第五节蛋白质合成后的加工及转运
- 格式:ppt
- 大小:2.11 MB
- 文档页数:86




蛋⽩质的合成、转运、修饰蛋⽩质的合成蛋⽩质的种类是由基因决定的,也就是说⼈类基因组有多少个基因,⼈体就有多少种蛋⽩质,只是蛋⽩质表达的时期和部位不同.根据⼈类基因组计划分析得知:全部⼈类基因组约有2.91Gbp,约有39000多个基因;也就是说⼈体蛋⽩质的种类有39000多种蛋⽩质⽣物合成可分为五个阶段,氨基酸的活化、多肽链合成的起始、肽链的延长、肽链的终⽌和释放、蛋⽩质合成后的加⼯修饰⼀.氨基酸的活化分散在胞液中的各种氨基酸需经特异的氨基酰-tRNA合成酶催化,ATP供能,并需Mg2+或Mn2+参与在氨基酸的羧基上进⾏活化,⽣成中间复合物()后者再与相应的tRNA作⽤,将氨基酰转移到tRNA分⼦的氨基酸臂上,即3′末端腺苷酸中核糖的3′(或2′)羟基以酯键相结合形成氨基酰-tRNA【氨基酰tRNA的⽣成】tRNA各种tRNA的⼀级结构互不相同,但它们的⼆级结构都呈三叶草形三叶草形结构的主要特征是:含有四个螺旋区、三个环和⼀个附加叉四个螺旋区构成四个臂,其中含有3′末端的螺旋区称为氨基酸臂,因为此臂的3′-末端都是C-C-A-OH序列,可与氨基酸连接三个环分别⽤Ⅰ、Ⅱ、Ⅲ表⽰环Ⅰ含有5,6⼆氢尿嘧啶,称为⼆氢尿嘧啶环(DHU环)环Ⅱ顶端含有由三个碱基组成的反密码⼦,称为反密码⼦环;反密码⼦可识别mRNA分⼦上的密码⼦,在蛋⽩质⽣物合成中起重要的翻译作⽤环Ⅲ含有胸苷(T)、假尿苷(ψ)、胞苷(C),称为假尿嘧啶环(TψC环);此环可能与结合核糖体有关tRNA在⼆级结构的基础上进⼀步折叠成为倒“L”字母形的三级结构起始因⼦原核起始因⼦只有三种(IF1、IF2、IF3)真核起始因⼦(简称为eIF)种类多且复杂,已鉴定的真核起始因⼦共有12种延长因⼦原核⽣物(简称EF)由三部分组成:EF-Tu,EF-Ts,和EF-GEF-Tu它介导氨酰-tRNA进⼊核糖体的空位EF-Ts充当EF-Tu亚基的鸟嘌呤核苷酸交换因⼦,催化EF-Tu释放GDPEF-G催化tRNA的移位和多肽延伸的每个循环后期mRNA从核糖体上掉下来真核⽣物(简称eEF)真核⽣物中分为:eEF-1和eEF-2eEF-1有两个亚基,α和βγα相当于原核⽣物中的EF-Tu亚基,它介导氨酰-tRNA进⼊核糖体的空位Βγ相当于原核⽣物中EF-Ts,核苷酸交换因⼦α,催化GDP从α上释放eEF-2相当于原核⽣物的EF-G,催化tRNA的移位和多肽延伸的每个循环后期mRNA从核糖体上掉下来终⽌因⼦(释放因⼦)原核⽣物细胞的释放因⼦(简称RF):识别终⽌密码⼦引起完整的肽链和核糖体从mRNA 上释放的蛋⽩质释放因⼦1(RF1):能识别终⽌密码⼦UAA和UAG⽽终⽌蛋⽩质合成的细菌释放因⼦释放因⼦2(RF2):能识别终⽌密码⼦UAA和UGA⽽终⽌蛋⽩质合成的细菌释放因⼦释放因⼦3(RF3):与延长因⼦EF-G有关的细菌蛋⽩质合成终⽌因⼦当它终⽌蛋⽩质合成时,它使得因⼦RF1和RF2从核糖体上释放真核⽣物细胞只有⼀种终⽌因⼦(称为eRF)能识别所有的终⽌密码⼦因为它没有与GTP结合的位点,所以它不能帮助完成合成的多肽从P位点的tRNA的释放在真核⽣物内可能还存在能与eRF合作、帮组多肽从核糖体释放的蛋⽩质核糖体的活性部位单个核糖体上存在四个活性部位,在蛋⽩质合成中各有专⼀的识别作⽤1.A部位:氨基酸部位或受位:主要在⼤亚基上,是接受氨酰基-tRNA的部位2.P部位:肽基部位或供位:主要在⼩亚基上,是释放tRNA的部位3.肽基转移酶部位(肽合成酶),简称T因⼦:位于⼤亚基上,催化氨基酸间形成肽键,使肽链延长4.GTP酶部位:即转位酶(EF-G),简称G因⼦,对GTP具有活性,催化肽键从供体部位→受体部位核糖体上还有许多与起始因⼦、延长因⼦、释放因⼦以及各种酶相结合的位点核糖体的⼤⼩是以沉降系数S来表⽰,S数值越⼤、颗粒越⼤、分⼦量越⼤原核细胞与真核细胞核糖体的⼤⼩亚基是不同的⼆.核糖体循环(肽链合成)1.肽链启动阶段在蛋⽩质⽣物合成的启动阶段,核蛋⽩体的⼤、⼩亚基,mRNA与⼀种具有启动作⽤的氨基酸tRNA共同构成启动复合体。
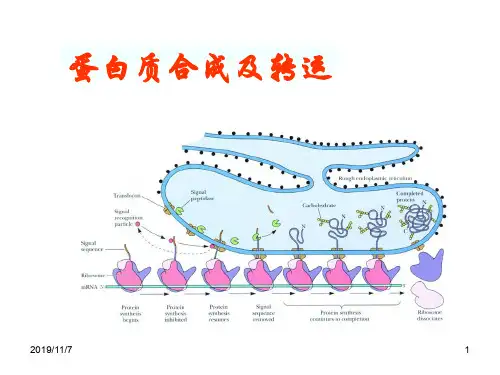




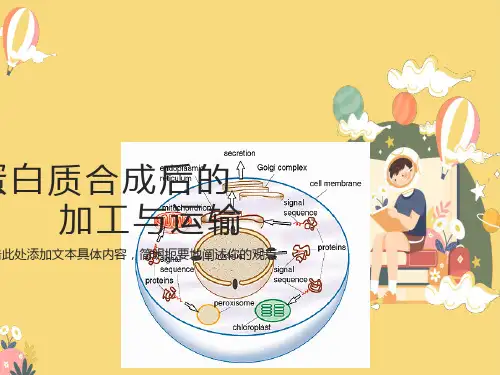

蛋白质的合成蛋白质的种类是由基因决定的,也就是说人类基因组有多少个基因,人体就有多少种蛋白质,只是蛋白质表达的时期和部位不同.根据人类基因组计划分析得知:全部人类基因组约有2.91Gbp,约有39000多个基因;也就是说人体蛋白质的种类有39000多种蛋白质生物合成可分为五个阶段,氨基酸的活化、多肽链合成的起始、肽链的延长、肽链的终止和释放、蛋白质合成后的加工修饰一.氨基酸的活化分散在胞液中的各种氨基酸需经特异的氨基酰-tRNA合成酶催化,ATP供能,并需Mg2+或Mn2+参与在氨基酸的羧基上进行活化,生成中间复合物()后者再与相应的tRNA作用,将氨基酰转移到tRNA分子的氨基酸臂上,即3′末端腺苷酸中核糖的3′(或2′)羟基以酯键相结合形成氨基酰-tRNA【氨基酰tRNA的生成】tRNA各种tRNA的一级结构互不相同,但它们的二级结构都呈三叶草形三叶草形结构的主要特征是:含有四个螺旋区、三个环和一个附加叉四个螺旋区构成四个臂,其中含有3′末端的螺旋区称为氨基酸臂,因为此臂的3′-末端都是C-C-A-OH序列,可与氨基酸连接三个环分别用Ⅰ、Ⅱ、Ⅲ表示环Ⅰ含有5,6二氢尿嘧啶,称为二氢尿嘧啶环(DHU环)环Ⅱ顶端含有由三个碱基组成的反密码子,称为反密码子环;反密码子可识别mRNA分子上的密码子,在蛋白质生物合成中起重要的翻译作用环Ⅲ含有胸苷(T)、假尿苷(ψ)、胞苷(C),称为假尿嘧啶环(TψC环);此环可能与结合核糖体有关tRNA在二级结构的基础上进一步折叠成为倒“L”字母形的三级结构起始因子原核起始因子只有三种(IF1、IF2、IF3)真核起始因子(简称为eIF)种类多且复杂,已鉴定的真核起始因子共有12种延长因子原核生物(简称EF)由三部分组成:EF-Tu,EF-Ts,和EF-GEF-Tu它介导氨酰-tRNA进入核糖体的空位EF-Ts充当EF-Tu亚基的鸟嘌呤核苷酸交换因子,催化EF-Tu释放GDPEF-G催化tRNA的移位和多肽延伸的每个循环后期mRNA从核糖体上掉下来真核生物(简称eEF)真核生物中分为:eEF-1和eEF-2eEF-1有两个亚基,α和βγα相当于原核生物中的EF-Tu亚基,它介导氨酰-tRNA进入核糖体的空位Βγ相当于原核生物中EF-Ts,核苷酸交换因子α,催化GDP从α上释放eEF-2相当于原核生物的EF-G,催化tRNA的移位和多肽延伸的每个循环后期mRNA从核糖体上掉下来终止因子(释放因子)原核生物细胞的释放因子(简称RF):识别终止密码子引起完整的肽链和核糖体从mRNA 上释放的蛋白质释放因子1(RF1):能识别终止密码子UAA和UAG而终止蛋白质合成的细菌释放因子释放因子2(RF2):能识别终止密码子UAA和UGA而终止蛋白质合成的细菌释放因子释放因子3(RF3):与延长因子EF-G有关的细菌蛋白质合成终止因子当它终止蛋白质合成时,它使得因子RF1和RF2从核糖体上释放真核生物细胞只有一种终止因子(称为eRF)能识别所有的终止密码子因为它没有与GTP结合的位点,所以它不能帮助完成合成的多肽从P位点的tRNA的释放在真核生物内可能还存在能与eRF合作、帮组多肽从核糖体释放的蛋白质核糖体的活性部位单个核糖体上存在四个活性部位,在蛋白质合成中各有专一的识别作用1.A部位:氨基酸部位或受位:主要在大亚基上,是接受氨酰基-tRNA的部位2.P部位:肽基部位或供位:主要在小亚基上,是释放tRNA的部位3.肽基转移酶部位(肽合成酶),简称T因子:位于大亚基上,催化氨基酸间形成肽键,使肽链延长4.GTP酶部位:即转位酶(EF-G),简称G因子,对GTP具有活性,催化肽键从供体部位→受体部位核糖体上还有许多与起始因子、延长因子、释放因子以及各种酶相结合的位点核糖体的大小是以沉降系数S来表示,S数值越大、颗粒越大、分子量越大原核细胞与真核细胞核糖体的大小亚基是不同的二.核糖体循环(肽链合成)1.肽链启动阶段在蛋白质生物合成的启动阶段,核蛋白体的大、小亚基,mRNA与一种具有启动作用的氨基酸tRNA共同构成启动复合体。