第三章 蛋白质的转运、加工与
- 格式:ppt
- 大小:25.18 MB
- 文档页数:159








细胞内各种蛋白质的合成和转运途径引言:细胞是生物体的基本单位,其中蛋白质是构成细胞的重要组成部分。
细胞内的蛋白质合成和转运途径是维持细胞正常功能的关键过程。
本文将介绍细胞内蛋白质合成的主要途径,包括转录、翻译和后转录修饰,以及蛋白质的转运途径,包括核糖体、内质网和高尔基体等。
一、蛋白质合成的途径1. 转录蛋白质合成的第一步是转录,即将DNA中的基因信息转录成RNA。
在细胞核中,DNA的双链解旋,RNA聚合酶结合到DNA上,根据DNA模板合成mRNA。
mRNA是一条单链RNA,它携带着从DNA中转录得到的基因信息。
2. 翻译翻译是蛋白质合成的第二步,即将mRNA上的基因信息翻译成蛋白质。
翻译发生在细胞质中的核糖体中。
核糖体由rRNA和蛋白质组成,它能够识别mRNA上的密码子,并将相应的氨基酸连接起来,形成多肽链。
翻译的过程包括起始、延伸和终止三个阶段,通过tRNA和蛋白因子的参与完成。
3. 后转录修饰蛋白质合成的最后一步是后转录修饰,即对新合成的蛋白质进行修饰和折叠。
这一过程发生在内质网和高尔基体中。
内质网是一个复杂的膜系统,它能够将新合成的蛋白质进行折叠和修饰,如糖基化、磷酸化等。
高尔基体则进一步对蛋白质进行修饰,并将其定位到细胞的不同位置。
二、蛋白质的转运途径1. 核糖体核糖体是蛋白质合成的场所,它位于细胞质中。
在核糖体中,mRNA上的密码子与tRNA上的反密码子互补配对,通过蛋白因子的辅助,将氨基酸连接成多肽链。
核糖体能够识别起始密码子和终止密码子,从而控制蛋白质的合成过程。
2. 内质网内质网是一个复杂的膜系统,它位于细胞质中。
内质网上的核糖体能够合成蛋白质,并将其进行折叠和修饰。
折叠不正确的蛋白质将被内质网上的分解酶降解,而正确折叠的蛋白质则会进一步转运到高尔基体或其他细胞器。
3. 高尔基体高尔基体是一个复杂的膜系统,它位于细胞质中。
高尔基体接收来自内质网的蛋白质,并对其进行进一步修饰和定位。

蛋白质的加工和运输1. 引言蛋白质是生物体内最重要的大分子之一,它们在生命过程中扮演着重要的角色。
蛋白质不仅是细胞结构的组成部分,还参与调节细胞的代谢过程、传递信号和执行功能等。
在细胞内,蛋白质的加工和运输至关重要,它们需要经过一系列的修饰和排序,以确保正确的定位和功能的实现。
本文将介绍蛋白质的加工和运输过程,包括蛋白质的合成、翻译后修饰和定位,以及蛋白质的运输和排序机制。
2. 蛋白质的合成蛋白质的合成是细胞中重要的生化过程之一。
在真核细胞中,蛋白质的合成发生在细胞质中的核糖体上。
蛋白质的合成包括两个主要步骤:转录和翻译。
转录是指DNA上的基因信息被转录成RNA分子的过程。
RNA分子是一条与DNA互补的单链分子,它携带了DNA上的遗传信息。
经过转录后,RNA分子称为mRNA(messenger RNA),它将带着基因信息离开细胞核,进入细胞质。
翻译是指mRNA上的遗传信息被翻译成氨基酸序列的过程。
翻译发生在细胞质中的核糖体上,它将mRNA上的三碱基密码子与特定的氨基酸配对,从而合成蛋白质的氨基酸序列。
3. 蛋白质的翻译后修饰和定位在翻译完成后,蛋白质通常需要经过一系列的修饰和定位才能实现其功能。
3.1 翻译后修饰翻译后修饰是指蛋白质在合成后进一步修饰的过程。
这些修饰包括磷酸化、甲基化、乙酰化等。
磷酸化是指蛋白质上的羟基被磷酸基团取代,这一修饰可以改变蛋白质的结构和功能。
甲基化和乙酰化则是通过将甲基和乙酰基添加到特定的氨基酸上,从而调节蛋白质的活性和稳定性。
3.2 蛋白质的定位蛋白质的定位是指将蛋白质定位到细胞的特定位置。
细胞内蛋白质的定位是由信号序列决定的,这些信号序列可以存在于蛋白质的氨基酸序列中。
这些信号序列被称为信号肽,它们可以将蛋白质定位到细胞质、细胞核、内质网、高尔基体或细胞膜等不同的位置。
定位蛋白质的机制涉及一系列的分子机制,包括信号识别粒子、转运蛋白、蛋白质通道等。
这些机制确保了蛋白质能够准确地定位到其执行功能的位置。

一、蛋白质的合成1、核糖体是合成蛋白质的机器,其功能是按照mRNA的指令由氨基酸合成蛋白质。
2、游离核糖体游离于胞质中,合成细胞内的基础蛋白质;附着核糖体,附着在内质网表面,构成粗面内质网的核糖体,合成分泌蛋白和膜蛋白。
3、蛋白质合成的一般过程:1)氨基酸的活化。
氨基酸和tRNA在氨酰—tRNA合成酶作用下合成活化的氨酰—tRNA。
2)起始、延伸和终止。
3)蛋白质合成后的加工。
肽链N端Met的去除;氨基酸残基的化学修饰,乙酰化、甲基化、磷酸化等;肽链的折叠;二硫键的形成。
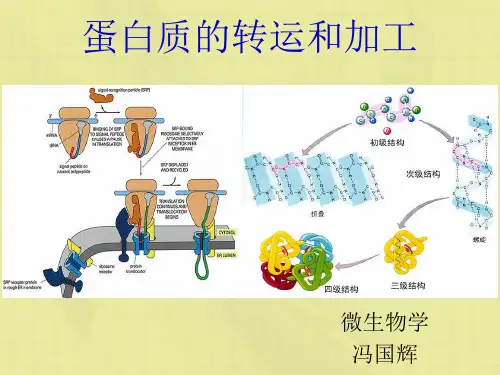
二、蛋白质的分泌合成、加工修饰和转运1、信号肽介导分泌性蛋白在粗面内质网的合成。
1)信号肽是蛋白质合成中最先被翻译出来的一段氨基酸序列,通常由18-30个疏水氨基酸组成,能指引核糖体与内质网结合,并引导合成的多肽链进入内质网腔。
2)新生分泌性蛋白质多肽链在胞质中的游离核糖体上起始合成。
当新生肽链N端的信号肽被翻译后,可立即被细胞质基质中的信号识别颗粒(SRP)识别、结合。
3)与信号肽识别结合的SRP,识别结合内质网膜上的SRP-R,并介导核糖体锚泊附着于内质网膜的通道蛋白移位子上。
而SRP则从信号肽—核糖体复合体上解离,返回细胞质基质中重复上述过程。
4)在信号肽的引导下,合成中的肽链,通过由核糖体大亚基的中央管和移位子蛋白共同形成的通道,穿膜进入内质网网腔。
随之,信号肽序列被内质网膜戗面的信号肽酶且除,新生肽链继续延伸,直至完成而终止。
最后完成肽链合成的核糖体大、小亚基解聚,并从内质网上解离。
2、跨膜驻留蛋白的插入和转移决定了蛋白质的两种去处:1)穿过膜进腔,为可溶性蛋白质,包括分泌蛋白和内质网驻留蛋白。
2)嵌入内质网膜中,形成膜蛋白。
3、粗面内质网与外输性蛋白质的分泌合成、加工修饰和转运过程密切相关。
1)新生多肽链的折叠与装配,与合成同时发生。
内质网为新生多肽链正确的折叠和装配提供了有利的环境。
分子伴侣通过对多肽链的识别结合来协助它们的折叠组装和转运。