内共生学说的起源和发展
- 格式:doc
- 大小:279.00 KB
- 文档页数:3
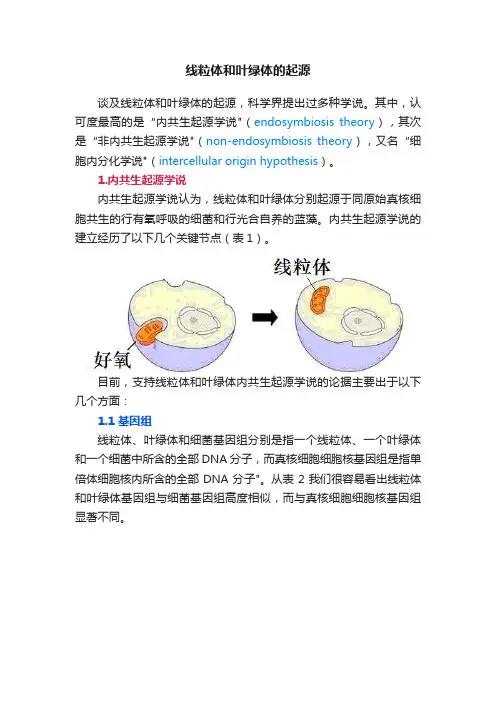
线粒体和叶绿体的起源谈及线粒体和叶绿体的起源,科学界提出过多种学说。
其中,认可度最高的是“内共生起源学说"(endosymbiosis theory),其次是“非内共生起源学说"(non-endosymbiosis theory),又名“细胞内分化学说"(intercellular origin hypothesis)。
1.内共生起源学说内共生起源学说认为,线粒体和叶绿体分别起源于同原始真核细胞共生的行有氧呼吸的细菌和行光合自养的蓝藻。
内共生起源学说的建立经历了以下几个关键节点(表1)。
目前,支持线粒体和叶绿体内共生起源学说的论据主要出于以下几个方面:1.1 基因组线粒体、叶绿体和细菌基因组分别是指一个线粒体、一个叶绿体和一个细菌中所含的全部DNA分子,而真核细胞细胞核基因组是指单倍体细胞核内所含的全部DNA分子"。
从表2我们很容易看出线粒体和叶绿体基因组与细菌基因组高度相似,而与真核细胞细胞核基因组显著不同。
1.2 蛋白质合成系统线粒体、叶绿体蛋白质合成系统与细菌十分接近,而与真核细胞差异较大。
(1)叶绿体tRNA合成酶、氨酰-tRNA合成酶能与细菌tRNA合成酶、氨酰-tRNA合成酶交叉识别,但不能与真核细胞细胞质中的tRNA合成酶、氨酰-tRNA合成酶交叉识别。
(2)线粒体的蛋白质合成因子不能识别真核细胞细胞质中的核糖体,但可以部分取代细菌的蛋白质合成因子。
(3)叶绿体核糖体小亚基与大肠杆菌核糖体大亚基可以组成一个有功能的杂交核糖体,但叶绿体核糖体与真核细胞细胞质核糖体无法形成有功能的杂交核糖体。
(4)叶绿体16S rRNA,23S rRNA,5S rRNA在序列上与蓝藻(蓝细菌)的同源性远高于真核细胞。
除上述4点外,表3和表4比较的6个方面能更加有力地反映出线粒体、叶绿体蛋白质合成系统与细菌的高度相似,与真核细胞的显著不同。
1.3 生物膜线粒体、叶绿体是具有双层膜的细胞器,但其外膜与内膜在性质、成分等方面具有明显差异,外膜与真核细胞内膜系统相似,内膜与细菌质膜相似。
真核细胞的起源摘要: 真核细胞的起源是生物进化过程中重要的环节,在生物进化中有着举足轻重的地位。
对真核细胞起源的研究,不仅有利于解释远古时代各种各样奇异的生命进化现象,更有利于我们对人类的起源有一个更为深刻的认识。
随着科学技术水平的提高,生物学也蓬勃发展起来,对真核细胞起源也有了更进一步的认识,各种各样与真核细胞起源有关的学说也不断地被发表出来。
关键词:真核细胞,起源,内共生性,古核细胞,原细菌目前,围绕着真核细胞起源问题,主要有两种学说——内共学说和经典学说(即内陷说,内膜扩张说)。
但从现有的资料来看,内共生学说有它更多的合理性,更受到学术界的支持。
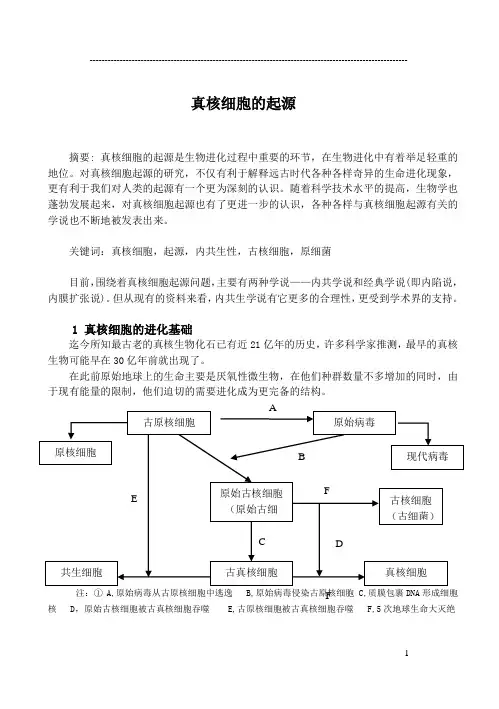
1 真核细胞的进化基础迄今所知最古老的真核生物化石已有近21亿年的历史,许多科学家推测,最早的真核生物可能早在30亿年前就出现了。
在此前原始地球上的生命主要是厌氧性微生物,在他们种群数量不多增加的同时,由于现有能量的限制,他们迫切的需要进化成为更完备的结构。
注:① A,原始病毒从古原核细胞中逃逸 B,原始病毒侵染古原核细胞 C,质膜包裹DNA 形成细胞核 D ,原始古核细胞被古真核细胞吞噬 E,古原核细胞被古真核细胞吞噬 F,5次地球生命大灭绝古原核细胞 原始病毒原始古核细胞(原始古细古真核细胞 共生细胞 真核细胞原核细胞 现代病毒 古核细胞(古细菌) ABCD F FE2 “真核细胞起源”的内共生学说随着生物学的发展,围绕着真核细胞的起源问题,已有较丰富的资料,并形成了多种学说因而有必要作一简要的介细。
目前,围绕着真核细胞起源问题,主要有两种学说——内共生学说和经典学说(即内陷说,内膜扩张说)。
但从现有的资料来看,内共生学说有它更多的合理性,更受到学术界的支持。
2.1 原细菌与真核细胞密切的亲缘关系近年来随着现代分子生物学和微生物学的迅猛发展,特别是一类不同于一般原核生物的生物类群——原细菌的发现,给上述问题的解决带来了希望的曙光。
结合这类生物与真核细胞方面面的亲缘关系,对其进化地位尤其是它们对于揭示真核细胞起源进化问题的意义作些探讨。

内共生学说的主要内容
内共生学说的主要内容为:古生菌细胞先后吞噬了α朊细菌和蓝细菌,并发
生了内共生,从而两者进化成宿主细胞的细胞器一线粒体和叶绿体。
宿主也发展
成各类真核生物。
许多科学家认为,线粒体和叶绿体分别起源于原始真核细胞内共生的细菌和蓝藻。
1970年Margulis在分析了大量资料的基础上提出了一种设想,认为真核
细胞的祖先是一种体积巨大的、不需氧的、具有吞噬能力的细胞,能将吞噬所得的糖类进行酵解取得能量。
而线粒体的祖先——原线粒体则是一种革兰氏阴性菌,含有三羧酸循环所需的酶系和电子传递链,故它可利用氧气把糖酵解的产物丙酮酸进一步分解,获得比酵解更多的能量。
当这种细菌被原始真核细胞吞噬后,即与宿主细胞间形成互利的共生关系,原始真核细胞利用这种细菌(原线粒体)充分供给能量,而原线粒体从宿主细胞获得更多的原料。

内共生学说的起源和发展记得前段时间在准备期中考试时,曾经看到这么一段有意思的话“这些事实都使人设想,真核细胞中的线粒体是由侵入细胞或被细胞吞入的某种细胞经过漫长的岁月演变而来的。
”这确实是对我这个从小对达尔文进化理论深信不移的人产生了很多疑惑。
然而,很多证据都表明线粒体并非真核细胞自生发展而来:(1). 线粒体含有DNA,这些DNA与细胞核中的很不同,却类似细菌的环状DNA。
(2). 线粒体具有和真核宿主细胞不同的遗传密码,这些密码与细菌和古菌中的很类似。
(3). 线粒体被两层或更多的膜所包被,其中最里面一层的成分与细胞中其它膜的都不同,而更接近于原核生物的细胞膜。
(4). 细胞器的大小与细菌相当。
(5). 细胞器的核糖体和细菌相似,细菌的核糖体是70S,线粒体的核糖体是55S。
在查阅这些资料时,一个对我来说完全陌生的词汇被反复提及——内共生学说(endosymbiotichypothesis;endosymbiont theory)。
内共生学说是关于真核生物细胞中的细胞器,线粒体和叶绿体起源的学说。
根据这个学说,它们起源于内共生于真核生物细胞中的原核生物。
这个理论认为线粒体起源于好氧性细菌(很可能是接近于立克次体的变形菌门细菌)。
线粒体祖先原线粒体(一种可进行三羧酸循环和电子传递的革兰氏阴性菌)被原始真核生物吞噬后与宿主间形成共生关系。
在共生关系中,对共生体和宿主都有好处:原线粒体可从宿主处获得更多的营养,而宿主可借用原线粒体具有的氧化分解功能获得更多的能量。
这个理论的证据非常完整,目前已经被广泛接受。
这个学说的确立也可以说是一波三折。
最早提出这些观点的是一群俄国的科学家。
安德雷.谢尔盖耶维奇.法明茨恩(1835-1918)尝试从植物中分离叶绿体并使其生长。
康斯坦丁.谢尔盖耶维奇.梅里日可夫斯基(1855-1921)发展了“双原生质”(twoplasm)理论(细胞内的细胞),宣称叶绿体起源于蓝绿藻。

第六节原核细胞内无成形的细胞核课标内容要求核心素养对接描述原核细胞与真核细胞的最大区别是原核细胞没有由核膜包被的细胞核。
通过比较原核细胞与真核细胞的异同点,认同细胞的统一性与差异性。
(生命观念、科学思维)·与真核细胞相比,原核细胞个体微小、结构简单。
·原核生物由单个原核细胞构成。
细菌和蓝细菌属于原核生物。
一、小资料马古利斯和她的“内共生学说”1.学说的倡导者美国科学家琳恩·马古利斯倡导的内共生学说,是关于早期生命演化中细胞如何从简单到复杂的学说。
2.学说的内容马古利斯认为,真核细胞中线粒体、叶绿体等结构起源于早期的原核生物,某些细菌被原始的真核生物吞噬后,经过长期共生成为线粒体;而蓝细菌被吞噬后逐渐演化为叶绿体.3.证据表明越来越多的证据表明内共生学说是正确的,例如,线粒体和叶绿体内的结构更接近原核生物而不是真核生物等。
4.普遍性共生现象在自然界是普遍存在的,如双小核草履虫内的某种结构实际上是一种细菌,一些海洋动物如造礁珊瑚与能进行光合作用的虫黄藻共生。
二、地球上最早岀现的是原核细胞1.化石证据35亿年前的地层中,就发现了早期蓝细菌的化石。
通过对地质岩石分析,可靠的、大量的真核生物化石于10亿~8亿年前才出现。
2.早期地球特点早期地球没有臭氧层,紫外线很强,大气中CO2含量很高,导致地表温度比现在高得多.诸多因素导致早期地球环境并不适合生命生存。
3.蓝细菌的出现改变了地球大气环境蓝细菌在其出现后的10多亿年时间里,一直是地球的主要生物类群。
在如此长的时间里,蓝细菌光合作用释放的氧气逐渐在大气圈积累,为真核生物的起源创造了条件,同时促进了臭氧层的形成,阻挡了大量对生物有害的紫外线;蓝细菌的光合作用促使大气圈中CO2含量下降,将碳元素转移至岩石圈中形成碳酸盐;CO2含量的下降逐渐降低了地球表面的平均温度,为陆生生物的出现创造了条件。
4.真核生物逐渐出现随着地球环境的改变,逐渐出现原始单细胞真核生物。

《进化生物学》作业答案绪论一、名词解释进化:广义进化指的是事物的变化发展。
它包含了宇宙的演化即天体的消长,生物的进化,以及人类的出现和社会的发展。
生物进化:生物在与其生存环境相互适应作用过程中,其遗传系统随时间而发生一系列不可逆的改变,并导致相应的表型改变,在大多数情况下这种改变导致生物体对其生存环境的相对适应。
(张昀)生物进化论:是研究生物界进化发展的规律,以及如何运用这些规律的科学。
它的主要研究对象是生物界的系统发展,当然也包括某一物种或某一完整的生物类群的发展。
其重点是研究生物如何由简单向复杂,由低等向高等的发展过程。
进化生物学就是研究生物进化的科学,不仅包括进化的过程,更重要的是研究生物进化的原因、机制、速率和方向,是回答为什么的科学,是追究事物或过程的因果关系的科学。
进化生物学是在生物进化论随着分子生物学的发展由推论走向验证,由定性走向定量的过程中应运而生的科学。
二、比较拉马克学说和达尔文学说的异同。
相同点:两人都认为生物是可变的,并支持进化论;遗传法则上,拉马克提出“用进废退”和“获得性状遗传”;由于历史的局限,达尔文也认为,生物性状符合“用进废退”和“获得性状遗传”的规律。
不同点:起源上不同:拉马克主张,最原始的生物源于自然发生,生物进化是多元的;达尔文主张物种具有共同起源(一元论)对于进化的机制两人见解不同,拉马克主张“用进废退和获得性状遗传”,强调了环境变化在生物变异方面所起的“诱导”,但却主张变异是以生物本性即内因(一种趋于完善的需求)为主因。
但达尔文认为变异和环境是相互独立的,在环境发生作用前变异就产生了,环境只是对变异其选择的作用,生物(以物种为单位)通过生存斗争,适应环境的性状得到保留,比适应的性状被淘汰(适者生存)。
三、简述现代综合进化论的主要内容。
代表人物:杜布赞斯基,主要著作:《遗传学和物种起源》(1937)主要内容:第一,认为自然选择决定进化的方向,使生物向着适应环境的方向发展。

线粒体内共生学说
线粒体内共生学说是指线粒体与宿主细胞之间存在共生关系的一种学说。
线粒体是细胞内的一个重要细胞器,它通过氧化磷酸化过程产生能量,供给宿主细胞的各种生命活动。
然而,线粒体不仅仅是一个能量供应者,它还在许多细胞过程中发挥着重要的作用。
因此,线粒体与宿主细胞之间的共生关系对于维持生命活动至关重要。
线粒体内共生学说最初是由美国科学家琳达·布克斯提出的。
她在20世纪60年代开始研究线粒体与宿主细胞之间的关系,并发现线粒体不仅具有自我复制和自我维护的能力,还可以与宿主细胞相互作用,参与细胞内的许多重要过程。
在线粒体内共生学说中,线粒体被认为是宿主细胞和细胞内其他细胞器之间的调节中心。
它通过产生能量、参与脂质代谢、调节细胞凋亡等多种方式影响宿主细胞的生命活动。
同时,宿主细胞也可以通过调节线粒体的数量和功能来适应环境的变化。
除此之外,线粒体内共生学说还提出了一种新的生命起源学说,即内共生起源学说。
这一学说认为,早期的细胞可能是由不同类型的细胞共同演化而来的。
其中一个重要的环节就是线粒体和细胞质基因组之间的共同演化。
这种共同演化不仅产生了新的生物种类,也推动了生命的进化和多样性。
线粒体内共生学说是细胞生物学和进化生物学中的重要理论之一。
它不仅揭示了线粒体与宿主细胞之间的共生关系,还为我们理解生命的起源和演化提供了新的思路。
随着科学技术的不断进步,相信线粒体内共生学说的研究将会取得更加深入的进展。

简述真核细胞起源的内共生学说真核细胞起源是生命演化中一个重要的问题,一直以来吸引着科学家们的关注。
内共生学说是一个备受青睐的真核细胞起源假说,认为原核生物和一个或多个细菌形成了共生关系,进而演化成了真核生物。
本文将简述内共生学说的主要观点和证据。
一、内共生学说的主要观点内共生学说认为,早期的原核生物在寻找合适生存环境的过程中,可能遇到了某些具有较强适应能力的细菌。
这些细菌与原核生物建立了共生关系,为原核生物提供了一些基本生存条件,如能量、营养、修复等功能。
在长期的共生过程中,原核生物逐渐发生变异,形成了真核细胞。
共生物也演化成了微粒体、叶绿体等细胞器,进一步强化了真核细胞的生存和繁殖能力。
内共生学说的观点主要有以下几个方面:1. 原核生物和细菌存在相似的细胞结构和代谢途径,这可以解释真核细胞中出现的一些特殊细胞器,例如线粒体和叶绿体,它们的形态和结构类似于细菌。
2. 真核细胞中的染色体有较高的相似性,这可以解释为共生物进化过程中的“基因交换”结果。
3. 微粒体和叶绿体有自主的DNA和蛋白质合成能力,这可能是由于这些细胞器最初是细菌,在共生过程中逐渐进化为了真核细胞的组成部分。
二、内共生学说的证据内共生学说并不是凭空想象而来的,而是有一批实验证据的。
下面将简要介绍几点证据:1. 细胞器DNA的来源微粒体和叶绿体拥有自己独立的DNA,这些DNA部分与大肠杆菌(Escherichia coli)和紫细菌(Rickettsia)等以共生方式存在于真核细胞中的细菌DNA 具有近似,也与古菌(Archaea)或真核DNA无关。
这表明微粒体和叶绿体可能起源于某些能够进行自我复制的细菌。
2. 组成真核生物的实验理论上,如果细菌确实是真核细胞起源的真正祖先,那么它们应该能够组成真正的真核生物。
实验研究表明,用现代细菌模拟原细胞,并与其他类似细胞一起制成一个细胞群落,能够达到类似原细胞的生物状态。
这表明,进化的开始比我们以前想象的可能要小得多。

名词解释1.进化生物学:研究生物进化的科学,研究进化的过程、原因、机制、速度和方向,还研究物种的形成和绝灭、系统发生以及适应性起源的一门学科。
它的基本理论就是进化论。
2.内共生起源学说:真核细胞的线粒体和叶绿体来源于共生的真细菌,运动器来自于共生的螺旋体类真细菌,它们最早被原始真核细胞吞噬进细胞内,与宿主进行长期共生,进而演化为重要的细胞器。
3.生物重演律:生物在个体发育中迅速重演其祖先的主要演化阶段。
即个体发育是系统发育史的简单而迅速的重演。
4.五届说:美国生物学家惠特克将生物界分为动物界、植物界、真菌界、原生生物界和原核界。
5.生物表型的进化:相对于细胞水平和分子水平而提出的,包括形态、行为、生理功能三个方面的进化。
表型进化是以分子进化为基础的,其本质还是基因频率的变异。
6.进化稳定对策〔ESS〕:是基于最适性理论提出的,用于解释动物的各种行为对策的一种理论。
是一种混合对策,是经过长期进化检验的、最稳定的、而且最少受到其他可选对策的干扰。
是一种不可侵犯的对策,一旦确立就会稳定下来。
7.平衡性选择:又称保留不同等位基因的选择。
是指能使两个或几个不同质量性状在群体假设干世代中的比例保持平衡的现象。
这种选择常导致群体中存在两种或两种以上不同类型个体,从而维持遗传学多样性。
8.遗传漂变:指的是由于种群太小引起的基因频率随机增减甚至丧失的现象。
又称为赖特效应,是生物进化的一种重要机制。
9.正态化选择:又称为稳定性选择,即把趋于极端的变异淘汰掉而保留那些中间类型的个体,使生物类型具有相对的稳定性。
10.前进性选择:自然选择最基本的一种类型,包括单向性选择和分裂性选择。
前者通过淘汰一种极端而保留另一极端的变异,使生物类型朝某一方向发展;后者把一个群体中极端变异按不同方向保留下来而减少中间常态性。
11.适合度:也称为适合值,是指某一基因型个体与其他基因型个体相比能够存活并把它的基因传给下一代的能力。
12.遗传平衡定律:哈迪温伯格定律,在理想状态下,〔大的随机交配的额群体,无迁移,突变和选择〕各等位基因的频率和等位基因的基因型频率在遗传中是稳定不变的,即保持着基因平衡。
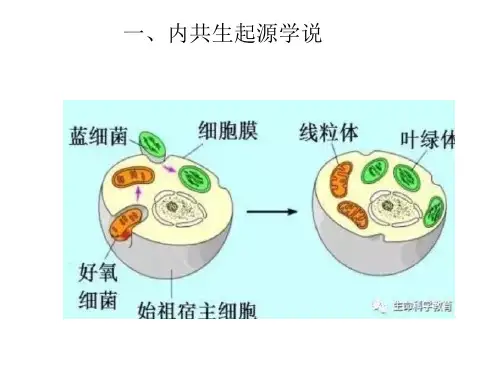
高考专题复习:线粒体和叶绿体的内共生起源学说由于线粒体和叶绿体具有独特的半自主性并与细胞核建立了复杂而协调的互作关系,它们的起源一直以来多被认为有别于其他细胞器。
在人们为这两种细胞器设计的起源假说中,内共生起源学说很好地贴合了线粒体和叶绿体的半自主性和核质关系特征,因而得到了广泛的认可和支持。
(2019年广州一模第29题第3问)内共生起源学说:认为线粒体和叶绿体分别起源于原始真核cell内共生的细菌和蓝藻。
线粒体来源于细菌,即细菌被真核生物吞噬后,在长期共生过程中,通过演变,形成了线粒体。
叶绿体来源于蓝藻,被原始真核cell摄入胞内,在共生关系中,形成了叶绿体。
(纤毛虫(Nassula ornata)与藻类的共生现象)主要论据:①线粒体和叶绿体的基因组在大小、形态和结构方面与细菌的相似。
②线粒体核叶绿体有自己完整的蛋白质合成系统,能独立合成蛋白质。
③线粒体和叶绿体的两层被膜有不同的进化来源,外膜与内膜的结构和成分差异很大。
④线粒体和叶绿体能以分裂的方式进行繁殖,这与细菌的繁殖方式类似。
⑤线粒体和叶绿体能在异源细胞内长期生存。
⑥线粒体的祖先很可能来自反硝化副球菌或紫色非硫光合细菌。
⑦发现介于包内共生蓝藻与叶绿体之间的结构---蓝小体,其特征在很多方面可作为原始蓝藻向叶绿体演化的佐证。
不足之处:a.从进化角度:如此解释在代谢上明显占优势的共生体反而将大量的遗传信息,转移到宿主cell中,不能解释细胞核是如何进化来的,即原核cell如何演化为真核cell。
b.线粒体和叶绿体的基因组中存在内含子,而真细菌原核生物基因组中不含有内含子,不能解释其内含子从何而来。
香梅说:蓝藻可以进行有氧呼吸,为什么植物还需要线粒体? 欢迎各位参与讨论!例1:生物学家提出了“线粒体是起源于好氧细菌”的假说。
该假说认为,在进化过程中原始真核细胞吞噬了某种好氧细菌形成共生关系,最终被吞噬的好氧细菌演化成线粒体。
下列多个事实中无法支持该假说的是()A.哺乳动物细胞的核DNA由雌雄双亲提供,而线粒体DNA则主要来自雌性亲本B.线粒体外膜的成分与真核细胞的细胞膜相似,而内膜则同现存细菌的细胞膜相似C.高等植物细胞的核DNA与蛋白质结合呈线状,而线粒体DNA裸露且主要呈环状D.真核细胞中有功能不同的多种细胞器,而线粒体中仅存在与细菌中类似的核糖体例2.线粒体起源的内共生学说认为,原始真核细胞吞噬了能进行有氧呼吸的原始细菌,它们之间逐渐形成了互利共生关系,最终原始细菌演变成线粒体。
内共生学说的论据内共生学说是一种基于生物学的理论,它认为生物之间存在着内在的、共生的关系。
这种理论得以提出的基础是,所有生命体都是由细胞组成的,而这些细胞之间形成了广泛而复杂的交互作用。
下面我们来探究一下内共生学说的一些重要的论据。
1. 共生是生命起源的过程之一内共生学说认为,生命的起源是由原始的单一生命体开始,然后一些细胞互相合并形成了更复杂的生命体。
之后再逐渐地演化为更高级别的生命体。
这个原理可以从一些现有的生物进化过程中得到证实。
例如,某些单细胞原核生物与非血缘关系的细胞体系已经成功地融为一体,形成了更为复杂的生物体系。
2. 共生有利于生物的进化和适应内共生学说认为,共生关系使不同类型的生物能够互相合作并利用彼此的资源,这有助于它们在环境条件发生变化时适应和进化。
例如,昆虫和细菌的共生关系能够帮助昆虫在营养不足的环境中生存。
此外,有些植物和菌根共生,这种关系有助于植物获取土壤中的营养和水分,有利于它们在干旱或贫瘠的环境中生长和存活。
3. 共生关系可以改变生物的形态和生理学特征内共生学说认为,共生关系可以改变生物的形态和生理学特征。
例如,植物和细菌的共生关系能够使植物根系发生形态和生理学变化,使它们更具有吸收铁和其他矿物质的能力。
此外,多种生物之间的共生关系也能够在生理学和形态上相互影响。
4. 共生关系对生态系统的稳定性有着重要的影响内共生学说认为,共生关系对生态系统的稳定性有着重要的影响。
例如,海洋生态系统中细菌和浮游植物之间的共生关系能够在海洋食物链中扮演重要的角色。
同样,土壤生态系统中植物和真菌之间的共生关系也能够影响腐生物质分解和土壤结构的稳定性。
总之,内共生学说认为,生命的进化和生态系统的稳定性都离不开生物之间的共生关系。
这种理论提供了一种对生命进化和生态系统稳定性更深入的认识。
内共生起源学说名词解释
内共生起源学说是一种科学理论,用来解释不同物种间内共生关系的起源和演化。
它认为不同物种通过长时间的共同生活和相互依赖,发展出彼此互利的关系,最终形成了内共生关系。
根据内共生起源学说,内共生可以通过以下几种方式形成:
1. 互利共生:两个物种之间相互依赖,并从中得到利益。
例如,植物提供食物和庇护给昆虫,在昆虫身上获得传粉服务。
2. 互生共生:两个物种共同生活,相互影响但没有直接利益交换。
例如,一些动物在其他物种身上找到庇护,而这些物种提供了一种保护伞。
3. 共生转变:一个物种开始依赖另一个物种,然后演化成为共生关系。
例如,原本是寄生虫的物种逐渐演化为共生关系,不再对宿主造成伤害。
内共生起源学说提供了一种理论框架,帮助科学家了解内共生关系的起源和对生态系统的影响。
该理论也有助于理解物种之间的相互依赖和进化过程。
在各种细胞器中,线粒体具有特殊性,因其含有核糖体且自身带有遗传物质。
线粒体DNA是环状的,且有一些和标准真核生物遗传密码不同的变化。
这些特性导致了内共生学说——线粒体起源于内共生体。
这种被广泛接受的学说认为,原先独立生活的细菌在真核生物的共同祖先中繁殖,形成今天的线粒体。
这种说法还被应用与科幻小说当中,其中小说寄生前夜说的是,在亿万年间,生物都在不停的进化。
在生物的体内,直接提供能量的线粒体进化速率快于生物本身,以致现在线粒体已经有了意识,并且拥有强大的力量,甚至可以幻化出人形。
于是在某个时刻,线粒体终于爆发了,它们要消灭人类,主宰这个世界。
真核生物起源的内共生起源学说的主要内容
真核生物起源的内共生起源学说是关于真核生物的起源与进化的一种学说,认为真核生物起源于原核生物与内共生的关系演化而来。
该学说的主要内容包括以下几个方面:
1. 内共生关系的形成:起源学说认为,早期的真核生物起源于原核生物(如古细菌或细菌)与其他微生物之间的内共生关系。
原核生物中的一种微生物进入到另一种原核生物中,形成了共生的关系,并逐渐演化为内共生。
2. 合作关系的形成:在内共生的过程中,两种微生物开始合作生活,彼此互利共生。
例如,内共生微生物可以提供重要的代谢产物或复杂的细胞器官,从而帮助寄主细胞生存和繁殖。
3. 合作关系的演化:合作关系的演化过程中,内共生微生物和寄主细胞之间逐渐形成了更加密切的联系。
内共生微生物逐渐演化为真核生物的细胞器官,如线粒体和叶绿体。
这些细胞器官在真核生物细胞中具有特定的功能,如能量产生和光合作用。
4. 结构和功能的演化:内共生微生物逐渐与寄主细胞发生结构和功能上的相互适应。
这种适应性演化使得内共生微生物与宿主细胞之间形成了密切的联系,从而促进了真核生物细胞的复杂化和多样性的发展。
总之,真核生物起源的内共生起源学说认为真核生物起源于原核生物与内共生微生物的合作演化过程。
这种内共生关系导致了真核生物细胞的复杂化和种类的增加。
线粒体的功能与前世今生线粒体(mitochondrion)是一种存在于大多数细胞中的由两层膜包被的细胞器,是细胞中制造能量的结构,是细胞进行有氧呼吸的主要场所,被称为"power house"。
其直径在0.5到1.0微米左右。
除了溶组织内阿米巴、篮氏贾第鞭毛虫以及几种微孢子虫外,大多数真核细胞或多或少都拥有线粒体,但它们各自拥有的线粒体在大小、数量及外观等方面上都有所不同。
线粒体拥有自身的遗传物质和遗传体系,但其基因组大小有限,是一种半自主细胞器。
除了为细胞供能外,线粒体还参与诸如细胞分化、细胞信息传递和细胞凋亡等过程,并拥有调控细胞生长和细胞周期的能力。
一、线粒体主要功能:1. 能量转化线粒体是真核生物进行氧化代谢的部位,是糖类、脂肪和氨基酸最终氧化释放能量的场所。
线粒体负责的最终氧化的共同途径是三羧酸循环与氧化磷酸化,分别对应有氧呼吸的第二、三阶段。
细胞质基质中完成的糖酵解和在线粒体基质中完成的三羧酸循环在会产还原型烟酰胺腺嘌呤二核苷酸(reduced nicotinarnide adenine dinucleotide,NADH)和还原型黄素腺嘌呤二核苷酸(reduced flavin adenosine dinucleotide,FADH2)等高能分子,而氧化磷酸化这一步骤的作用则是利用这些物质还原氧气释放能量合成ATP。
在有氧呼吸过程中,1分子葡萄糖经过糖酵解、三羧酸循环和氧化磷酸化将能量释放后,可产生30~32分子ATP(考虑到将NADH运入线粒体可能需消耗2分子ATP)。
如果细胞所在环境缺氧,则会转而进行无氧呼吸。
此时,糖酵解产生的丙酮酸便不再进入线粒体内的三羧酸循环,而是继续在细胞质基质中反应(被NADH还原成乙醇或乳酸等发酵产物),但不产生ATP。
所以在无氧呼吸过程中,1分子葡萄糖只能在第一阶段产生2分子ATP。
2. 三羧酸循环糖酵解中生成的每分子丙酮酸会被主动运输转运穿过线粒体膜。
简述真核细胞的内共生学说真核细胞内共生学说是指在活细胞内,一些微生物和宿主之间形成相互关系的学说。
它是为了解释宿主细胞内部的微生物和其余有机物质的耦合关系,以及它们影响共同的生物过程,而受到人们越来越多的关注。
实际上,内共生学说认为,宿主和微生物之间是一种相互关联的关系,双方受到彼此的贡献和影响。
宿主微生物共聚关系为真核细胞内共生提供了基础。
宿主微生物共生关系主要指宿主细胞中的微生物群体共同构成的遗传、共享资源的稳定的数量结构模式。
此外,宿主细胞内互共生关系还主要反映在细菌细胞内的基因网络上,例如共同参与的系统生物学复杂性过程。
它可以帮助科学家们分析不同物种之间的关系,进而更好地了解真核细胞内部的功能和结构。
通过内共生学,它允许宿主细胞和细菌细胞之间形成复杂的系统,使它们之间能够进行有效的交流,增强细胞间的相互作用,使它们能够更好地共同服务宿主。
它允许宿主细胞和细菌细胞之间形成复杂的系统,使它们能够进行有效的交流,增强细胞间的相互作用,使它们能够更好地共同服务宿主。
通过内共生学,科学家们可以分析不同微生物之间的关系,学习它们之间的功能影响,从而帮助我们更好地认识真核生物内部的整个系统。
真核细胞内共生学说的发展直接从宿主微生物共聚关系下手,此外,它还考虑了宿主和细菌之间的不同类型的关系,例如寄生、寄生、寄主共用营养、促进寄主增长和免疫反应等,以及宿主和细菌之间的元基因相互影响。
此外,它还考虑到宿主与微生物之间的外因干扰,这些外部因素可能会影响宿主的发育过程和细胞内共生关系,从而影响真核细胞内部的生命过程。
因此,真核细胞内部有一个复杂而动态的共生系统,它涉及到宿主和微生物之间的相互作用关系和元基因的影响,如果能够更好地理解这个系统,可能会更有效地利用这个共生系统来调节宿主细胞内部环境,改善宿主的健康状况,促进宿主的发育过程,从而增强宿主本身的保护性体系。
内共生学说的起源和发展
记得前段时间在准备期中考试时,曾经看到这么一段有意思的话“这些事实都使人设想,真核细胞中的线粒体是由侵入细胞或被细胞吞入的某种细胞经过漫长的岁月演变而来的。
”这确实是对我这个从小对达尔文进化理论深信不移的人产生了很多疑惑。
然而,很多证据都表明线粒体并非真核细胞自生发展而来:
(1). 线粒体含有DNA,这些DNA与细胞核中的很不同,却类似细菌的环状DNA。
(2). 线粒体具有和真核宿主细胞不同的遗传密码,这些密码与细菌和古菌中的很类似。
(3). 线粒体被两层或更多的膜所包被,其中最里面一层的成分与细胞中其它膜的都不同,而更接近于原核生物的细胞膜。
(4). 细胞器的大小与细菌相当。
(5). 细胞器的核糖体和细菌相似,细菌的核糖体是70S,线粒体的核糖体是55S。
在查阅这些资料时,一个对我来说
完全陌生的词汇被反复提及——内共生
学说(endosymbiotic
hypothesis;endosymbiont theory)。
内共生
学说是关于真核生物细胞中的细胞器,
线粒体和叶绿体起源的学说。
根据这个
学说,它们起源于内共生于真核生物细
胞中的原核生物。
这个理论认为线粒体
起源于好氧性细菌(很可能是接近于立
克次体的变形菌门细菌)。
线粒体祖先原
线粒体(一种可进行三羧酸循环和电子
传递的革兰氏阴性菌)被原始真核生物
吞噬后与宿主间形成共生关系。
在共生
关系中,对共生体和宿主都有好处:原线
粒体可从宿主处获得更多的营养,而宿主可借用原线粒体具有的氧化分解功能获得更多的能量。
这个理论的证据非常完整,目前已经被广泛接受。
这个学说的确立也可以说是一波三折。
最早提出这些观点的是一群俄国的科学家。
安德雷.谢尔盖耶维奇.法明茨恩(1835-1918)尝试从植物中分离叶绿体并使其生长。
康斯坦丁.谢尔盖耶维奇.梅里日可夫斯基(1855-1921)发展了“双原生质”(twoplasm)理论(细胞内的细胞),宣称叶绿体起源于蓝绿藻。
波利斯.米哈伊洛维奇.库佐-波利延斯基(Boris M. 1890-1957)则认为细胞的游动性(motility)源于共生。
这三位当时在俄国学术界享有很高的声望。
然而西方主流科学界对于他们的这些想法并不以为然,第一位对这种学说进行发展的是美国人伊万.沃琳(Ivan E. Wallin)。
上个世纪20年代,他在哥伦比亚大学做“推测细胞成分”的实验时,提出叶绿体和线粒体起源于共生的细菌。
他认为细胞质中的叶绿体和线粒体都是外来的,是曾经独立生活的原核细胞。
而这个提法却为其引来了同行的讥讽和排斥。
已过不惑之年的沃琳被迫放弃了关于共生的研究,移居丹佛。
然而,着人们发现它们含有DNA,这些想法被Henry Ris重新提出。
内共生假说被琳·马古利斯所普及。
1967年的下半年,琳.马古利斯以林恩.萨根的名字,在《理论生物学杂志》上发表了《有丝分裂细胞的起源》(《Origin of Eukaryotic Cells》)一文,完整阐述了她对于“内共生起源”的观点。
1969年,她将自己的“内共生起源”观点进行了整理并且以自己的名字琳.马古利斯出版了专著《真核
细胞的起源》,吸引了更大量的科学家投入此领域进行研究,获
得了大量的证据,仅仅10几年的时间,这个“内共生起源假说”
就被当作“内共生起源学说”写进了中学生物学课本。
在1981年的
《细胞进化中的共生》中,她认为真核细胞起源于相互作用的个
体组成的群落,也包括内共生的螺旋体形成了真核生物的鞭毛和
纤毛。
后一种想法没有被广泛接受,因为鞭毛缺乏DNA,在结构
之外和原核生物也没有明显相似性。
根据Margulis和
Sagan(1996),“生命并不是通过战斗,而是通过协作占据整个全
球的”,而达尔文关于进化由竞争驱动的想法是不完善的。
然而
也有人认为内共生更像奴隶制而非互利共生。
这一假说由于证据充分,已被越来越多的人所接受。
其主要证据为:
(1).共生是生物界的普遍现象,例如根瘤菌与豆科植物的共生关系,蓝藻或绿藻与真菌共生形成地衣等。
有一种草履虫(Paramoeciumbursaria),其体内有小的藻类与之共生,并能进行光合作用;过去说澳洲白蚁消化道内生活着一种所谓混毛虫(Mixotricha paradoxa),实际由两种螺旋体、两种真细菌和一种纤毛虫组成,它们能分泌有关的酶,消化纤维素。
特别是近年发现的灰孢藻(Glancocystis),它本身并无叶绿素,但有许多叶蓝小体(cyanella)生活在体内,进行光合作用制造食物。
这种共生关系看来建立不很久,因为叶蓝小体在细胞内还不大固定。
灰孢藻的发现是对“内共生假说”的有力支持。
(2).叶绿体和线粒体都有其独特的DNA,可以自行复制,不完全受核DNA的控制。
线粒体和叶绿体的DNA同细胞核的DNA有很大差别,但同细菌和蓝藻的DNA却很相似。
蓝藻的核糖体RNA(rRNA)不仅可以与蓝藻本身的DNA杂交,而且还可与眼虫叶绿体的DNA杂交,这些都说明它们之间的同源性。
(3).线粒体和叶绿体都有自己特殊的蛋白质合成系统,不受核的合成系统的控制。
原核生物的核糖体由30S和50S两个亚基组成,真核生物的核糖体由40S和60S两个亚基组成。
线粒体和叶绿体的核糖体分别与细菌和蓝藻的一致,也是由30S和50S两个亚基组成,这说明细菌和线粒体、蓝藻和叶绿体是同源的。
抗生素可以抑制细菌和蓝藻的生长,也可以抑制真核生物中的线粒体和叶绿体的作用,这也说明线粒体与细菌、叶绿体与蓝藻是同源的。
(4).线粒体、叶绿体的内、外膜有显著差异,内、外膜之间充满了液体。
研究发现,它们内、外膜的化学成分是不同的。
外膜与宿主的膜比较一致,特别是和内质网膜很相似;内膜则分别同细菌和蓝藻的膜相似。
但是任何学说都不是完美的,内共生学说也不例外,仍然有很多问题是内共生学说所解决不了的:
(1).内共生过程中,拥有先进的氧化代谢途径的好氧细菌无疑应该占优势,但按内共生假说,好氧细菌反而逐步丧失了独立自主性并将其遗传信息成批地转移到了宿主细胞中,这种观点不符合进化论思想。
(2).好氧细菌据估计发生在15亿年以前(甚至更晚一些),但化石提供的证据说明蓝藻大约在22—27亿年以前就形成了。
那么,为什么蓝藻开始共生的时间却远比好氧细菌进行共生的时间迟得多呢?
(3).细菌的细胞色素a十a3不能或极缓慢氧化哺乳动物的细胞色素c,反之,哺乳动物细胞色素a十a3也不能作用于细菌的细胞色素c。
因此,原核细胞的侵入无助于真核细胞的呼吸作用,起不到彼此互利的共生作用。
(4).在叶绿体基因与线粒体基因中发现内含子是惊人的,因为细菌基因中没有发现内含子。
这种内含子的来源及其对生物的效应目前没有得到很好的解释。
(5).线粒体含有肌动蛋白—肌球蛋白系统,但细菌和蓝藻中没有这种系统,也没有相
应的运动能力。
(6).内共生假说没有很好解释细胞核的进化。