染色体组分析法与小麦的合成
- 格式:pdf
- 大小:867.01 KB
- 文档页数:37

《作物育种学》课程考试样卷1一、名词解释(每小题2 分,共10 分)DUS,选择育种,聚合杂交,杂种优势,体细胞杂交二填空题(每空1分共15分)1.现代农业对作物品种性状的要求主要有高产、⑪、⑫、适应性强;2.根据作物品种的繁殖方式、商品种子生产方法、遗传基础、育种特点和利用形式等可将作物品种区分为下列类型⑬、⑭、群体品种和⑮;3.种质资源工作的内容包括收集、保存、⑯、⑰、⑱;4.轮回选择的方法包括群体间改良和⑲;5.配合力是自交系的一种内在属性,受多种基因效应支配,农艺性状好的自交系不一定就是配合力高,只有配合力高的自交系才能产生强优势的杂交种。
试用格子方试验计算SCA和GCA填表(表格在最后)。
三、选择题(每小题 1 分,共11分)1.杂合基因型的作物,自交后代的生活力();⑪衰退⑫增强⑬不变⑭难以确定2.由胚珠或子房壁的二倍体细胞经过有丝分裂而形成胚,和由正常胚囊中的极核发育成的胚乳而形成种子,这种生殖方式称为()。
⑪无孢子生殖⑫二倍体孢子生殖⑬不定胚生殖⑭单性生殖3. 纬度相近的东西地区之间引种比经度相同的南北之间引种成功的可能性()一些。
⑪大⑫小⑬不好确定4.纯系学说认为,在自花授粉作物原始品种群体中选择()⑪有效⑫无效⑬有的人选择有效,有的人选择无效⑭难以确定5.从理论上讲,杂交育种工作中,所用亲本越多越()⑪好⑫不好⑬易行(成本越低)⑭难以育出好品种6.杂种优势的遗传基础是()⑪显性假说⑫显性假说和超显性假说⑬超显性假说⑭纯系学说7.作物诱变育种中,主要处理植物的()⑪植株或植株的局部⑫花粉⑬种子⑭都可以8.普通小麦×硬粒小麦杂交,是()⑪种间杂交⑫属间杂交⑬种内不同类型杂交⑭)亚种间杂交9. 小种分化明显的的病原菌群体,实由若干个毒性有所不同的小种组成,其中比例较小的小种,称为()⑪优势小种⑫次要小种⑬生理小种⑭)毒性小种10.群体改良有各种不同的方法,常用的是()。
⑪不同变异类型的形成⑫作物雄性不育性的利用⑬轮回选择⑭)回交育种11.迄今为止,一切基因工程的载体都是由()来充任。

染色体组判断1.右图是甲、乙两种生物的体细胞内染色体情况示意图,则染色体数与图示相同的甲、乙两种生物体细胞的基因型可依次表示为:A.甲:AaBb 乙:AAaBbbB.甲:AaaaBBbb 乙:AaBBC.甲:AAaaBbbbCCCc 乙:AaaBBbD.甲:AaaBBb 乙:AaaaBbbb染色体组是指:细胞中的一组非同源染色体,在形态和功能上各不相同,携带着控制生物生长发育的全部遗传信息,这样一组染色体,叫做一个染色体组。
对染色体组的判断通常有两种题型:①图示体,即给出某个体体细胞的染色体示意图,通过分析图示判断染色体组数;②基因型题,即给出某个体的基因型,通过基因型判断染色体组数。
本题则将两种题合二为一,是一道很好的习题。
解本题的关键是对上述两种题型的掌握:图示题首先将细胞中的染色体分成同源染色体组,然后数出一组同源染色体中染色体的条数,即为该细胞染色体组的数目,而每一个染色体组所含染色体条数即为同源染色体组数目;基因型题,首先看同种字母,不区分大小写,有几个就意为着细胞中有几个染色体组,每一个染色体组含有的染色体条数则是不同字母的种数。
就本题而言,甲图中的染色体可以分为3组同源染色体,每组同源染色体含有4条染色体,故此细胞中含有3个染色体组,每个染色体组含有4条染色体;乙图中的染色体可以分为2组同源染色体,每组同源染色体含有3条染色体,故此细胞中含有2个染色体组,每个染色体组含有3条染色体;乙图中的染色体可以分为2组同源染色体。
所以甲图对应的基因型应为AAaaBbbbCCCc或类似组成,乙图基因型应为AaaBBb或类似组成。
2.下列细胞中具有三个染色体组的是()①二倍体水稻的受精卵②二倍体水稻的受精极核③普通小麦的花粉细胞④四倍体水稻的胚乳细胞⑤普通小麦的胚乳细胞⑥香蕉的体细胞A.①③④B.②④⑥ C.②③⑥ D.②④⑤答:二倍体水有稻的精子,卵细胞和1个极核中,均是只含有一个染色体组,所以,受精卵含2个染色体组,而受精极核中则含有3个染色体组,普通小麦是六倍体,它的花粉细胞中含有3个染色体组,四倍体水稻的胚乳细胞含有6个染色体组,但普通小麦的胚乳细胞则含有9个染色体组,香蕉是三倍体,其体细胞中含有3个染色体组,所以具有三个染色体组的是②③⑥选C。

普通小麦川农16与圆锥小麦杂交后代分析李伟;陈国跃;魏育明;郑有良【摘要】对小麦川农16与圆锥小麦地方品种杂交后代进行了分析.结果表明,杂种F1均舍有35条染色体,减数分裂时出现多种配对形式,染色体组成上存在差异.F1植株株高和小穗数普遍介于双亲之间,穗长表现出了超亲优势.稳粒数不同组合间存在差异,同时F1正常结实.F2代表现出较大分离,单株问差异较大,同时育性发生分离.采用优异品种回交可以有效控制远缘杂种后代分离.【期刊名称】《种子》【年(卷),期】2008(027)011【总页数】4页(P30-33)【关键词】川农16;圆锥小麦;杂交后代【作者】李伟;陈国跃;魏育明;郑有良【作者单位】四川农业大学小麦研究所,四川,雅安,625014;四川农业大学农学院,四川,雅安,625014;四川农业大学小麦研究所,四川,雅安,625014;四川农业大学小麦研究所,四川,雅安,625014;四川农业大学小麦研究所,四川,雅安,625014【正文语种】中文【中图分类】S512.1+9随着人们生产和生活水平的提高,对小麦育成品种提出了更多更高的要求。
而充分利用野生资源所蕴藏的特征特性,在育成品种中创造出更加丰富的变异类型成为实现目标的有效途径之一。
普通小麦近缘属中含有丰富的可供利用的优异基因资源,Harlan和de Wet(1971)将作物基因资源划分为三级基因源,并针对不同基因源提出了利用策略。
普通小麦的二级基因源是指具有1~2个与普通小麦相同基因组物种,其与普通小麦杂交的难易程度差别较大[1]。
随着育种方法和手段的提高,以及性状筛选技术的改进,对普通小麦各级基因源进行了利用,得到了一大批小麦新品种和新材料,有力地促进了小麦育种的发展。
如利用中间偃麦草育成“龙麦1号”“龙麦2号”、“新曙光6号”等春小麦品种,以及八倍体小偃麦“远中1号”,通过属间杂交育成一批小麦新种质。
如小麦与山羊草10个种的双二倍体,小麦与黑麦、长穗偃麦草、中间偃麦草的双二倍体、附加系、代换/易位系,小麦与冰草的附加系和代换/易位系,小麦与赖草、羊草、鹅观草、纤毛鹅观草等的易位系,还将东方旱麦草、新麦草、华山新麦草与小麦杂交成功[2]。

小麦族染色体组命名本文旨在详细探讨小麦族染色体组的命名方式。
小麦族一共包含7个染色体,他们的命名方式是由来自不同物种的A、B、D三组染色体组成的。
A组染色体由23条染色体组成,其中21条染色体来自物种Triticum monococcum(小麦),2条染色体分别来自Aegilops speltoides(斯芬塔尔小麦)和Aegilops tauschii(穗小麦)。
B组染色体由14条染色体组成,它们都来自于物种Triticum dicoccoides(双花小麦). D组染色体由6条染色体组成,它们来自物种Aegilops tauschii(穗小麦)。
此外,还有一个特殊的染色体,称为U要素,它来自3种物种:Triticum aestivum(小麦)、Triticum durum(小麦)和Triticum turgidum(杂粮小麦)。
小麦族染色体组命名采用国际上普遍采用的“染色体-原物种-编号”格式,A,B和D组染色体分别用“A,B,D”表示,然后是一个括号内的原物种简写名称和编号,例如A1和B2。
U要素中的染色体用“U”表示,然后是一个括号内的3个物种简写名称,按字典序排列,代表这个特殊染色体来自3种物种,例如U(tse tsita)。
因此,为了简洁有效地命名小麦族染色体组,我们采用“染色体-原物种-编号”格式,详细地记录小麦族染色体来自哪些物种,以及哪些物种组成U要素,使得小麦族染色体组的命名变得清晰明了。
除了物种名称,染色体组还有其他常见命名方式。
在一些研究中,小麦族染色体组也可以简单地用字母“T”,“P”和“R”表示。
其中“T”代表Triticum sp.染色体,“P”代表Aegilops speltoides 染色体,“R”代表Aegilops tauschii 染色体。
此外,还有一种常见的拼写法“7A-7B-7D”,以表示小麦族染色体组的完整性和复杂性,其中“7A-7B-7D”分别表示A组、B组和D组染色体的数量。

小麦核型分析实验
实验原理:一个二倍体植物的配子的全套染色体,称为一个染色体组。
各种动植物的体细胞,都有其特定的染色体组型。
染色体组型或称为核型是指染色体组在有丝分裂中期的表型,包括这一组染色体的数目、大小、形态、着丝点位置以及副溢痕、随体的有无等。
染色体组型分析就是对染色体组中的染色体作上述各种形态特征的描述。
实验材料:小麦
实验药品:α-溴代;2%Hcl;卡宝品红;蒸馏水
实验仪器:三角瓶或者其他容器;载玻片;盖玻片;酒精灯;铅笔
实验步骤:
1.种子萌发和取材:小麦种子置于培养皿内湿滤纸上,25℃恒温箱中,根长2-3cm切取。
2.固定:α-溴代固定幼根1-2d。
3.解离:取根尖2个(每个大约0.1-0.2mm)于载玻片上,滴加2%Hcl 2-3滴3min(主要是
进行破壁),3 min后吸干Hcl,再滴几滴蒸馏水保持3min,3min后用吸水纸吸干。
4.染色:在载玻片上滴加卡宝品红染液染色2min,2min后盖上盖玻片,用大拇指垂直往
下压,压好后,用铅笔头进行敲片,边敲边在在酒精灯下烘干。
5.观察:在显微镜下观察,并拍照。

第3课时 生物变异在育种上的应用课标要求 阐明生物变异在育种上的应用。
1.单倍体育种(1)原理:染色体(数目)变异。
(2)过程(3)优点:明显缩短育种年限,且得到纯合二倍体。
(4)缺点:技术复杂。
2.多倍体育种(1)方法:用秋水仙素或低温处理。
(2)处理材料:萌发的种子或幼苗。
(3)原理:染色体(数目)变异。
(4)实例:三倍体无子西瓜的培育①两次传粉⎩⎪⎨⎪⎧第一次传粉:杂交获得三倍体种子第二次传粉:刺激子房发育成果实 ②用秋水仙素处理幼苗后,分生组织分裂产生的茎、叶、花的染色体数目加倍,而未经处理部分(如根部细胞)的染色体数目不变。
③三倍体西瓜无子的原因:三倍体西瓜植株在减数分裂过程中,由于染色体联会紊乱,不能产生正常配子。
3.杂交育种(1)原理:基因重组。
(2)过程①培育杂合子品种选取符合要求的纯种双亲杂交(♀×♂)→F 1(即为所需品种)。
②培育隐性纯合子品种:选取符合要求的双亲杂交(♀×♂)→F 1――→⊗F 2→选出表型符合要求的个体种植并推广。
③培育显性纯合子品种 a .植物:选择具有不同优良性状的亲本杂交,获得F 1→F 1自交→获得F 2→鉴别、选择需要的类型,连续自交至不发生性状分离为止。
b .动物:选择具有不同优良性状的亲本杂交,获得F 1→F 1雌雄个体交配→获得F 2→鉴别、选择需要的类型与隐性类型测交,选择后代不发生性状分离的F 2个体。
(3)优点:操作简便,可以把多个品种的优良性状集中在一起。
(4)缺点:获得新品种的周期长。
4.诱变育种 (1)原理:基因突变。
(2)过程(3)优点①可以提高突变率,在较短时间内获得更多的优良变异类型。
②大幅度地改良某些性状。
(4)缺点:有利变异个体往往不多,需要处理大量材料。
考向一 分析单倍体育种与多倍体育种的应用1.(2022·宁波高三模拟)如图为二倍体玉米花粉培育成植株的过程。
下列有关叙述错误的是( )A.过程①是花药离体培养B.过程②若正常培养,则植株B是单倍体C.过程②若使用秋水仙素处理幼苗使其染色体加倍,则植株B是二倍体纯合子D.若该过程为单倍体育种,则育种原理是基因重组答案D2.一粒小麦(染色体组AA,2n=14)与山羊草(染色体组BB,2n=14)杂交,产生的杂种AB经染色体自然加倍,形成了具有AABB染色体组的四倍体二粒小麦(4n=28)。


小麦的起源小麦源于何处?其祖先是谁?对这类问题很多人都会感到惊异,怎么会想出这类怪问题?小麦不是全世界到处都长吗?其祖先难道不是小麦?看起来这些问题都很简单,但细究起来可有点不那么容易了。
下面我们就来看看日本遗传学家木原均(生于1893年10月21日)是如何来探明这些问题的。
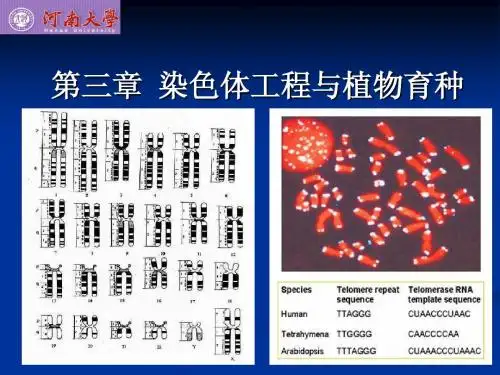
1918年,日本科学家坂村彻确定了小麦正确的染色体数,他发现一粒系小麦(Einkor ngrou p)核内有14条染色体(2n=14),二粒系小麦(Emmerg roup)有28条(2n=28),普通系为42条(2n=42)。
在坂村彻工作的基础上,木原均选用五倍体开始他的研究。
其五倍体是四倍体的波兰小麦(Tritic umpol onicu m,2n=4x=28)和六倍体的斯卑尔脱小麦(T.spelta,2n=6x=42)的杂交种,染色体数为35,其中14条来自波兰小麦(二粒系),21条来自斯卑尔脱小麦(普通系)。
经过研究,木原均提出了“染色体组”的概念,认为在小麦中每个染色体组为7条染色体。
这之后,木原均就不同系的小麦染色体组进行分析,观察其染色体组同源的程度。
他建立了一种方法即观察染色体的配对,视配对的完全程度来判定其是否同源。
在两个染色体组之间,如果新有的染色体都配对,则两个组完全同源;若完全不配对,则非同源;部分染色体配对,即可判定两个组部分同源。
根据这种方法,他判定二倍体(一粒系)、四倍体(二粒系)、六倍体(普通系)的染色体组组成分别为A A、AABB、AABB-DD,此外他还发现四倍体小麦中有一部分种中有新的染色体组G,因此除了先前新知的三个小麦系外,还有第四个系即提莫菲维小麦系(T.timoph eevig roup,AAGG).至此木原均查明栽培小麦的祖先是些微不足道的野生植物。


2024.5生物(考试时间90分钟满分100分)第一部分本部分共15题,每题2分,共30分。
在每题列出的四个选项中,选出最符合题目要求的一项。
1. 细胞在迁移过程中会产生并释放一种单层膜的细胞器——迁移体,其内部含有细胞因子、mRNA等物质。
当迁移体被周围细胞吞噬后,其中的mRNA翻译形成蛋白质,进而改变该细胞的行为。
关于迁移体的推断正确的是()A. 包含四层磷脂分子B. 其膜不属于生物膜系统C. 可能参与细胞间的信息交流D. 其被吞噬依赖于细胞膜的选择透过性【答案】C【解析】【分析】1、生物膜系统包括细胞器膜和细胞膜、核膜等结构。
这些生物膜的组成成分和结构很相似,在结构和功能上紧密联系,进一步体现了细胞内各种结构之间的协调与配合。
2、细胞膜的组成成分主要是蛋白质和脂质。
细胞膜的功能有:将细胞与外界环境分隔开、控制物质进出细胞、进行细胞间的信息交流。
细胞膜的结构特点是具有一定的流动性,功能特点是具有选择透过性。
【详解】A、细胞在迁移过程中会产生并释放一种单层膜的细胞器——迁移体,其膜结构包括两层磷脂分子,A错误;B、迁移体外侧有膜包被,是一种单层膜结构的细胞器,而生物膜系统包括细胞膜、细胞器膜和核膜等,可见,迁移体的膜结构属于细胞的生物膜系统,B错误;C、迁移体内部含有细胞因子、mRNA等物质。
当迁移体被周围细胞吞噬后,其中的mRNA翻译形成蛋白质,进而改变该细胞的行为,这说明迁移体可能参与细胞间的信息交流,C正确;D、当迁移体可被周围细胞吞噬,该过程属于胞吞过程,胞吞过程不依赖于细胞膜的选择透过性实现,D 错误。
故选C。
2. 肺炎克雷伯菌(Kpn)存在于某些人群的肠道中,可通过细胞呼吸不断产生大量乙醇,引起内源性酒精性肝病。
下列叙述正确的是()A. Kpn 在细胞质基质中将丙酮酸转化为乙醇并产生大量ATPB. Kpn 无氧呼吸使有机物中稳定的化学能全部转化为活跃的化学能C. 乳酸菌、酵母菌、Kpn 都可以引起内源性酒精性肝病D. 高糖饮食可能会加重内源性酒精性肝病患者的病情【答案】D【解析】【分析】1、有氧呼吸是指细胞在氧的参与下,通过多种酶的催化作用,把葡萄糖等有机物彻底氧化分解,产生二氧化碳和水,释放能量,生成大量ATP 的过程。
专题(七)变异与进化明考点·析考情考点1.基因突变和基因重组。
2.染色体结构变异和数目变异。
3.生物变异在生产实践中的应用。
4.现代生物进化理论的主要内容。
5.生物进化与生物多样性的形成。
考情1.考查题型:多以选择题呈现。
2.命题趋势:(1)变异常以实例辨析背后原理或以变异为基础考查育种的流程及背后的原理。
(2)进化多侧重基本观点考查及基因频率的计算。
命题点1变异及其育种应用1.突破生物变异的4大问题2.“两看法”界定二倍体、多倍体、单倍体3.界定“三倍体”与“三体”4.几种常考类型产生配子种类及比例5.育种方式及原理辨析(1)诱变育种原理(2)单倍体育种与杂交育种的关系(3)多倍体育种的原理分析6.准确选取育种方案7.细胞癌变的原理和特征8.变异类型的分析推理与实验设计思路(1)显性突变与隐性突变的判断(适于单基因突变)(2)一对基因突变和两对基因突变的判断①两种突变来自一对基因突变(隐性突变)②两种突变来自两对基因突变(隐性突变)(3)基因突变和染色体变异的判断现有一红眼雄果蝇X A Y与一白眼雌果蝇X a X a杂交,子代中出现一只白眼雌果蝇(注:染色体缺失的纯合子致死)。
①观察法:取该果蝇有分裂能力的细胞制成装片,显微镜下观察染色体结构。
若染色体正常,可能是基因突变引起的;反之可能是染色体缺失引起的。
②杂交法:选该白眼雌果蝇与红眼雄果蝇杂交,若杂交子代中雌果蝇数与雄果蝇数比为1∶1,则这只白眼雌果蝇的出现是由于基因突变引起的;若杂交子代中雌果蝇数与雄果蝇数比为2∶1,则这只白眼雌果蝇的出现是染色体缺失造成的。
1.(2023·福建,9)无义突变是指基因中单个碱基替换导致出现终止密码子,肽链合成提前终止。
科研人员成功合成了一种tRNA(sup—tRNA),能帮助A基因第401位碱基发生无义突变的成纤维细胞表达出完整的A蛋白。
该sup—tRNA对其他蛋白的表达影响不大。
过程如图。
⼩麦(异源六倍体)的形成过程⼩麦的六倍体属于异源六倍体,它的六个染⾊体组的写法是AABBDD.究其来源是由⼀种⼆倍体植物AA(A表⽰染⾊体组,AA表⽰这个植物体有两个染⾊体组,每⼀组A中含有7条染⾊体,2N=14)和另⼀种⼆倍体植物BB(B表⽰另⼀个染⾊体组,BB表⽰这个植物⼜不同于上⼀个植物AA的两个染⾊体组,但每⼀组B中也含有7条染⾊体,但与A不同,记做2N=14)减数分裂(N=7)并受精作⽤形成的AB(2N=14),但是,请注意,这⾥的A 染⾊体组和B染⾊体组之间并⽆任何染⾊体能配对在⼀起,减数分裂联会紊乱,所以是不可育的.这时,可能由于环境温度骤变导致纺锤丝没有形成,结果染⾊体加倍⽽得出的AABB.因为AB的形成过程是经过受精作⽤的,所以它不叫单倍体,⽽应该叫异源⼆倍体(所谓异源是指物种来源不同),那么它加倍后就叫异源四倍体了,这⾥⾯A这个染⾊体组⾥所有的染⾊体只能和另⼀组A⾥的所有染⾊体配对,成为同源染⾊体.同理B只能和B配对,既然能配对,当然可以进⾏减数分裂,所以AABB这个个体是可育的.后来这个AABB个体⼜和⼀个⼆倍体DD(D表⽰再⼀个染⾊体组,DD表⽰这个植物⼜不同于上两种植物AA和BB 的两个染⾊体组,但每⼀组D中也含有7条染⾊体,但与A、B中的染⾊体均不同,记做2N=14)相遇,再⼀次经历了上述过程,即:AABB减数分裂形成AB(2N=14)与DD减数分裂形成的D(N=7)精卵结合成为ABD(3N=21),因为有受精作⽤,所以这是个异源三倍体,因为没有可以配对的同源染⾊体,所以不可育.还是环境条件造成染⾊体加倍,形成了AABBDD异源六倍体(6N=42).好了,我们可以看到这个怪物的形成过程,也就知道了它虽然叫做六倍体,但是A和B和D这三组染⾊体组中的染⾊体并不能进⾏配对,所以并不能简单的拿42除以6=7对,⽽只能是42除以2=21对同源染⾊体.下⾯我们⽤遗传图解来表⽰⼀下:P ⼀粒⼩麦 x 拟斯卑尔脱⼭⽺草 AA x BB F1 AB(异源⼆倍体) 环境突变、染⾊体加倍 = AABB ⼆粒⼩麦 ⼆粒⼩麦 x ⽅穗⼭⽺草AABB x DD F2 ABD(异源三倍体) 环境突变、染⾊体加倍 = AABBDD 六倍体普通⼩麦 ⼩麦系由⼭⽺草属、⼴义的冰草属和⼩麦属3个属的种类杂交形成的,染⾊体组为AABBDD是6n=42的异源6倍体植物.所以六倍体⼩麦中有21对同源染⾊体.。
染色体组分析法与小麦的合成验证普通小麦的起源与演化主要内容•小麦的染色体数目统计与倍性分析•染色体组分析法与小麦的染色体组型•验证普通小麦的D染色体组来源:(1)柱穗山羊草与粗山羊草的染色体组分析(2)“人工合成六倍体小麦”与“木原小麦”•验证普通小麦的D染色体组来源:(1)5B“抑制因子”(2)染色体组分析法的局限性“地球的历史是写在它的岩层上,而有机体的历史则是刻在它的染色体上。
”——木原均(Hitoshi Kihara)•通过对小麦属(Triticum)和山羊草属(Aegilops)的研究确立了染色体组(Genome)的概念;•由于染色体组概念的确立,首创了染色体组分析的方法(Genome analysis method);•由于染色体组分析方法的应用,阐明了普通小麦的起源;•用与小麦近缘的几个物种,通过远缘杂交和杂种染色体加倍人工合成普通小麦;小麦的染色体数目统计与倍性分析染色体标本的制备:根尖压片法1. 取材:把小麦种子预先浸泡6h,然后转入垫有湿润滤纸的培养皿中,置25℃温箱中发芽,幼根长至1~2cm,于上午9:00-10:00剪下根尖。
2. 预处理:将剪下的根尖置于0.02%秋水仙碱溶液中,浸泡处理4h。
3. 固定:用水洗净处理过的根尖,经甲醇-冰醋酸(3:1)固定6~12h,再经95%乙醇、85%乙醇各浸泡0.5h,最后转入70%乙醇中,4℃保存备用。
4. 解离和染色:取出根尖,用蒸馏水洗净,放入1mol/L HCl中60℃解离8~10min。
解离后的材料于45%冰醋酸中软化40~50min,之后用蒸馏水冲洗。
用改良苯酚品红染色12~24h后进行压片。
敲好的玻片在显微镜下观察,选择染色体分散好,形态清晰的标本,经冷冻脱片封片法制成永久制片。
染色体数目的统计与倍性分析对染色体标本进行仔细观察,每种麦类植物挑选30~100个分裂中期的细胞进行染色体数目统计,以此分析植物染色体的倍性。
Triticum L. 小麦属染色体组分析法与小麦的染色体组型染色体组分析法Genome analysis method所谓“染色体组分析”,就是将小麦属与其近缘种属进行杂交,根据杂种减数分裂中染色体是否配对作为染色体组之间异同的判断标准,配对(形成二价体),说明同源,属同一染色体组;不配对(形成单价体),说明不同源,则属不同染色体组。
1. 选定亲本,对母本进行人工去雄,套袋,2~3天后授以父本的花粉。
统计杂交的结实率,收获杂种种子后,干燥保存。
2. 将杂种种子在培养皿内于25℃的恒温下发芽,统计杂种发芽率,然后在移栽于盆内。
3. 杂种和亲本孕穗期取处于减数分裂盛期的幼穗用卡诺液II(乙醇:氯仿:醋酸=6:3:1)固定,以醋酸洋红染色压片,观察并统计杂种F1及亲本花粉母细胞减数分裂中期I染色体配对行为。
1. 如果在减数分裂中观察到正常的染色体配对或很高的染色体配对数——染色体同源(homology),则表示亲本种之间有非常近的亲缘关系;2. 如果染色体配对数较低而且染色体之间的联合状况较差——染色体部分同源(homoeology),则表明亲本种之间有一定的亲缘关系;3. 如果杂种中的染色体之间完全不发生联合配对——染色体完全不同源(non-homology),则亲本种之间的亲缘关系很远。
同时,与染色体配对行为的分析相结合,对于杂种的育性,如:花粉育性、结实率等的观察统计,也作为估计亲本种之间亲缘关系的补充指标。
小麦的染色体组型一粒小麦(Triticum momococcum, 2n=2x=14)是同源二倍体,染色体组型可表示为AA;是最原始的小麦。
1. 圆锥小麦(T. trugidum, 2n=4x=28)与一粒小麦同源的为A染色体组,不同源的可表示为B染色体组,则染色体组型为AABB;2. 普通小麦(T. aestivum, 2n=6x=42)与一粒小麦同源的为A染色体组,与圆锥小麦同源的是B染色体组;与圆锥小麦不同源的可表示为D染色体组,则染色体组型为AABBDD。
验证普通小麦的D染色体组来源1926Cybindricu (Ae. cylindrica染色体组。
六倍体小麦和柱穗小麦的杂种通常可发现有体(或不能配对。
1944草DD的形成。
1. 小麦属的传统属界:限定于栽培小麦及其近缘的野生种。
2. 小麦属与山羊草属分别是沿着栽培与野生两个方向进化的。
一年生草本。
山羊草属的生育期与小麦属的接近,有些略早于小麦,有些略小于小麦,通过分期(晚秋、初冬、早春三期)播种即可使两者花期相遇。
如果以普通小麦做母本,山羊草属做父本,较容易获得杂种种子;如果以山羊草属做母本,普通小麦做父本时,则较难获得,但如果利用“桥梁种”或幼胚培养等方法,均不难克服。
柱穗山羊草与粗山羊草的染色体组分析普通小麦的D染色体组(1)柱穗山羊草(染色体数为2n=2x=28)的D 染色体组与普通小麦的D染色体组同源,而C染色体组的存在说明:柱穗山羊草不是普通小麦D染色体组的直接供体;似乎一定有一个二倍体种DD,参与柱穗山羊草和普通小麦的形成。
人工合成六倍体小麦1. 1937年,有人发现利用秋水仙碱处理植物,可以较容易地使其染色体加倍,加倍率高达40%,加倍后一般不会引起突变,而且几乎对各种植物都有效。
2. 1944年,木原均等以二粒小麦与粗山羊草杂交后先得到具有“ABD”三组单倍染色体的杂种,再用秋水仙碱药剂处理杂种幼苗,使其染色体加倍,即可获得具有“AABBDD”染色体组型的人工合成六倍体小麦。
圆锥小麦×粗山羊草1. 调整花期:通过分期(晚秋、初冬、早春三期)播种即可使两者花期相遇。
2. 杂交技术:在小麦开花前,人工去雄、套袋,待柱头成熟时采集粗山羊草花粉用镊子或毛笔进行授粉。
在受精的次日,在重复授粉一次,可有效地提高杂交结实率。
3. 杂种F1的种子一般比较瘦瘪,发芽顶土能力差。
为了提高种子发芽和出苗率,可以将种子预先在0.1%升汞水溶液中浸5分钟,杀死附在种子表面的霉菌,并采用营养土苗床育苗。
用秋水仙碱诱导杂种F1染色体加倍的规范流程:1. 杂种种子(染色体组型为ABD)萌发,培育出壮苗。
2. 处理进行期间,温度要控制在15℃以下。
3. 将分蘖苗自苗圃挖出,洗净,然后用刀片在苗的基部割一浅伤口。
(处理后,染色体加倍的植物,割伤的比不割伤的可提高8.8% 。
)4. 分蘖苗基部浸在0.04-0.05%秋水仙碱水溶液中经过4天,光线要充足,而又不直晒。
如果秋水仙碱溶液中加入1.5-2%二甲基亚砜(DMSO)助渗剂,可使分生组织更活跃,促进秋水仙碱溶液渗入分生组织,以提高染色体加倍的效果。
5. 处理完毕后,分蘖苗要充分洗净,然后移植于温室中。
温室必须保持高湿(每天可在苗子上洒水两次)、低温(一般不超过10℃),直至分蘖苗从秋水仙碱的毒害中恢复过来。
(处理后的幼苗的根几乎全部死亡,必须小心地控制湿度和温度,才能使90%以上经过处理的分蘖苗恢复生长。
)6. 出现新的分蘖表明新根已经发生,分蘖苗已经恢复生长,此后的管理工作就应转到使恢复苗多发分蘖并使之尽可能成穗。
“人工合成六倍体”≠普通小麦 人工合成六倍体小麦原始品系存在籽粒不饱满和结实率低的缺点,可通过品系间相互杂交和系统选择的方法,使问题逐步得到解决。
1. 利用各种不同二粒小麦品种粗山羊草杂交,制造大量各种性状不同的人工合成六倍体小麦品系;2. 选择较好的人工合成六倍体小麦品系做亲本,进行品系间杂交和杂种后代的选育。
木原小麦(T. Kiharae)•木原小麦(T. Kiharae)也是一个人工合成六倍体,其染色体组型为AAGGDD。
(亲本为提莫菲维小麦AAGG×粗山羊草DD)•从理论上说,二倍体小麦与四、六倍体小麦,小麦与近缘植物间,还可以合成其它许多人工种。
(八倍体小黑麦AABBDDVV)•小麦进化过程中的第二次远缘杂交这一段历史,就由人工合成六倍体小麦实验而得到重复性验证。
验证普通染色体组B染色体组来源普通小麦是在进化过程中由几种野生麦类植物远缘杂交的不育种通过染色体加倍而形成的,自或其近似种羊草(1. A基因组从一粒小麦进化为二粒小麦,然后再进一步进化到现在普通小麦的分化率为0,而B基因组从二粒小麦进化到现在普通小麦就出现了0.61%的分化。
即A、B、D基因组在进化中DNA 序列的演化速度存在明显的差异,B基因组要快于A和D基因组,这可能是目前难以确定B基因组供体物种的症结所在。
2. 通过形态学、染色体配对、分子标记的研究,证明了拟斯卑尔托山羊草最有可能是B基因的供体,只是在进化过程中发生了部分改变。
染色体组分析法的局限性•染色体组分析是否能够成功进行,这在很大程度上决定于物种之间是否能够进行杂交、产生正常生长的杂种植物和该杂种是否能够进行减数分裂,缺乏这三个步骤的任何一环,染色体组分析均不能进行;•在减数分裂中期I所观察到的染色体密切配对,并不一定完全代表染色体上的每一基因位点核苷酸顺序的一致性;•染色体配对本身并不总是反映了物种间遗传上的相似性;即使染色体配对可以反映物种间染色体组的相似性,但染色体不配对并不就意味着物种间的非相似性;•由于染色体配对促进因子或抑制因子造成的染色体组间配对上的遗传变异将会歪曲不同的物种间染色体组配对的模式,进而造成对物种之间亲缘关系的错误估计。
•要做到尽可能客观、真实地解开小麦的起源和演化之谜,单靠染色体组分析法是不够的;•应该尽可能全面地利用不同的研究手段,综合分析尽可能多的研究结果和来源不同的信息(地理分布、细胞学资料以及分子水平的研究结果等)。
除普通小麦外,已有人通过实验证明胜利油菜(Brassica rapas)、陆地棉(Gossoypium hirstutum)和普通烟草(Nicotiana tabacum),也是通过不同种植物的远缘杂交形成的。
这些作物形成过程的基本原理与普通小麦进化过程一样。
参考文献•李振声. 《小麦远缘杂交》(1985).•卢宝荣, and 刘继红. "染色体组分析及小麦族的系统学." 植物学通报 9.1 (1992): 26-31.•余波澜, et al. "小麦 A, B 和 D 基因组同源DNA 序列在进化过程中的演变研究."遗传学报 28.7 (2001): 635-639.•崔娜欣, et al. "几种菊属植物及其种间杂种减数分裂行为观察." 园艺学报 33.5 (2006): 1033-1038.The End。