为一本书。
核小体便是书的章节,每个章节里面都装载着大量的DNA。
而染色质纤维就是书的页码和内容之间的地方,如果书的页码非常密集,并且每个页码的文字内容都非常紧密,里面的信息量就会非常大,读起来也会比较困难。
相反,如果书的一页内容相对松散,那么阅读起来就会更舒适一些,同时一页中信息的呈现也更加清晰。
除了染色体本身的结构特点外,还有一种影响组成染色体的方式的因素,这便是表观遗传学。
表观遗传学是指通过改变DNA不改变序列,从而影响基因表达的机制,这种机制通常发生在细胞分化和认知功能形成等过程中。
有些表观遗传调控的机制是直接影响着染色体的构成方式。
例如,甲基化会促进染色质纤维的萎缩,从而使得染色体在表达时更加紧凑。
而异染色质是指染色体在不同周期呈现不同的表达状态,这种状态通常与蜕变和分化过程相关。
同时,表观遗传调控的机制也可以间接影响着染色体结构的特点。
例如,转录因子可以调节基因的表达状态,从而影响染色体中的DNA部分。
此外,组蛋白也可以通过改变DNA的包裹方式来影响基因表达。
总的来说,真核生物的染色体结构和表观遗传调控是紧密相关的。
染色体结构直接关乎着DNA的转录和表达能力,而表观遗传调控则可以影响着染色体的构成方式以及基因的表达周期和方式。