DNA复制、引物、酶及蛋白质
- 格式:doc
- 大小:45.00 KB
- 文档页数:14


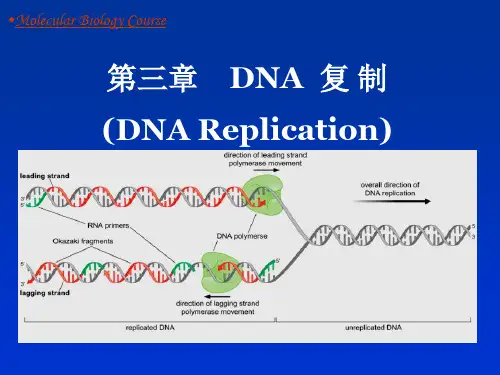

一、半保留复制(semi-conservation replication)(一)证据:1.用氮15标记大肠杆菌DNA,然后在氮-14中培养,新形成的DNA是杂合双链,即双链中一条是重链(约重1%),一条是轻链。
第二代则有一半全是轻链,一半是杂合双链。
2.大肠杆菌DNA在用氚标记的胸苷复制近两代,放射自显影,未复制部分银密度低,由一条放射链和一条非放射链组成;已复制部分有一条双链是放射的,一条双链有一半是放射的。
这证明大肠杆菌DNA是环状分子,以半保留方式复制。
(二)特点:子代保留一条亲代链,而不是将它分解。
这说明DNA是相对稳定的。
双螺旋DNA(或RNA)是所有已知基因的复制形式。
二、复制的起点和单位(一)基因组能独立进行复制的单位称为复制子。
原核生物是单复制子,真核生物是多复制子。
每个复制子有起点。
通过测定基因出现的频率可以确定起点位置,距离起点越近的基因出现的频率越高。
起点有发动复制的序列,也有决定拷贝数的序列。
起点的结构是很保守的。
(二)复制终止点:已发现Ecoli的与复制终止有关位点,其中含有23bp的保守序列,由tus蛋白与此位点结合参与复制的终止。
真核生物中似乎没有复制终止点。
(三)复制多数是双向、对称的,但也有例外。
通过放射自显影可以判断复制是双向还是单向:先在低放射性培养基中起始复制,再转移到高放射性培养基中,如是双向复制,其放射自显影图是中间银密度低;单向复制则为一端低。
(四)单向复制有一些特殊方式:1.滚动环:噬菌体φX174DNA是环状单链分子,复制时先形成双链,再将正链切开,将5’连接在细胞膜上,从3’延长,滚动合成出新的正链。
2.取代环:线粒体DNA复制时是高度不对称的,一条链先复制,另一条链保持单链而被取代,呈D环形状。
这是因为两条链的复制起点不同,另一条链的起点露出才能复制。
三、有关的酶(一)反应特点:1.以四种dNTP为底物2.需要模板指导3.需要有引物3’-羟基存在4.DNA链的生长方向是5’-3’5.产物DNA的性质与模板相同(二)大肠杆菌DNA聚合酶1.DNA聚合酶I:单链球状蛋白,含锌。

原核生物与真核生物DNA复制共同得特点:1底物成分:亲代DNA分子为模板,四种脱氧三磷酸核苷(dNTP)为底物,多种酶及蛋白质:DNA拓扑异构酶、DNA解链酶、单链结合蛋白、引物酶、DNA聚合酶、RNA酶以及DNA连接酶等;2过程:分为起始、延伸、终止三个过程;3聚合方向:5'→3';4化学键: 3',5'磷酸二酯键;5遵从碱基互补配对规律;6一般为双向复制、半保留复制、半不连续复制。
原核生物与真核生物DNA复制不同得特点:1真核生物为线性DNA,具有多个复制起始位点,形成多个复制叉,DNA聚合酶得移动速度较原核生物慢。
原核生物为一般为环形DNA,具有单一复制起始位点。
2真核生物DNA复制只发生在细胞周期得S期,一次复制开始后在完成前不再进行复制,原核生物多重复制同时进行。
3真核生物复制子大小不一且并不同步。
4原核生物有9-mer与13-mer得重复序列构成得复制起始位点,而真核生物得复制起始位点无固定形式。
5真核生物有五种DNA聚合酶,需要Mg+。
主要复制酶为DNA聚合酶δ(ε),引物由DNA聚合酶α合成。
原核生物只有三种,主要复制酶为DNA聚合酶III。
6真核生物末端靠端粒酶补齐,而原核生物以多联体得形式补齐。
7真核生物冈崎片段间得RNA引物由核酸外切酶MF1去除,而原核生物冈崎片段由DNA聚合酶I去除。
8真核生物DNA聚合酶γ负责线粒体DNA合成。
9真核生物DNA聚合酶δ得高前进能力来自于RF-C蛋白与PCNA 蛋白得互相作用。
原核生物DNA聚合酶III得前进能力来自与γ复合体(夹钳装载机)与β亚基二聚体(β夹钳)得相互作用。
10原核生物得聚合酶没有5→3外切酶活性,需要一种FEN1得蛋白切除5端引物,原核生物DNA聚合酶工具有5→3外切酶活性。
11原核得DNA Pol─Ⅱ复制时形成二聚体复合物,而真核生物得聚合酶保持分离状态。
原核生物与真核生物基因信息传递过程中得差异。



原核生物与真核生物DNA复制共同的特点:1底物成分:亲代DNA分子为模板,四种脱氧三磷酸核苷(dNTP)为底物,多种酶及蛋白质:DNA拓扑异构酶、DNA解链酶、单链结合蛋白、引物酶、 DNA聚合酶、RNA 酶以及DNA连接酶等;2过程:分为起始、延伸、终止三个过程;3聚合方向:5'→3';4化学键: 3',5'磷酸二酯键;5遵从碱基互补配对规律;6一般为双向复制、半保留复制、半不连续复制。
原核生物与真核生物DNA复制不同的特点:1真核生物为线性DNA,具有多个复制起始位点,形成多个复制叉,DNA聚合酶的移动速度较原核生物慢。
原核生物为一般为环形DNA,具有单一复制起始位点。
2真核生物DNA复制只发生在细胞周期的S期,一次复制开始后在完成前不再进行复制,原核生物多重复制同时进行。
3真核生物复制子大小不一且并不同步。
4原核生物有9-mer和13-mer的重复序列构成的复制起始位点,而真核生物的复制起始位点无固定形式。
5真核生物有五种DNA聚合酶,需要Mg+。
主要复制酶为DNA聚合酶δ(ε),引物由DNA聚合酶α合成。
原核生物只有三种,主要复制酶为DNA聚合酶III。
6真核生物末端靠端粒酶补齐,而原核生物以多联体的形式补齐。
7真核生物冈崎片段间的RNA引物由核酸外切酶MF1去除,而原核生物冈崎片段由DNA聚合酶I去除。
8真核生物DNA聚合酶γ负责线粒体DNA合成。
9真核生物DNA聚合酶δ的高前进能力来自于RF-C蛋白与PCNA蛋白的互相作用。
原核生物DNA聚合酶III的前进能力来自与γ复合体(夹钳装载机)与β亚基二聚体(β夹钳)的相互作用。
10原核生物的聚合酶没有5→3外切酶活性,需要一种FEN1的蛋白切除5端引物,原核生物DNA聚合酶工具有5→3外切酶活性。
11原核的DNA Pol─Ⅱ复制时形成二聚体复合物,而真核生物的聚合酶保持分离状态。
原核生物与真核生物基因信息传递过程中的差异1. DNA的复制。

原核生物dna复制过程需要的酶和蛋白质以原核生物DNA复制过程需要的酶和蛋白质为标题,本文将详细介绍在原核生物中DNA复制的过程以及其中涉及到的关键酶和蛋白质。
DNA复制是细胞分裂过程中最为重要的一步,它确保了新生的细胞拥有与母细胞完全相同的遗传信息。
在原核生物中,DNA复制是一个复杂而精确的过程,需要多种酶和蛋白质的协同作用。
DNA复制的第一步是DNA的解旋,这由解旋酶(helicase)完成。
解旋酶能够结合到DNA的双链上,并将其解开,形成两条单链。
解旋酶在DNA复制过程中起到了“拆线员”的作用,为下一步的复制提供了单链DNA。
接下来,DNA复制酶(DNA polymerase)开始介入。
DNA复制酶是一类能够将新的DNA链合成到已有的DNA模板链上的酶。
在原核生物中,有多种类型的DNA复制酶参与DNA复制的不同阶段。
其中,DNA polymerase III是最主要的复制酶,它能够快速而准确地合成新的DNA链。
此外,DNA polymerase I在DNA复制的后期起到了修复和整理DNA的作用。
DNA复制的过程中还需要DNA结合蛋白(SSB),它们能够结合到DNA的单链上,防止DNA链的重新结合和降解。
DNA结合蛋白在DNA复制过程中起到了“保护者”的作用,确保了DNA的完整性和稳定性。
在DNA复制过程中,还有一种酶称为DNA拼接酶(DNA ligase)。
DNA拼接酶能够将DNA链中的断裂连接起来,使DNA形成一个完整的双链。
DNA拼接酶在DNA复制过程中起到了“粘合剂”的作用,保证了新合成的DNA链的连续性。
除了上述酶和蛋白质外,DNA复制还需要DNA引物(primer)和DNA引物合成酶(primase)。
DNA引物是一段短的RNA或DNA 链,它能够提供一个起始点,使DNA复制酶能够开始合成新的DNA链。
DNA引物合成酶能够合成DNA引物,为DNA复制提供必要的起始物质。
在DNA复制过程中,还有其他辅助蛋白质参与其中,如DNA修复酶、拓扑异构酶等。
第十二章DNA的复制和修复一:填空题1.参与DNA复制的主要酶和蛋白质包括________________、________________、________________、________________、________________、________________和________________。
2.DNA复制的方向是从________________端到________________端展开。
3.大肠杆菌在DNA复制过程中切除RNA引物的酶是________________,而真核细胞DNA复制过程中切除RNA 引物的酶是________________或________________。
4.大肠杆菌染色体DNA复制的起始区被称为________________,酵母细胞染色体DNA复制的起始区被称为________________,两者都富含________________碱基对,这将有利于________________过程。
5.大肠杆菌DNA连接酶使用________________能源物质,T4噬菌体DNA连接酶使用________________作为能源物质。
6.________________和________________酶的缺乏可导致大肠杆菌体内冈崎片段的堆积。
7.体内DNA复制主要使用________________作为引物,而在体外进行PCR扩增时使用________________作为引物。
8.使用________________酶或________________酶可将大肠杆菌DNA聚合酶Ⅰ水解成大小两个片段,其中大片段被称为________________酶,它保留________________和________________酶的活性,小片段则保留了________________酶的活性。
9.参与大肠杆菌DNA复制的主要聚合酶是________________,该酶在复制体上组装成________________二聚体,分别负责________________链和________________链的合成,已有证据表明后随链的模板在复制中不断形成________________结构。
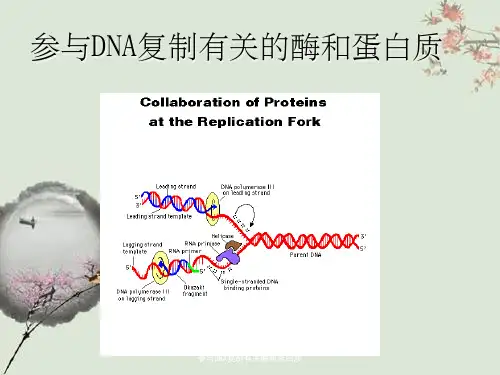
DNA复制起始引发体的形成及所参与的酶和蛋白质、DNA复制过程一、DNA复制起始引发体的形成及所参与的酶和蛋白质DNA复制起始一共涉及到DnaA(复制起始因子,识别OriC序列)、DnaB(DNA解链酶)、DnaC(召唤DnaB到复制叉)、HU(结合DNA使之弯曲)、引物合成酶、单链DNA结合蛋白、RNA聚合酶、DNA旋转酶、Dam甲基化酶,一共是9种重要的酶或蛋白质,其中DnaA、DnaB、引物合成酶、单链DNA结合蛋白、Dam甲基化酶非常重要。
DNA复制时,往往先由RNA聚合酶在DNA模板上合成一段RNA引物,再由聚合酶从RNA引物3’端开始合成新的DNA链。
对于前导链来说,这一引发过程比较简单,只要有一段RNA引物,DNA聚合酶就能以此为起点,一直合成下去。
对于后随链,引发过程较为复杂,需要多种蛋白质和酶参与。
后随链的引发过程由引发体来完成。
引发体由6种蛋白质构成,预引体或引体前体把这6种蛋白质结合在一起并和引发酶或引物过程酶进一步组装形成引发体。
引发体似火车头一样在后随链分叉的方向前进,并在模板上断断续续的引发生成滞后链的引物RNA短链,再由DNA聚合酶 III 作用合成DNA,直至遇到下一个引物或冈崎片段为止。
由RNA酶H降解RNA引物并由DNA聚合酶 I 将缺口补齐,再由DNA连接酶将每两个冈崎片段连在一起形成大分子DNA.。
1.解链酶(helicase,unwinding enzyme)复制叉的形成则是由多种蛋白质及酶参与的较复杂的复制过程。
在DNA不连续复制过程中,结合于复制叉前面,在起始点处解开双链,反应是在解链酶的催化下进行的。
解链酶有ATP酶活性的酶,两种活性相互偶联,通过水解ATP提供解链的能量。
解链酶的作用就是打开DNA双链之间的氢键。
解链酶分解ATP的活性依赖于单链DNA的存在。
如果双链DNA中有单链末端或切口,则DNA解链酶可以首先结合在这一部分,然后逐步向双链方向移动。
★目前已知的DNA聚合酶都只能延长已存在的DNA链,而不能从头合成DNA链。
★凡能从多核苷酸链的末端开始水解核酸的酶称为核酸外切酶,凡能从多核苷酸链中间开始水解核酸的酶称为核酸内切酶。
而能识别特定的核苷酸顺序,并从特定位点水解核酸的内切酶称为限制性核酸内切酶(限制酶)。
★起始位点:真核生物每条染色体上面可以有多处,而原核生物已有一个起始点。
真核生物的染色体在全部完成复制之前,各个起始点上DNA的复制不能再开始,而在快速生长的原核生物中,复制起点上可以连续开始新的DNA复制,表现为虽只有一个起始位点,但有多个复制叉。
★DNA半保留复制:(semiconservative replication)DNA在复制过程中,每条链分别作为模板合成新链,产生互补的两条链。
这样新形成的两个DNA分子与原来的DNA分子的碱基顺序是完全一样的。
因此,每个子代分子的一条链来自亲代DNA,另一条链则是新合成的,这种复制方式即为semiconservation replication。
由Watson and Crick 提出,由Meselson and Stahl通过经15N标记3个世代的大肠杆菌DNA实验得以验证,★DNA半不连续复制:(semi-discontinuous replication):DNA复制过程中,前导链的复制是连续的,而后随链的复制是中断的、不连续的。
★冈崎片段(Okazaki fragment):是DNA半不连续复制中产生的长度为1000~2000个碱基的短的DNA片段,能被连接形成一条完整的DNA链。
★复制子(replicon):单独复制的一个DNA单元被称为一个复制子,它是一个可移动的单元。
一个复制子在任何细胞周期只复制一次。
★复制叉(replication fork):复制时,双链DNA要解开成两股链分别进行DNA结合,所以,复制起点呈叉子形状,被称为复制叉。
★引发酶(primase):是依赖于DNA的RNA聚合酶,其功能是在DNA复制过程中合成RNA引物。
DNA复制起始引发体的形成及所参与的酶和蛋白质、DNA复制过程一、DNA复制起始引发体的形成及所参与的酶和蛋白质DNA复制起始一共涉及到DnaA(复制起始因子,识别OriC序列)、DnaB(DNA解链酶)、DnaC(召唤DnaB到复制叉)、HU(结合DNA使之弯曲)、引物合成酶、单链DNA结合蛋白、RNA聚合酶、DNA旋转酶、Dam甲基化酶,一共是9种重要的酶或蛋白质,其中DnaA、DnaB、引物合成酶、单链DNA结合蛋白、Dam甲基化酶非常重要。
DNA复制时,往往先由RNA聚合酶在DNA模板上合成一段RNA引物,再由聚合酶从RNA引物3’端开始合成新的DNA链。
对于前导链来说,这一引发过程比较简单,只要有一段RNA引物,DNA聚合酶就能以此为起点,一直合成下去。
对于后随链,引发过程较为复杂,需要多种蛋白质和酶参与。
后随链的引发过程由引发体来完成。
引发体由6种蛋白质构成,预引体或引体前体把这6种蛋白质结合在一起并和引发酶或引物过程酶进一步组装形成引发体。
引发体似火车头一样在后随链分叉的方向前进,并在模板上断断续续的引发生成滞后链的引物RNA短链,再由DNA聚合酶 III 作用合成DNA,直至遇到下一个引物或冈崎片段为止。
由RNA酶H降解RNA引物并由DNA聚合酶 I 将缺口补齐,再由DNA连接酶将每两个冈崎片段连在一起形成大分子DNA.。
1.解链酶(helicase,unwinding enzyme)复制叉的形成则是由多种蛋白质及酶参与的较复杂的复制过程。
在DNA不连续复制过程中,结合于复制叉前面,在起始点处解开双链,反应是在解链酶的催化下进行的。
解链酶有ATP酶活性的酶,两种活性相互偶联,通过水解ATP提供解链的能量。
解链酶的作用就是打开DNA双链之间的氢键。
解链酶分解ATP的活性依赖于单链DNA的存在。
如果双链DNA中有单链末端或切口,则DNA解链酶可以首先结合在这一部分,然后逐步向双链方向移动。
复制时,大部分DNA解旋酶可沿滞后模板的5’—〉3’方向并随着复制叉的前进而移动,只有个别解旋酶(Rep蛋白)是沿着3’—〉5’方向移动的。
故推测Rep蛋白和特定DNA解链酶是分别在DNA 的两条母链上协同作用以解开双链DNA。
大肠杆菌中DnaB蛋白就有解链酶活性,与随从链的模板DNA结合,沿5′→3′方向移动,还有一种叫做Rep蛋白和前导链的模板DNA结合沿3′→5′方向移动。
2.单链结合蛋白(single strand binding proteins,SSBP)ssbDNA蛋白是较牢固的结合在单链DNA上的蛋白质。
原核生物ssbDNA蛋白与DNA结合时表现出协同效应:若第1个ssbDNA蛋白结合到DNA上去能力为1,第2个的结合能力可高达103;真核生物细胞中的ssbDNA蛋白与单链DNA结合时则不表现上述效应。
ssbDNA蛋白的作用是保证解旋酶解开的单链在复制完成前能保持单链结构,它以四聚体的形式存在于复制叉处,待单链复制后才脱下来,重新循环。
所以,ssbDNA蛋白只保持单链的存在,不起解旋作用。
ssbDNA蛋白稳定解开的单链,保证此局部不会恢复成双链。
它与解开的单链DNA结合,使其稳定不会再度螺旋化并且避免核酸内切酶对单链DNA的水解,保证了单链DNA做为模板时的伸展状态,SSBP可以重复利用(图)。
SSBP与DNA单链结合,既防止核酸水解酶的作用,又避免解开的单链DNA重新缔合形成双链,从而保持一种伸展状态,以保证复制顺利进行。
在E.coli中SSB为四聚体,对单链DNA有很高的亲和性,但对双链DNA和RNA没有亲和力。
它们与DNA结合时有协同作用。
图大肠杆菌DNA复制叉中复制过程简图3、DNA旋转酶(DNA gyrase )是一个由两个GraA和两个GraB亚基构成的四聚体蛋白,DNA旋转酶以其可以利用ATP结合和水解的能量来介导负超螺旋到松弛的共价闭合DNA而闻名于所有拓扑异构酶。
它是DNA拓扑异构酶Ⅱ中的一种亚类,其主要功能为引入负超螺旋,在DNA 复制中起十分重要的作用。
迄今为止,只有在原核生物中才发现DNA旋转酶。
作用机理: DNA复制时解链产生正螺旋,阻碍复制体的前进,DNA旋转酶通过引起瞬间DNA双链的断裂和重新连接,以改变DNA的拓扑状态,引入负超螺旋。
4.引发体的形成:DNA复制起始的关健步骤是前导链DNA的合成,一旦前导链DNA的聚合作用开始,随从链DNA的合成也随着开始。
由于前导链的合成是连续进行的,所以它的起始相对简单,而随从链的合成是不连续进行的,所以引发阶段比较复杂。
大肠杆菌的引发前体由Dna B. Dna C和单链结合蛋白组成。
(1)引物酶(primase)它是一种特殊的RNA聚合酶,可催化短片段RNA的合成。
这种短RNA片段一般十几个至数十个核苷酸不等,它们在DNA复制起始处做为引物。
RNA引物的3′桹H末端提供了由DNA聚合酶催化形成DNA分子第一个磷酸二酯键的位置。
催化RNA引物合成的酶叫引发酶(primerase)。
引发酶识别DNA 单链模板特异序列,以核糖核苷三磷酸为底物合成寡聚核苷酸,产生3′-OH,为DNA聚合酶起始磷酸二酯键提供条件。
引物与典型的RNA不同,他们在合成以后并不与模板分离,而是以氢键与模板结合。
实验表明,在细菌中,前导链的引物和滞后链中前体片断的引物在合成时需要不同的条件。
滞后链前体片断引物的合成是由引发酶催化的,而引发酶对利福平(rifampicin)不敏感,因此不受利福平抑制;前导链引物的合成是由RNA聚合酶催化的,而RNA聚合酶对利福平十分敏感,所以受利福平强烈抑制。
(2)引发体(primosome)高度解链的模板DNA与多种蛋白质因子形成的引发前体促进引物酶结合上来,共同形成引发体,引发体主要在DNA随从链上开始,它连续地与引物酶结合并解离,从而在不同部位引导引物酶催化合成RNA引物,在引物RNA的3′桹H末端接下去合成DNA片段,这就是随从链不连续合成的开始。
5、DNA聚合酶1)DNA聚合酶IDNA聚合酶I最初由Kornberg于1955年在大肠杆菌中发现的,纯化的酶是一条相对分子量为109KD的多肽链。
用枯草杆菌蛋白酶处理可以将DNA聚合酶I水解为一大一小两个片段。
较大的C末端片断分子量为68KD,叫klenow片断,常用做体外合成DNA 的工具酶;具有5′→3′的聚合酶活性,可以将脱氧核苷三磷酸加到DNA链的3′-OH上,形成3′,5′-磷酸二酯键,这个活性需要DNA模板和引物的存在。
klenow片断还具有3′→5′的外切酶活性,可以从3′端将DNA链水解。
当正确的核苷三磷酸进入正在合成的链的末端,酶就向前移动,一旦发生错误,酶就退回,将错误的碱基切除,这是一种校对(proofreading)功能。
较小的N末端片断分子量为35 KD,具有5′→3′的外切酶活性,在体内功能是切除小的DNA片断,包括切除RNA引物。
尽管DNA聚合酶I活性很高,但它们的主要功能是用于修复体内DNA双螺旋区中的单链区,这些单链区是在DNA复制时或DNA受损伤后留下的。
DNA聚合酶I的5′→3′核酸外切酶活性从5′端切除DNA受损伤的部位,DNA聚合酶I的5′→3′聚合酶活性随即开始工作,使DNA链向3′端延伸,这称为切口平移(nick translation)。
在DNA复制中,RNA引物的切除实际上就是切口平移;DNA的损伤修复中也存在切口平移。
在体外,可以用切口平移来制备高放射性活性的DNA 探针。
2)DNA聚合酶ⅢDNA聚合酶Ⅲ全酶以异源复合体发挥功能。
α亚基、ε亚基和θ亚基相连组成核心酶(core enzyme),每种亚基两个,形成两个催化合成DNA的活性中心。
α亚基具有5′→3′的聚合酶活性,并具有5′→3′的外切酶活性;ε亚基具有3′→5′的外切酶活性,这对校正功能十分重要。
核心酶在τ亚基存在下,形成二聚体。
其他亚基的添加促进二聚化和增强了酶活性(τ亚基起着促使核心酶二聚化的作用)。
DNA聚合酶Ⅲ具有持续合成能力,β亚基在二聚体形成中发挥重要的作用。
β亚基的功能犹如夹子、两个β亚基夹住DNA分子并可以向前滑动,使聚合酶在完成复制前不再脱离DNA,从而提高了酶的持续合成能力(通过它形成一个环使DNA聚合酶Ⅲ的核心酶附着在DNA模板上,并可以沿单链DNA滑动而使DNA的复制过程程序化)。
β亚基的定位能防止核心酶在DNA复制过程中从模板上掉下来,如果没有β亚基,核心酶只能合成大约20个核苷酸就要脱离模板。
除了β亚基以外,γ亚基是一种依赖于DNA的ATP酶,两个γ亚基与另4个亚基(δ亚基、δ′亚基、χ亚基、ψ亚基)构成γ复合物,其主要功能时帮助β亚基夹住DNA,故称夹子装置器(clamp loader)。
β亚基在DNA上定位和解离都需要γ复合物介导。
完整全酶为不对称二聚体结构。
当聚合酶遇到逆向模板上前面合成的冈崎片断时,γ复合物的存在允许β亚基从模板上解离。
这样当核心酶合成了冈崎片断,将从后滞链的全酶中释放出来,与下一个由DNA引发酶提供的模板引物的起始前复合物结合。
在半不连续复制过程重,同一个DNA聚合酶Ⅲ分子可以同时合成前导链和滞后链。
更确切地讲,在前导链和滞后链的生长点,核苷酸的增加同时进行。
如前所述,DNA聚合酶Ⅲ有两个DNA合成催化中心,很可能前导链和滞后链各利用一个催化中心合成DNA。
滞后链的模板链围绕一个催化中心折叠成环状结构,使滞后链的方向颠倒(生化方向不变)。
6、DNA连接酶DNA连接酶催化双股链内相邻单链切口的3′-OH与5′- P酰基形成磷酸酯键。
酶的活化? NAD+或ATP中的腺苷酰基(AMP)与酶活性中心的赖氨酸残基的ε-NH2以磷酰胺键结合,形成共价中间体酶- AMP;腺苷酰化DNA? 酶- AMP将腺苷酰基(AMP)转移给DNA切口处的5′磷酰基团,以焦磷酸的形式活化,形成AMP-P-DNA。
亲核攻击完成DNA连接? 通过相邻DNA的3′-OH对活化的P原子进行亲核攻击,生成3′,5′-磷酸二酯键,同时释放出AMP。
二、DNA复制过程真核、原核的复制大致可以分为复制的引发,DNA链的延伸和DNA复制的终止三个阶段。
起始:DNA解旋酶在局部展开双螺旋结构的DNA分子为单链,引物酶辨认起始位点,以解开的一段DNA为模板,按照5'到3'方向合成RNA短链。
形成RNA引物。
DNA片段的生成:在引物提供了3'-OH末端的基础上,DNA聚合酶催化DNA的两条链同时进行复制过程,由于复制过程只能由5'->3'方向合成,因此一条链能够连续合成,另一条链分段合成,其中每一段短链成为冈崎片段(Okazaki fragments)(由日本冈崎夫妇发现并命名)。