人类Y染色体DNA单倍型类群介绍2018
- 格式:docx
- 大小:1.62 MB
- 文档页数:12

目录 [隐藏]∙ 1 树形图∙ 2 单倍型类群A 和 B∙ 3 有M168 (CT)变异的单倍型类群 ∙ 4 单倍型类群 F (G, H & IJK) ∙ 5 单倍型类群 K (M9) ∙ 6 单倍型类群 NO (M214) ∙ 7 单倍型类群 P (M45) ∙ 8 单倍型类群在欧洲的时间发展 ∙ 9 同见 ∙ 10参考文献 ∙11 外部链接[编辑]树形图单倍型类群 S [编辑]单倍型类群A 和B单倍型类群A是非洲人的子单倍型类群,现代的所有单倍型类群起源点。
BT是单倍型类群A的分支。
它有两个主要谱系,单倍型类群B和CT。
定义突变分离CT(除A和B的所有单倍型类群)为M168和M294。
这些突变早于“走出非洲”的迁移。
DE突变的定义可能发生在非洲东北部,大约65,000年前。
[1] P143突变定义了单倍型类群CF。
可能发生在那时,将现代人类带至亚洲南部海岸。
亚, 密克罗尼西亚, 和玻利尼西亚▪单倍型类群C3分布▪单倍型类群D2 (M55, M57, M64。
1, M179, P12, P37。
1, P41。
1 (M359。
1), 12f2。
2)▪单倍型类群D3 (P47)▪单倍型类群E(Y-DNA)分布单倍群F和其后代分布▪单倍型类群F*分布于南部印度, 斯里兰卡, 云南, 朝鲜半岛▪单倍型类群G分布▪单倍型类群G2c1▪单倍型类群H分布45000年前分离▪单倍型类群I分布岛 I2B1 (m223)主要分布于西部, 中部,和北欧。
▪单倍型类群J分布▪单倍型类群L (M20) 分布于南亚, 中亚, 西南亚,地中海▪单倍型类群T分布单倍型类群O分布单倍型类群Q分布Q被定义由SNP M242。
认为出现在大约35000-40000年前的中亚。
单倍型类群Q的亚类型和定义变异,根据2008 ISOGG树[4]在下文提供。
ss4 bp, rs41352448, 不出现在ISOGG 2008树因为STR。

人类23条染色体的故事——Y染色体父系遗传五百年前是一家Y染色体基因从Y染色体的角度来讲,Y是决定男性体征的关键染色体,而且绝对遗传自父亲。
经过N代的繁衍,Y染色体基本相同,但是也逐代递减,也就是说,确实做到了正宗遗传,我们常说“五百年前是一家”。
不管我们如何以Y染色体来界定遗传的正宗与否,有这样一个事实我们不得不承认,Y染色体是个非常不稳定的染色体,都快要因为自己本身的缺陷丢失掉自己的所有遗传密码了,光从染色体的本性来说,Y绝对是个弱者。
Y染色体真小Y染色体是人体染色体中最短的,长度只有X染色体的1/3,而且正逐渐退化,大多数序列无实质功能,包含的遗传信息非常少,Y染色体由诞生之初的1438个基因,衰减到现在仅剩的45个基因。
事实上,Y染色体上没有生命必需的基因,因而人类的女性虽然不拥有Y染色体也可以正常存活,Y染色体的少数功能基因基本上只与男性特征有关(惟一明确的与Y染色体的缺陷有关的人类疾病是睾丸发育不良)。
从功能的角度来讲,所有正常男性的Y染色体都是一样的。
母系遗传就是线粒体基因,人身体所有细胞里面都有线粒体,mtDNA(线粒体DNA)是承载线粒体遗传密码的物质,其表现为母系遗传,也就是说,只有女性的线粒体基因能随其卵子遗传给后代,线粒体基因组是16569个碱基对组成的双股DNA分子,有37个基因。
人类23对染色体受精过程不是两个细胞的单纯融合,是精子的细胞核进入卵子,父母双方的核内基因混合,再加上母亲的这37个基因,才是下一代的全部基因,这37个基因只通过母系一脉来传承,男性也能从母亲那里继承,但是无法将它遗传给后代。
也就是说,如果一个女人生的全是儿子,她的这37个基因就传丢了。
从线粒体基因的角度来讲,人类应该是母系遗传?就象Y染色体可以追溯到父亲、祖父、曾祖父乃至以上无数代的父系宗谱一样,线粒体基因组也可以追溯到妈妈、外婆、外婆她妈乃至以上无数代的母系宗谱,与Y染色体不断消亡不同的是线粒体基因一般很难发生改变,平均每2万年才会发生微小的变异,因此,mtDNA结构类型是反映母系脉络的重要指标,通过检测现代人mtDNA,能弄清各民族、各地人的母系血缘关系。

Y染色体,简单却又复杂的不定游者摘要:Y染色体是属于XY性别决定系统的大多数哺乳动物的两条性染色体之一,其中SRY 基因控制着睾丸的形成而决定是否为雄性。
Y染色体的变异会引发罕见或常见的遗传病,通过对其研究,不仅可以找出致病原因,还可研究Y染色体自身功能,亦能追溯家族或人群起源。
它由常染色体演化而来,一亿多年以来不断改变,直到新的性别决定系统产生而完成自己的遗传变异使命。
关键词:Y染色体;遗传病;SRY;缺陷;微缺失Y染色体的存在:在XY性别决定系统中,雄性个体所特有的性染色体是Y染色体,在哺乳动物的Y染色体中含有的SRY基因能触发睾丸的生长,并由此决定雄性性状。
具有Y染色体的非哺乳动物包括部分昆虫(如果蝇)、鱼类、爬行动物,部分植物也具有Y染色体。
就黑腹果蝇而言,Y染色体并不引起雄性果蝇的发育,包含对雄性生育能力必要的基因。
性别决定于X染色体的数目,基因型为XXY的黑腹果蝇为雌性,基因型为XO,只拥有单个X染色体的为雄性但不能生育。
那么Y染色体的起源是怎样的呢?最近鸭嘴兽基因组测序的研究表明,XY性别决定系统只是在大约1.66亿年以前出现的,是在原兽亚纲从其他哺乳动物中分离出来开始的。
(Warren WC et al 2008)X染色体和Y染色体被认为是由一对相同的染色体演变而来的,这对染色体原属于常染色体。
某个远古哺乳动物的祖先发生了等位基因的变异,只要拥有这对等位基因的个体就会成为雄性。
包含这对等位基因之一的染色体最终形成了Y染色体,另一半的染色体最终形成了X染色体。
Y染色体不断通过染色体易位获得对雄性个体有利而雌性个体有害或没有明显作用的基因。
(Grützner .F et al 2004)Y染色体的结构与功能:人类的Y染色体中包含约6千万个碱基对,约占男性体细胞中DNA的2%。
有86个基因,这些基因只编码了23种不同的蛋白质。
人类的Y染色体除了在端粒上的拟常染色体区的少部分片段,只占有染色体长度约5%,能与相应的X染色体发生重组,其外都不能发生重组。
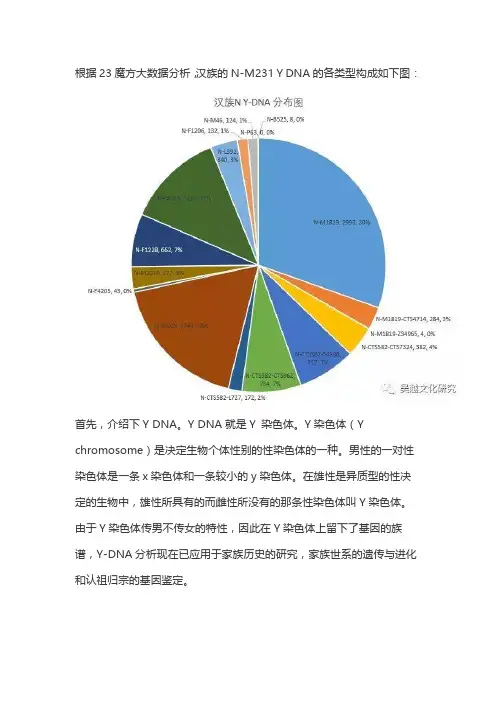
根据23魔方大数据分析,汉族的N-M231 Y DNA的各类型构成如下图:首先,介绍下Y DNA。
Y DNA就是Y 染色体。
Y染色体(Y chromosome)是决定生物个体性别的性染色体的一种。
男性的一对性染色体是一条x染色体和一条较小的y染色体。
在雄性是异质型的性决定的生物中,雄性所具有的而雌性所没有的那条性染色体叫Y染色体。
由于Y染色体传男不传女的特性,因此在Y染色体上留下了基因的族谱,Y-DNA分析现在已应用于家族历史的研究,家族世系的遗传与进化和认祖归宗的基因鉴定。
N-M231 是现代人类父系单倍群类型N-M231主要分布于欧亚大陆北方。
Y-N是Y-NO子孙,被认为是一个相对年轻的单倍群。
Y-N和Y-O分离后,没有翻过青藏高原,沿北线,于21000年前在阿尔泰山一带分化出N1和N2(N2只分布于阿尔泰山、东欧、中欧、巴尔干半岛,微基因样本中中国地区有一例金姓N2,23魔方用户中有河南郑氏家族属于N2 N-B482下游),N2向西部挺进,N1向东部挺进,从中国西部进入中国。
16200年前N1在祁连山或河湟一带分化为N1a和N1b。
一部分N1b向西藏、青海、云南、四川、贵州以及中南半岛及印度挺进。
另一部分N1b跟随N1a继续东进。
15000年前N1a在河套到燕山一带分化成N1a1和N1a2。
6000至8000年前N1a1从燕山一带向西北迁徙,N1a1 的一支3500年前到达波罗的海。
波罗的海N-YDNA最早的古代遗骨发现在3500年左右。
N-P43高频于北极涅涅茨人群。
N-M128高频于中国北方地区。
N-M46大约有1.4万岁,是Y-N系统中最高频的支系。
他可能起源于今天的中国,随后在西伯利亚经历了严重的瓶颈,高频于雅库特人中,并向东欧方向进行第二次扩张。
N1a1主要发现于东北欧人群,尤其是芬兰(63%), 拉普兰(斯堪的纳维亚半岛的最北端地区,53%), Sami人(47%)爱沙尼亚(40%), 拉脱维亚(43%), 立陶宛(42%)和北俄罗斯(39%), 也中低频分布于中俄罗斯(16%), 白俄罗斯(10%), 乌克兰(6%), 瑞典(14%), 挪威(4%)波兰(3%) 和土耳其(4%)捷克(1.6%),德国(1.6%)。
目录 [隐藏]∙ 1 树形图∙ 2 单倍型类群A 和 B∙ 3 有M168 (CT)变异的单倍型类群 ∙ 4 单倍型类群 F (G, H & IJK) ∙ 5 单倍型类群 K (M9) ∙ 6 单倍型类群 NO (M214) ∙ 7 单倍型类群 P (M45) ∙ 8 单倍型类群在欧洲的时间发展 ∙ 9 同见 ∙ 10参考文献 ∙11 外部链接[编辑]树形图单倍型类群 S [编辑]单倍型类群A 和B单倍型类群A是非洲人的子单倍型类群,现代的所有单倍型类群起源点。
BT是单倍型类群A的分支。
它有两个主要谱系,单倍型类群B和CT。
定义突变分离CT(除A和B的所有单倍型类群)为M168和M294。
这些突变早于“走出非洲”的迁移。
DE突变的定义可能发生在非洲东北部,大约65,000年前。
[1] P143突变定义了单倍型类群CF。
可能发生在那时,将现代人类带至亚洲南部海岸。
亚, 密克罗尼西亚, 和玻利尼西亚▪单倍型类群C3分布▪单倍型类群D2 (M55, M57, M64。
1, M179, P12, P37。
1, P41。
1 (M359。
1), 12f2。
2)▪单倍型类群D3 (P47)▪单倍型类群E(Y-DNA)分布单倍群F和其后代分布▪单倍型类群F*分布于南部印度, 斯里兰卡, 云南, 朝鲜半岛▪单倍型类群G分布▪单倍型类群G2c1▪单倍型类群H分布45000年前分离▪单倍型类群I分布岛 I2B1 (m223)主要分布于西部, 中部,和北欧。
▪单倍型类群J分布▪单倍型类群L (M20) 分布于南亚, 中亚, 西南亚,地中海▪单倍型类群T分布单倍型类群O分布单倍型类群Q分布Q被定义由SNP M242。
认为出现在大约35000-40000年前的中亚。
单倍型类群Q的亚类型和定义变异,根据2008 ISOGG树[4]在下文提供。
ss4 bp, rs41352448, 不出现在ISOGG 2008树因为STR。

人类Y染色体410、Y-N单倍群现代人类父系单倍群类型N-M231主要分布于欧亚大陆北方。
Y-N是Y-NO 子孙,被认为是一个相对年轻的单倍群。
N的一些支系也低频发现于东南亚、太平洋群岛、西南亚和巴尔干半岛。
这些因素导致认为N可能起源于东亚或东南亚。
N系人群大约2.1万年前出现于东南亚,并从东南亚向北扩散,大约1.4-1.2万年前从东亚北部扩散进入西伯利亚,大约1万到8千年前扩散进入中亚和欧洲。
然而N系人群没有能进入美洲,其历史原因仍然待解。
N系虽然被认为起源于东南亚,但其下游人群得到发展,高频出现于北欧的芬兰(60%)、波罗的海(45%)和斯堪的纳维亚的萨米(40%)人群中,东普鲁士的德国人约有28%的N。
N在西伯利亚的突厥语系雅库特人(Yakuts)和北极地区的涅涅茨人(Nenets)中占有75%。
N1*-CTS11499原来也叫N1*-LLY22g,N1b-P43高频于北极涅涅茨人群。
N1a-M128高频于乌拉尔地区。
N1c1-M46大约有1.4万岁,是Y-N系统中最高频的支系。
他可能起源于今天的中国,随后在西伯利亚经历了严重的瓶颈,高频于雅库特人中,并向东欧方向进行第二次扩张。
N1c1主要发现于东北欧人群,尤其是芬兰 (61%), 拉普兰(斯堪的纳维亚半岛的最北端地区,53%), 爱沙尼亚(34%), Latvia (38%), 立陶宛(42%)和北俄罗斯(30%), 也中低频分布于中俄罗斯(15%), 白俄罗斯(10%), 东乌克兰(9%), 瑞典(7%), 波兰(4%) 和土耳其(4%)。
N1c 也突出分布于说乌拉尔语种族的伏尔加-乌拉尔地区,包括Udmurts (67%)、Komi (51%)、 Mari (50%) 和Mordvins (20%),也存在于突厥语邻居中,如楚瓦什人(28%)、伏尔加鞑靼人(21%) 和巴什基尔人(17%),以及俄罗斯南部Nogais(9%) 。
在红山文化牛河梁遗址6例古DNA中有4例属于N系,红山-小河沿文化的哈拉海沟遗址中12例全部为N。

人类染色体详细资料大全染色体(chromosome)是遗传物质,基因的载体,人类的常染色体是成对存在的。
人体的体细胞染色体数目为23对,其中22对为男女所共有,称为常染色体(autosome);另外一对为决定性别的染色体,男女不同,称为性染色体(sex chromosome),男性为XY,女性为XX。
在生殖细胞(generative cell)中,男性生殖细胞染色体的组成:22条常染色体+X或Y。
女性生殖细胞染色体的组成:22条常染色体+X。
基本介绍•中文名:人类染色体•外文名:Human chromosome•定义:人体的遗传物质,基因的载体形态特征,常见疾病,染色质,类别分析,临床套用,常见病例,先天愚型,18三体,13三体,5p-,性染色体,两性畸形,畸变原因,具体数目,结构,形态特征根据着丝粒的位置不同,把人类染色体分为三种类型:①中央着丝粒染色体(metacentric hrmosome),着丝粒位于染色体纵轴的1/2~5/8处;②亚中着丝粒染色体(submetacentric chromosome),着丝粒位于染色体纵轴的5/8~7/8处;③近端着丝粒染色体(acrocentric chromosome),着丝粒位于染色体纵轴的7/8至末端。
染色体分组、核型与显带技术(一)Denver体制为了更好、更准确地表达人体细胞的染色体组成,1960年,在美国丹佛(Denver)市召开了第一届国际细胞遗传学会议,讨论并确立了世界通用的细胞内染色体组成的描述体系―Denver体制。
这个体制按照各对染色体的大小和着丝粒位置的不同将22对染色体由大到小依次编为1至22号,并分为A、B、C、D、E、F、G共7个组,X和Y染色体分别归入C组和G组。
一个体细胞中的全部染色体所构成的图像即称核型。
将待测细胞的全部染色体,按照 Denver体制配对、排列后,分析确定其是否与正常核型完全一致,就叫核型分析(karyotype ***ysis)。

因纽特人y染色体单倍群dna的类型标题:探寻因纽特人y染色体单倍群DNA的类型及其文化意义导言1. y染色体单倍群DNA的概念和重要性在人类遗传学研究中,y染色体单倍群DNA是指由父系传承的DNA序列,通过对不同群体的y染色体单倍群DNA进行分析,可以揭示人类个体之间的遗传联系,同时也能够为人类种群的起源、迁徙和遗传多样性提供重要线索。
因纽特人作为北极地区的土著民族,其y染色体单倍群DNA类型的研究对于揭示他们的遗传渊源、文化演变及人类学研究有着重要意义。
2. 因纽特人的历史和文化背景作为北极圈内的原住民族,因纽特人(Inuit)主要居住在加拿大、格陵兰和阿拉斯加等地,他们有着悠久的狩猎和捕鱼传统,过着以捕鱼、游牧和海洋产物为生的生活。
在长期的气候环境适应过程中,因纽特人形成了独特的文化和社会制度,传承着丰富的口头传统和宗教信仰,其社会结构和生活方式也受到地理环境和资源条件的影响。
3. 研究目的和意义本文将从因纽特人y染色体单倍群DNA的类型出发,探讨他们的遗传源流、族群关系及文化发展,通过对y染色体单倍群DNA的研究,寻找因纽特人文化与遗传的联系,进一步了解他们的起源和历史。
正文一、因纽特人y染色体单倍群DNA的类型介绍根据相关研究,因纽特人的y染色体单倍群DNA类型主要包括A2a、C2a、Q1a等,在这些类型中,A2a是最为常见的,这与他们亲缘关系的研究结果相一致。
对不同因纽特人群体的研究发现,这些y染色体单倍群DNA类型的分布存在一定的地域差异,在不同地区形成了特定的遗传分布格局。
二、因纽特人y染色体单倍群DNA与世界各地其他人群的比较通过与其他世界各地人群的y染色体单倍群DNA比较分析,可以发现一些有趣的现象。
比如与东亚、西伯利亚和北美印第安人的关系密切,同时又与北欧以及格陵兰因纽特人有着一定的联系等。
这些比较分析可以为因纽特人的迁徙史和遗传亲缘关系提供重要线索。
三、因纽特人y染色体单倍群DNA的文化意义在因纽特人的文化传统中,包括狩猎、图腾崇拜、宗教信仰等诸多方面都与他们的遗传联系密切。

人类Y染色体36、Y-C单倍群Y染色体C-M130单倍群是Y-CF两个分支中的一支,C的地位与F相当。
Y-C 人群发现于除非洲以外的各个大陆古代人群中,是中亚、西伯利亚、北美和大洋洲一些土著部落的主流单倍群。
在早期时候,这个单倍群分支比较多,但在二叉树现象规律整合下,这个单倍群目前也归类为两大分支,分别是C1-F3393/Z15426(包括之前的C1,C2, C4,C5,C6)和C2(之前为C3)。
C-M130单倍群似乎在SNP突变M168产生之后不久就已出现,年龄大约有6-5.3万年。
虽然C-M130高频发现于蒙古、俄罗斯远东地区、波利尼西亚、澳大利亚一些土著人群中,以及中频发现于朝鲜和满族,但是C-M130最大多样性发现于印度现代人口中。
现在假设C-M130起源于印度或南亚大陆海岸,并经历过长时间的演化,再扩散于东南亚,大约4万年前C向北进入东亚。
大约8000-6000年前,推测C-M130被说纳-得内语的人群带进美洲,主要分布于北美西北部太平洋岸边。
Y-C的支系也扩散于欧洲。
C-F1171系统各分支分布于南欧、中东、印度、东南亚、日本、太平洋群岛和澳大利亚等地。
与Y-F下的"GHIJKLMST"各支系在这些地区扩张具有分布相似性。
C-M217系统(即原来C3)则分布分布于欧亚大陆北方,与K系下NOQR分布也具有相似性。
推理C系统人群和F系统人群在出非洲后,就有相当程度伴随性。
至于原C3南支和北支与N和O的分布也具有相似性。
C-M217高频发现于中亚、西伯利亚和北美土著人群。
在布里亚特、鄂温克、蒙古、哈萨克等民族中都有高频的M217成分。
C-M217(原C3)在汉族中分布从0-23%分布不等,在广西、湖南、江西地区汉族中基本缺失,在西安样本测试中出现最高23%的C3成分。
7、Y-D单倍群Y-D单倍群是是DE单倍群下的另一分支。
DE单倍群是由SNP M1(YAP)定义,它的独特性在于全球地理分布上有几个清晰的分离分支。

现代人的Y染色体谱系是根据人类Y染色体DNA单倍型类群(haplogroups)来划分的。
Y染色体DNA单倍型类群是通过遗传标记(SNPs)的差异来确定的,这些标记代表了基因突变,并且能够反映不同个体之间的进化关系。
截至我知识更新日期,人类Y染色体谱系树分为18个主要类型,用从A到R的十八个字母作为索引,每个主类型下又有多个次类型和子类型,由数字和小写字母表示。
这些类型的命名反映了它们在演化树中的位置以及与特定突变的关系。
例如,类型A是最古老的Y染色体类型,被认为是从非洲起源的人类共同祖先所拥有的类型。
随着人类迁徙和发展,其他类型如B、C、D等逐渐形成,每一个类型都有其独特的地理分布和历史背景。
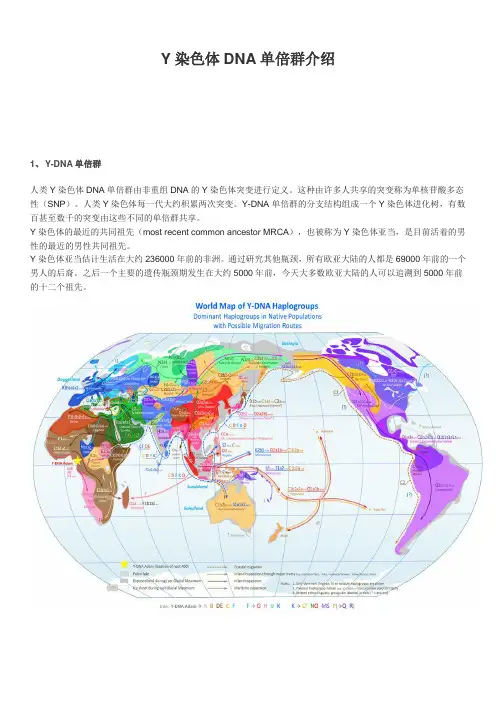
Y染色体DNA单倍群介绍1、Y-DNA单倍群人类Y染色体DNA单倍群由非重组DNA的Y染色体突变进行定义。
这种由许多人共享的突变称为单核苷酸多态性(SNP)。
人类Y染色体每一代大约积累两次突变。
Y-DNA单倍群的分支结构组成一个Y染色体进化树,有数百甚至数千的突变由这些不同的单倍群共享。
Y染色体的最近的共同祖先(most recent common ancestor MRCA),也被称为Y染色体亚当,是目前活着的男性的最近的男性共同祖先。
Y染色体亚当估计生活在大约236000年前的非洲。
通过研究其他瓶颈,所有欧亚大陆的人都是69000年前的一个男人的后裔。
之后一个主要的遗传瓶颈期发生在大约5000年前,今天大多数欧亚大陆的人可以追溯到5000年前的十二个祖先。
Y-DNA单倍群进化树单倍群 A & B 单倍群 A(M91)单倍群A是所有单倍群起源点。
现代所有单倍群都是单倍群A的后代,稀疏分布在非洲,主要集中在西南部的科伊桑人和尼罗河谷东北部人群。
单倍群 BT (M42,M94,M139,M299)约55000年前分,BT是单倍群A的分支单倍群B(M60)单倍群B主要分布于非洲,主要集中于俾格米人群。
详细树形图:见B单倍群文件夹单倍群 CT (P143)标识单倍群 CT的突变标记是M168和M294.包含单倍群D、E、C、F,可能88000年前在亚洲或非洲出现。
单倍群 C (M130)历史起源:C单倍群携带M130突变,来源于CF单倍群。
中国境内的C单倍群主要是C2(携带M217突变),占中国总人口比例大约为5%—10%。
其下游又可分为南北两大支,北支C2b(携带F1396突变),主要分布于蒙古族和满族等民族;南支C2c(携带F1067突变),几乎遍及全中国。
详细树形图:见C单倍群文件夹Haplogroup C (M130, M216) 分布在亚洲、大洋洲和北美等o Haplogroup C1 (F3393/Z1426)▪Haplogroup C1a (CTS11043)▪Haplogroup C1a1 (M8, M105, M131) 日本低频分布▪Haplogroup C1a2 (V20) 欧洲和尼泊尔低频分布▪Haplogroup C1b (F1370, Z16480)▪Haplogroup C1b1 (AM00694/K281)▪Haplogroup C1b1a (B66/Z16458)▪Haplogroup C1b1a1 (M356) 印度低频分布, 阿拉伯半岛和中国北部▪Haplogroup C1b2 (B477/Z31885)▪Haplogroup C1b2a (M38) 分布在印度尼西亚,新几内亚岛,美拉尼西亚,密克罗尼西亚,和玻利尼西亚▪Haplogroup C1b2b (M347, P309) 澳洲土著o Haplogroup C2 (M217, P44) 分布在欧亚大陆和北美,特别是在蒙古人,哈萨克人,通古斯人,西伯利亚人,和Na-Dené-speaking语民族单倍群DE(M1,M145,M203)约65000年前分离单倍群D (M174)详细树形图:见D单倍群文件夹∙Haplogroup D (M174) 分布在日本、中国(特别分布于西藏)和安达曼岛o D1 (CTS11577)▪D1a (Z27276, Z27283, Z29263)▪Haplogroup D1a1 (M15) 主要分布在西藏、羌族、彝族和苗瑶语人群▪Haplogroup D1a2 (P99) 主要分布在西藏、羌族、纳西族、突厥部落▪Haplogroup D1b (M55, M57, M64.1, M179, P12, P37.1, P41.1 (M359.1), 12f2.2) 主要在日本o D2 (L1366, L1378, M226.2) 菲律宾、麦克坦岛Haplogroup E (M96)详细树形图:见E倍群文件夹∙Haplogroup E (M40, M96) 分布在非洲、中东和欧洲o Haplogroup E1 (P147)▪Haplogroup E1a (M33, M132) 旧称E1▪Haplogroup E1b (P177)▪Haplogroup E1b1 (P2, DYS391p); 旧称E3▪Haplogroup E1b1a (V38) 非洲尼日尔-刚果语人群; 旧称E3a▪Haplogroup E1b1b (M215) 非洲之角,北非、中东和欧洲地中海地区; 旧称E3bo Haplogroup E2 (M75)Haplogroup F (M89)单倍群F和后代迁徙图单倍群F和后代构成了目前世界人口的90%,几乎都分布在撒哈拉以南非洲地区之外。
中国人种基因图谱随着分子人类学数据的不断积累,父系Y染色体与母系mtDNA研究的一系列进展,使得中国人群的多样性结构逐渐明晰。
现有的Y染色体数据揭示,现代人出非洲后由东南亚经多次迁徙进入东亚。
在旧石器时代,现代人最初定居东亚或东南亚之后,紧接着不断北迁,这奠定了中国人遗传结构的基础。
通过了解人类基因的遗传成分,绘制中国人种基因图谱。
中国人种基因图谱:一对为性染色体,XY组合的为男性,XX组合的为女性。
Y染色体只在父亲与儿子传代,呈严格的父系遗传,研究Y染色体,可以发现人群在父系关系上的迁徙和发展。
母系mtDNA表现为母系遗传。
通过检测现代人mtDNA,能弄清各民族、各地人的母系血缘关系。
通过研究Y-DNA与mtDNA的重合型,可以揭开中国人的祖先来源之谜。
单倍群人类父系基因Y-DNA人类母系基因mtDNAY-DNA与mtDNA的重合型中国人起源华夏族的起源与形成单倍群在分子进化的研究中,单倍群或单倍型类群是一组类似的单倍型,它们有一个共同的单核苷酸多态性祖先。
因为单倍群由相似的单倍型组成, 所以可以从单倍型来预测单倍群.单核苷酸多态性试验被用来确认单倍型。
单倍群以字母来标记,并且以数字和一些字母来做补充,,例如O3a4。
Y染色体和线粒体单倍群有不同的单倍群标记方法。
单倍群用来标记数千年前的祖先来源。
在人类遗传学中, 最普遍被研究的单倍群是『人类Y染色体脱氧核糖核酸单倍群(Y-DNA单倍群)』和『人类线粒体脱氧核糖核酸单倍群(mtDNA单倍群)』,这两个都可以被用来定义遗传群体。
Y染色体脱氧核糖核酸单倍群仅仅被从父系线遗传,同时mtDNA仅仅被从母系线遗传。
人类父系基因Y-DNA在人类基因学里,人类Y染色体DNA单倍型类群通过Y染色体遗传变异特性进行人类学研究的一门科学,主要用于研究人类的“非洲起源论”及以后的种群分布的遗传学证据。
人类有23对46条染色体,其中22对44条为常染色体,另外一对为性染色体,XY组合的为男性,XX组合的为女性。
• • • • • • • • • • •单倍型类群S [编辑]单倍型类群A 和B单倍型类群A是非洲人的子单倍型类群,现代的所有单倍型类群起源点。
BT是单倍型类群A的分支。
它有两个主要谱系,单倍型类群B和CT。
定义突变分离CT(除A和B的所有单倍型类群)为M168和M294。
这些突变早于“走出非洲”的迁移。
DE突变的定义可能发生在非洲东北部,大约65,000年前。
[1] P143突变定义了单倍型类群CF。
可能发生在那时,将现代人类带至亚洲南部海岸。
▪单倍型类群C3分布▪单倍型类群E(Y-DNA)分布单倍群F和其后代分布▪单倍型类群G分布▪单倍型类群H分布▪单倍型类群IJ (P123, P124, P125, P126, P127, P129, P130) 约45000年前分离▪单倍型类群I分布(m223)主要分布于西部, 中部,和北欧。
▪单倍型类群J分布▪LT (见下)▪单倍型类群T分布▪单倍型类群S (M230) (之前被称为单倍型类群K5) 分布于新几亚高地单倍型类群O分布单倍型类群Q分布Q被定义由SNP M242。
认为出现在大约35000-40000年前的中亚。
单倍型类群Q的亚类型和定义变异,根据2008 ISOGG树[4]在下文提供。
ss4 bp,rs41352448, 不出现在ISOGG 2008树因为STR。
作为一种新型的Q系(Q5)在印度人中已经被低频率发现[5]2008 ISOGG树▪Q (M242)▪Q*▪Q1 (P36。
2)▪Q1*▪Q1a (MEH2)▪Q1a3a*▪Q1a3a1 (M19) 分布于一些南美原住民, 如en:Ticuna和瓦尤人[8]▪Q1a3a2 (M194)▪Q1a3a3 (M199, P106, P292)▪Q1a4 (P48)▪Q1a5 (P89)▪Q1a6 (M323) 极少分布于也门犹太人单倍群R和其后代分布单倍群R和其后代分布。
• • • • • • • • • ••单倍型类群 S [编辑]单倍型类群A 和B单倍型类群A是非洲人的子单倍型类群,现代的所有单倍型类群起源点。
BT是单倍型类群A的分支。
它有两个主要谱系,单倍型类群B和CT。
定义突变分离CT(除A和B的所有单倍型类群)为M168和M294。
这些突变早于“走出非洲”的迁移。
DE突变的定义可能发生在非洲东北部,大约65,000年前。
[1] P143突变定义了单倍型类群CF。
可能发生在那时,将现代人类带至亚洲南部海岸。
▪单倍型类群C3分布▪单倍型类群E(Y-DNA)分布单倍群F和其后代分布▪单倍型类群G分布▪单倍型类群H分布▪单倍型类群IJ (P123, P124, P125, P126, P127, P129, P130) 约45000年前分离▪单倍型类群I分布岛 I2B1 (m223)主要分布于西部, 中部,和北欧。
▪单倍型类群J分布▪LT (见下)▪单倍型类群T分布▪单倍型类群S (M230) (之前被称为单倍型类群K5) 分布于新几内亚高地单倍型类群O分布单倍型类群Q分布Q被定义由SNP M242。
认为出现在大约35000-40000年前的中亚。
单倍型类群Q的亚类型和定义变异,根据2008 ISOGG树[4]在下文提供。
ss4 bp, rs41352448, 不出现在ISOGG 2008树因为STR。
作为一种新型的Q系(Q5)在印度人中已经被低频率发现[5]2008 ISOGG树▪Q (M242)▪Q*▪Q1 (P36。
2)▪Q1*▪Q1a (MEH2)▪Q1a3a*▪Q1a3a1 (M19) 分布于一些南美原住民, 如en:Ticuna和瓦尤人[8]▪Q1a3a2 (M194)▪Q1a3a3 (M199, P106, P292)▪Q1a4 (P48)▪Q1a5 (P89)▪Q1a6 (M323) 极少分布于也门犹太人单倍群R和其后代分布单倍群R和其后代分布。
染色体单倍群染色体是一种存在于细胞核中的结构体,其中包含了生物个体遗传信息的大部分。
染色体单倍群(chromosome haplogroup)指的是一组具有相似染色体序列的个体,这些个体在染色体上的变异在人类进化过程中被保留下来,并且在人类群体中具有一定的遗传意义。
本文将详细介绍染色体单倍群的相关知识,并探讨其在人类研究和基因组学中的应用。
染色体单倍群的概念最早由人类基因组学家Cann et al(1987)提出。
他们使用线粒体DNA研究了人类起源和迁徙的历史,发现了几个具有共同祖先的线粒体单倍群。
线粒体DNA由母亲传给子女,因此线粒体单倍群能够追溯到远古时代的女性。
随后,人类基因组项目的展开使得染色体单倍群的研究逐渐扩展到整个染色体组。
现在,我们可以基于单倍群的遗传变异和时间序列来探索人类起源和迁徙的历史。
染色体单倍群与遗传多样性密切相关。
不同的染色体单倍群在人类群体中的分布不均,反映了历史上人类迁徙的路径与模式。
比如,非洲是人类起源的地方,因此非洲人群中的染色体单倍群多样性最高。
而欧洲、亚洲、澳大利亚和美洲等地的人类群体则呈现出不同的单倍群分布模式,揭示了人类迁徙和历史事件的重要线索。
通过对染色体单倍群的研究,我们可以了解到不同人群之间的遗传差异。
例如,非洲人群的染色体单倍群具有更高的多样性,而亚洲和欧洲人群则呈现出明显的集中性,这可能反映了人类历史上的群体迁徙和天然选择的结果。
染色体单倍群研究还可以研究和追溯一些古代人类群体和祖先的遗传特征,比如尼安德特人和丹尼索瓦人。
在现代人类研究中,染色体单倍群也被广泛用于人类遗传学、疾病研究和谱系分析等方面。
通过研究染色体单倍群的遗传变异,我们可以了解到一些与疾病风险相关的基因变异。
例如,一些与乳腺癌和前列腺癌相关的基因变异与染色体单倍群有关联,帮助科学家理解疾病发生和发展的机制。
此外,染色体单倍群还被用于确定个人的遗传谱系,追溯家族历史,帮助人们了解自己的祖先和亲属关系。
单倍群O单倍群O-M175起源时间约3万年前起源地东南亚与东亚上游单倍群NO下游单倍群O-MSY2.2、O-M268、O-M122对应突变M175、P186、P191、P196单倍群O-M175(英语:Haplogroup O-M175)是⼈类Y染⾊体DNA单倍群之⼀,主要分布于东亚与东南亚,源于单倍群NO。
起源单倍群O⾸次出现于距今约3万年前,出现地为东南亚或东亚,是单倍群NO-M214的两个下游单倍群之⼀(另⼀下游单倍群则是主要分布于欧亚⼤陆北部的单倍群N)。
分布东亚与东南亚的⼈⼝中80%⾄90%属于单倍群O,⽽在其他地区该单倍群则⼗分罕见。
在西伯利亚西部、西亚、欧洲、⾮洲⼤部分地区与美洲⼏乎没有该单倍群的存在,这些地区极少数的单倍群O可能是近期移民的结果。
由单倍群O分出的三个下游单倍群分别为O-MSY2.2、O-M268与O-M122。
其中,O-MSY2.2主要分布中国东南沿海地区、侗台族群、南岛族群等。
O-M268的主要⽀系O-M95常见于中国南⽅少数民族及中南半岛等地,另⼀⽀系O-M176则常见于朝鲜半岛与⽇本等地。
⽽O-M122则遍布东亚与东南亚,也是中国最为常见的单倍群。
系统树(O-M175)以下系统树主要基于ISOGG于2017年的分类(wer.12.244):O1(F265/M1354)O1a(旧O1) (M119)O1a1 (B384/Z23193)O1a1a (M307.1/P203.1)O1a1a1 (F446)O1a1a1a (F140)O1a1a1a1 (F78)O1a1a1a1a (F81)O1a1a1a1a1 (CTS2458)O1a1a1a1a1a (F533)O1a1a1a1a1a1 (F492)O1a1a1a1a1a1a (F656)O1a1a1a1a1a1a1 (A12440)O1a1a1a1a1a1a1a (A12439)O1a1a1a1a1a1a2 (A14788)O1a1a1a1a1a1a3 (F65)O1a1a1a1a1a1a4 (MF1068)O1a1a1a1a1a1a5 (Z23482)O1a1a1a1a1a1b (FGC66168)O1a1a1a1a1a1b1 (CTS11553)O1a1a1a1a1a1c (Y31266)O1a1a1a1a1a1c1 (Y31261)O1a1a1a1a1a1d (A12441)O1a1a1a2 (YP4610/Z39229)O1a1a1a2a (AM00330/AMM480/B386) O1a1a1a2a1 (AM00333/AMM483/B387) O1a1a1a2a1a (B388)O1a1a1a2b (SK1555)O1a1a1b (SK1568/Z23420)O1a1a1b1 (M101)O1a1a1b2 (Z23392)O1a1a1b2a (Z23442)O1a1a1b2a1 (SK1571)O1a1a2 (CTS52)O1a1a2a (CTS701)O1a1a2a1 (K644/Z23266)O1a1b (CTS5726)O1a2 (M110)O1a2a (F3288)O1a2a1 (B392)O1a2a1a (B393)O1a3 (Page109)O1b(旧O2) (M268)O1b1 (F2320)O1b1a (M1470)O1b1a1 (PK4)O1b1a1a(旧O2a) (M95)O1b1a1a1 (F1803/M1348)O1b1a1a1a (F1252)O1b1a1a1a1 (F2924)O1b1a1a1a1a (M111)O1b1a1a1a1a1 (F2758)O1b1a1a1a1a1a (Z24083)O1b1a1a1a1a1a1 (Z24089)O1b1a1a1a1a1a1a (F923)O1b1a1a1a1a1a1a1 (CTS2022)O1b1a1a1a1a1a1a1a (F1399)O1b1a1a1a1a1a1a1a1 (F2415)O1b1a1a1a1a1a1a2 (Z24131)O1b1a1a1a1a1a1a3 (Z24100)O1b1a1a1a1a1a1b (SK1627/Z24091) O1b1a1a1a1a1a1b1 (Z39410)O1b1a1a1a1a1a2 (Z24088)O1b1a1a1a1a2 (F2890)O1b1a1a1a1a2a (Z24048)O1b1a1a1a1a2a1 (Z24050)O1b1a1a1a1a2b (Z24014)O1b1a1a1a1b (CTS5854)O1b1a1a1a1b1 (Z23810)O1b1a1a1a1b1a (CTS7399)O1b1a1a1a1b1a1 (FGC19713/Y14026) O1b1a1a1a1b1a1a (Z23849)O1b1a1a1a1b1a1a1 (FGC61038)O1b1a1a1a1b1b (CTS651)O1b1a1a1a1b1b1 (CTS9884)O1b1a1a1a2a (SK1636)O1b1a1a1b (F789/M1283)O1b1a1a1b1 (FGC29900/Y9322/Z23667)O1b1a1a1b1a (B426/FGC29896/Y9033/Z23671) O1b1a1a1b1a1 (FGC29907/YP3930)O1b1a1a1b1a2 (B427/Z23680)O1b1a1a1b1b (Z39485)O1b1a1a1b1c (B418)O1b1a1a1b2 (SK1646)O1b1a1a2 (CTS350)O1b1a1a3 (Page103)O1b1a1b (F838)O1b1a1b1 (F1199)O1b1a2 (Page59)O1b1a2a (F993)O1b1a2a1 (F1759)O1b1a2a1a (CTS1127)O1b1a2b (F417/M1654)O1b1a2b1 (F840)O1b1a2b1a (F1127)O1b1a2b2 (CTS1451)O1b1a2c (CTS9996)O1b2(旧O2b) (P49, M176)O1b2a1 (CTS9259)O1b2a1a (F1204)O1b2a1a1 (CTS713)O1b2a1a1a (CTS1875)O1b2a1a1a1 (CTS10682)O1b2a1a1b (Z24598)O1b2a1a1c (CTS203)O1b2a1a2 (F2868)O1b2a1a2a (L682)O1b2a1a2a1 (CTS723)O1b2a1a2a1a (CTS7620)O1b2a1a2a1b (A12446)O1b2a1a2a1b1 (PH40)O1b2a1a2b (F940)O1b2a1a3 (CTS10687)O1b2a1a3a (CTS1215)O1b2a1b (CTS562)O1b2a2 (Page90)O2(O-M122)单倍群O-M122单倍群O-M122的等频率分布图起源时间约3万年前起源地中国或东南亚上游单倍群单倍群O-M175对应突变M122O-M122 (M122)O2a (M324)O2a1a1a (F1867/Page124)O2a1a1a1 (F852)O2a1a1a1a (F2266)O2a1a1a1a1 (L599)O2a1a1a1a1a (Z43961)O2a1a1a1a1a1 (Z43963)O2a1a1a1b (F854)O2a1a1a1b1 (Z43966)O2a1a1a1c (Page130)O2a1a1b (F915)O2a1a1b1 (F1478)O2a1a1b1a (PF5390)O2a1a1b1a1 (CTS1936)O2a1a1b1a1a (Z43975)O2a1a1b1a2 (FGC33994)O2a1b (M164)O2a1c (IMS-JST002611)O2a1c1 (F18)O2a1c1a (F117)O2a1c1a1 (F13)O2a1c1a1a (F11)O2a1c1a1a1 (F632)O2a1c1a1a1a (F110/M11115)O2a1c1a1a1a1 (F17)O2a1c1a1a1a1a (F377)O2a1c1a1a1a1a1 (F1095)O2a1c1a1a1a1a1a (F856)O2a1c1a1a1a1a1a1 (F1418)O2a1c1a1a1a1a1a2 (Z25097)O2a1c1a1a1a1a2 (CTS7501)O2a1c1a1a1a1b (F793)O2a1c1a1a1a2 (Y20951)O2a1c1a1a1a2a (Y20932)O2a1c1a1a2 (F38)O2a1c1a1a3 (F12)O2a1c1a1a4 (F930)O2a1c1a1a4a (F2685)O2a1c1a1a5 (F1365/M5420/PF1558) O2a1c1a1a5a (Y15976)O2a1c1a1a5a1 (Y16154)O2a1c1a1a5a1a (Y26383)O2a1c1a1a5a1a1 (SK1686)O2a1c1a1a5b (FGC54486)O2a1c1a1a5b1 (FGC54507)O2a1c1a1a6 (CTS12877)O2a1c1a1a6a (F2527)O2a1c1a1a6a1 (CTS5409)O2a1c1a1a6a2 (F2941)O2a1c1a1a7 (F723)O2a1c1a1a8 (CTS2107)O2a1c1a1a9 (SK1691)O2a1c1a1b (PH203)O2a1c1b (F449)O2a1c1b1a2 (F724)O2a1c1b2 (F1266)O2a1c1c (CTS498)O2a1c2 (FGC3750/SK1673)O2a2 (IMS-JST021354/P201)O2a2a (M188)O2a2a1 (F2588)O2a2a1a (CTS445)O2a2a1a1 (CTS201)O2a2a1a1a (M159/Page96)O2a2a1a2 (M7)O2a2a1a2a (F1276)O2a2a1a2a1 (CTS6489)O2a2a1a2a1a (F1275)O2a2a1a2a1a1 (M113)O2a2a1a2a1a2 (N5)O2a2a1a2a1a3 (Z25400)O2a2a1a2a2 (F1863)O2a2a1a2a2a (F1134)O2a2a1a2a2a1 (F1262)O2a2a1a2b (Y26403)O2a2a1b (F1837)O2a2a2 (F879)O2a2a2a (F1226)O2a2a2a1 (F2859)O2a2b (P164)O2a2b1 (M134)O2a2b1a (F450/M1667)O2a2b1a1 (M117/Page23)O2a2b1a1a (M133)O2a2b1a1a1 (F438)O2a2b1a1a1a (Y17728)O2a2b1a1a1a1 (F155)O2a2b1a1a1a1a (F813/M6539)O2a2b1a1a1a1a1 (Y20928)O2a2b1a1a1a2 (F1754)O2a2b1a1a1a2a (F2137)O2a2b1a1a1a2a1 (F1442)O2a2b1a1a1a2a1a (F1123)O2a2b1a1a1a2a1a1 (F1369)O2a2b1a1a1a2a2 (A16636)O2a2b1a1a1a3 (Z25907)O2a2b1a1a2 (FGC23469/Z25852) O2a2b1a1a2a (F310)O2a2b1a1a2a1 (F402)O2a2b1a1a2a1a (F1531)O2a2b1a1a3 (CTS7634)O2a2b1a1a3a (F317)O2a2b1a1a3a1 (F3039)O2a2b1a1a3a2 (Y29861)O2a2b1a1a3b (CTS5488)O2a2b1a1a4 (Z25853)O2a2b1a1a4a (CTS5492)O2a2b1a1a5 (CTS10738/M1707)O2a2b1a1a5a (CTS9678)O2a2b1a1a5a1 (Z39663)O2a2b1a1a5a2 (M1513)O2a2b1a1a5b (A9457)O2a2b1a1a5b1 (F17158)O2a2b1a1a6 (CTS4658)O2a2b1a1a6a (CTS5308)O2a2b1a1a6b (Z25928)O2a2b1a1a6b1 (SK1730/Z25982)O2a2b1a1a6b1a (Z26030)O2a2b1a1a6b1b (Z26010)O2a2b1a1a6b2 (A9462)O2a2b1a1a6b3 (B456)O2a2b1a1a7 (YP4864)O2a2b1a1a7a (Z44068)O2a2b1a1a7a1 (F5525/SK1748)O2a2b1a1a7b (Z44071)O2a2b1a1a8 (Z44091)O2a2b1a1a8a (Z44092)O2a2b1a1b (CTS4960)O2a2b1a2 (F114)O2a2b1a2a (F79)O2a2b1a2a1 (F46/Y15)O2a2b1a2a1a (FGC16847/Z26091)O2a2b1a2a1a1 (F48)O2a2b1a2a1a1a (F152)O2a2b1a2a1a1a1 (F2505)O2a2b1a2a1a1b (CTS3149)O2a2b1a2a1a2 (F242)O2a2b1a2a1a2a (CTS4266)O2a2b1a2a1a2a1 (Z26108)O2a2b1a2a1a2a1a (F2173)O2a2b1a2a1a3 (F2887)O2a2b1a2a1a3a (F3607)O2a2b1a2a1a3a1 (F3525)O2a2b1a2a1a3b (CTS3763)O2a2b1a2a1a3b1 (A9472)O2a2b1a2a1a3b2 (FGC16863/Y7110)O2a2b1a2a1a3b2a (L1360)O2a2b1a2a1a3b2a1 (FGC16889)O2a2b1a2a1a3b2b (SK1768/Y7112/Z26257) O2a2b1a2a1a3b2b1 (F4249)O2a2b1a2a1a3b2b1a (FGC23868)O2a2b1a2a1a3b2b2 (CTS335)O2a2b1a2a1b (CTS53)O2a2b1a2a1b1 (CTS6373)O2a2b1a2a1b1a (A9473)O2a2b1a2a1c (F3386)O2a2b1a2a1d (Y29828)O2a2b1a2a1d1 (F735)O2a2b1a2a1d1a (FGC34973)O2a2b1a2a1d1b (F1739)O2a2b1a2b1a1 (A16629)O2a2b1a2b1a2 (CTS682)O2a2b1a2b2 (F748)O2a2b1a2b2a (F728)O2a2b1a2c (Page101)O2a2b2 (AM01822/F3223)O2a2b2a (AM01856/F871)O2a2b2a1 (N7)O2a2b2a1a (F4110)O2a2b2a1a1 (F4068)O2a2b2a1a2 (SK1780)O2a2b2a1b (F4124)O2a2b2a1b1 (IMS-JST008425p6)O2a2b2a1b2 (BY15188)O2a2b2a1b2a (F16411)O2a2b2a2 (AM01845/F706)O2a2b2a2a (F717)O2a2b2a2a1 (F3612)O2a2b2a2a2 (SK1783)O2a2b2a2b (AM01847/B451)O2a2b2a2b1 (A17418)O2a2b2a2b2 (AM01756)O2a2b2a2b2a (B450)O2a2b2a2b2b (AM00472/B452)O2a2b2a2b2b1 (F18942)O2a2b2a2b2c (A16427)O2a2b2b (A16433)O2a2b2b1 (A16438)O2a2b2b1a (SK1775)O2a2b2b1a1 (SK1774)O2a2b2b1b (A16440)O2a3 (M300)O2a4 (M333)O2b (F742)O2b1 (F1150)O2b1a (F837)O2b1a1 (F1025)O2b2 (F1055)O2b2a (F3021)⼈类Y染⾊体DNA单倍型类群进化树Y染⾊体最近共同祖先AA1b A1a-TA1a A2-TA2A3BTB CTDE CFD E C FG H IJKIJ KI J LT K(xLT)L T M NO P SO N Q R各族群Y染⾊体单倍型类群 · 著名的Y染⾊体单倍型类群⼈。
Y染色体单倍群知识科普本文主要来源于wiki翻译自英文简介在人类遗传学中,人类Y染色体DNA单倍群是由男性Y染色体的DNA的非重组部分中的发生突变的单倍群。
许多人在一个单倍群中有着相似短串联重复序列(STRs)数量还存在着称为单核苷酸多态性(SNPs)的核苷酸突变。
人类Y染色体每世代大约会有两个突变。
Y-DNA单倍群是Y染色体进化系统树的主要分支,每个单倍群共有数百甚至数千个独特的突变。
在Y染色体最近缘的共同祖先(Y-MRCA,俗称Y染色体亚当)是最早的共同祖先(MRCA),他是目前所有现代人类的父系起源。
据估计,Y染色体亚当大约生活在236,000年前的非洲。
通过研究遗传瓶颈,大多数欧亚男人(来自非洲以外地区的男人)都是一个来自69,000年前的男人的后裔。
其他主要遗传瓶颈发生在大约50,000和5,000年前,后来大多数(欧亚/非洲以外)男人的祖先可以追溯到50,000年前的四位祖先。
Y-DNA单倍群发展树状图人类Y-DNA发展图和单倍群地区分布名词解释单倍群:单倍型类群是一组类似的单倍型单倍型:是单倍体基因型的简称,在遗传学上是指在同一染色体上进行共同遗传的多个基因座上等位基因的组合等位基因:是指位于一对同源染色体相同位置上控制同一性状不同形态的基因Aa中 A和a就是一对等位基因基因座:基因在染色体上所占的位置DNA重组:遗传物质的交换,复制等重新组合短串联重复序列(STR):串联重复序列是指以相对恒定的短序列为重复单位,首尾相接,串联连接形成的重复序列。
在人类基因组中,串联重复序列约占10%,主要分布在非编码区,少数位于编码区。
非编码区:是不能够转录为相应信使RNA,不能指导蛋白质合成(也就是不能编码蛋白质)的区段。
单核苷酸多态性(SNP):主要是指在基因组水平上由单个核苷酸的变异所引起的DNA序列多态性。
SNP在人类基因组中广泛存在,平均每300个碱基对中就有1个,估计其总数可达300万个甚至更多。
Y染色体DNA单倍群介绍1、Y-DNA单倍群人类Y染色体DNA单倍群由非重组DNA的Y染色体突变进行定义。
这种由许多人共享的突变称为单核苷酸多态性(SNP)。
人类Y染色体每一代大约积累两次突变。
Y-DNA单倍群的分支结构组成一个Y染色体进化树,有数百甚至数千的突变由这些不同的单倍群共享。
Y染色体的最近的共同祖先(most recent common ancestor MRCA),也被称为Y染色体亚当,是目前活着的男性的最近的男性共同祖先。
Y染色体亚当估计生活在大约236000年前的非洲。
通过研究其他瓶颈,所有欧亚大陆的人都是69000年前的一个男人的后裔。
之后一个主要的遗传瓶颈期发生在大约5000年前,今天大多数欧亚大陆的人可以追溯到5000年前的十二个祖先。
Y-DNA单倍群进化树单倍群 A & B 单倍群 A(M91)单倍群A是所有单倍群起源点。
现代所有单倍群都是单倍群A的后代,稀疏分布在非洲,主要集中在西南部的科伊桑人和尼罗河谷东北部人群。
单倍群 BT (M42,M94,M139,M299)约55000年前分,BT是单倍群A的分支单倍群B(M60)单倍群B主要分布于非洲,主要集中于俾格米人群。
详细树形图:见B单倍群文件夹单倍群 CT (P143)标识单倍群 CT的突变标记是M168和M294.包含单倍群D、E、C、F,可能88000年前在亚洲或非洲出现。
单倍群 C (M130)历史起源:C单倍群携带M130突变,来源于CF单倍群。
中国境内的C单倍群主要是C2(携带M217突变),占中国总人口比例大约为5%—10%。
其下游又可分为南北两大支,北支C2b(携带F1396突变),主要分布于蒙古族和满族等民族;南支C2c(携带F1067突变),几乎遍及全中国。
详细树形图:见C单倍群文件夹Haplogroup C (M130, M216) 分布在亚洲、大洋洲和北美等o Haplogroup C1 (F3393/Z1426)▪Haplogroup C1a (CTS11043)▪Haplogroup C1a1 (M8, M105, M131) 日本低频分布▪Haplogroup C1a2 (V20) 欧洲和尼泊尔低频分布▪Haplogroup C1b (F1370, Z16480)▪Haplogroup C1b1 (AM00694/K281)▪Haplogroup C1b1a (B66/Z16458)▪Haplogroup C1b1a1 (M356) 印度低频分布, 阿拉伯半岛和中国北部▪Haplogroup C1b2 (B477/Z31885)▪Haplogroup C1b2a (M38) 分布在印度尼西亚,新几内亚岛,美拉尼西亚,密克罗尼西亚,和玻利尼西亚▪Haplogroup C1b2b (M347, P309) 澳洲土著o Haplogroup C2 (M217, P44) 分布在欧亚大陆和北美,特别是在蒙古人,哈萨克人,通古斯人,西伯利亚人,和Na-Dené-speaking语民族单倍群DE(M1,M145,M203)约65000年前分离单倍群D (M174)详细树形图:见D单倍群文件夹∙Haplogroup D (M174) 分布在日本、中国(特别分布于西藏)和安达曼岛o D1 (CTS11577)▪D1a (Z27276, Z27283, Z29263)▪Haplogroup D1a1 (M15) 主要分布在西藏、羌族、彝族和苗瑶语人群▪Haplogroup D1a2 (P99) 主要分布在西藏、羌族、纳西族、突厥部落▪Haplogroup D1b (M55, M57, M64.1, M179, P12, P37.1, P41.1 (M359.1), 12f2.2) 主要在日本o D2 (L1366, L1378, M226.2) 菲律宾、麦克坦岛Haplogroup E (M96)详细树形图:见E倍群文件夹∙Haplogroup E (M40, M96) 分布在非洲、中东和欧洲o Haplogroup E1 (P147)▪Haplogroup E1a (M33, M132) 旧称E1▪Haplogroup E1b (P177)▪Haplogroup E1b1 (P2, DYS391p); 旧称E3▪Haplogroup E1b1a (V38) 非洲尼日尔-刚果语人群; 旧称E3a▪Haplogroup E1b1b (M215) 非洲之角,北非、中东和欧洲地中海地区; 旧称E3bo Haplogroup E2 (M75)Haplogroup F (M89)单倍群F和后代迁徙图单倍群F和后代构成了目前世界人口的90%,几乎都分布在撒哈拉以南非洲地区之外。
最早出现在印度西北部地区。
IJ的突变对应于45000年前的一个中东或南亚地区的移民浪潮,随后迁移到欧洲(克罗马努)。
详细树形图:见F倍群文件夹Haplogroup G (M201)历史起源:Haplogroup G(M201) 起源于48,000年前,最近的共同祖先大概生活在26,000年前的中东,在新石器时代扩散到欧洲。
在欧亚地区很多种族都有发现,在高加索、伊朗、安纳托利亚和黎凡特地区常见。
在几乎所有的欧洲国家发现,最常见于高加索、罗马尼亚东南部、希腊、意大利、西班牙、葡萄牙、蒂罗尔州、波西米亚,一些地中海岛屿的浓度最高,在欧洲北部罕见。
G-M201也在中国西北部的印度,巴基斯坦,斯里兰卡,马来西亚,北非少量发现。
详细树形图:见G倍群文件夹Haplogroup H (M69)历史起源:单倍群H(M69)可能出现在南亚,约48000年前,目前仍然普遍存在,包括H1(M69)和H3(z5857)。
H2(P96)自从新石器时代在欧洲出现,h1a1随着中世纪罗姆人(吉普赛人)移民向西方传播。
详细树形图:见H倍群文件夹Haplogroup I (M170)Haplogroup I (M170, M258) 主要分布在欧洲和高加索,单倍群I(M170)的先祖是中东氏族M89的一部分,他们向西北的巴尔干迁移,后来扩散到欧洲中部,在大约2.1万至2.8万年前,这些群体在欧洲西部创造了格拉维特文化。
∙Haplogroup I1 (M253) 主要分布于欧洲北部∙Haplogroup I2 (P215) 主要分布在欧洲东南部和撒丁岛,以I2b1(m223)形式存在。
详细树形图:见I倍群文件夹Haplogroup J (M304)单倍群 J(m304,S6、S34,S35)主要是在中东和欧洲的东南部发现的。
详细树形图:见J倍群文件夹∙Haplogroup J* (J-M304*) 主要在索科特拉岛,岛外罕见.o Haplogroup J1 (M267) 主要发现于达吉斯坦的东北高加索人,中东、埃塞俄比亚和北非的闪米特语人群,也低频发现于欧洲地中海.o Haplogroup J2 (M172) 主要发现于闪米特人,安纳托利亚、希腊、巴尔干、意大利、伊朗、南亚/中亚和高加索地区。
Haplogroup K (M9)Haplogroup K(M9)遍布欧亚大陆、大洋洲和美洲土著。
详细树形图:见K倍群文件夹K(xLT K2a、K2b)–即K*,k2c,K2d或K2e, 主要在美拉尼西亚,澳洲土著,印度,波利尼西亚和东南亚岛屿。
Haplogroups L & T (K1)Haplogroup L (M20) 在南亚,中亚,西南亚和地中海.Haplogroup T (M184, M70, M193, M272) 在非洲之角高水平分布,另外也分布在南亚部分地区,中东和地中海. T-M184 低频出现在Sciaccensi, Stilfser, Fulbe, 埃及,Omanis, Sephardi Jews, Ibizans (Eivissencs), Toubou. 也在地中海和南亚其他地区的低频处被发现。
详细树形图:见L倍群文件夹Haplogroup K2 (K-M526)目前检测到的携带K2*的一位男性是土著澳大利亚人。
研究显示原住民的澳大利亚男性由27%携带K2*,而其他人携带K2的一个亚型。
Haplogroups K2a, K2a1, NO & NO1Haplogroup NHaplogroup N (M231) 主要在欧亚大陆北部,特别是在乌拉尔语系人群。
N系单倍群起源于亚洲东部,向西传播到西伯利亚,向北传播到乌拉尔语人群。
详细树形图:见N倍群文件夹Haplogroup OHaplogroup O (M175) 主要在东亚、东南亚和南太平洋地区。
Haplogroup O1 (F265/M1354, CTS2866, F75/M1297, F429/M1415, F465/M1422)o Haplogroup O1a (M119, CTS31, F589/Page20, L246, L466) 分布在华东、华南、台湾和东南亚南洋,特别是在南岛语族和侗傣语族人群。
o Haplogroup O2 (P31, M268)▪Haplogroup O2a1 (M95) 在日本,华南,台湾,东南亚和印度次大陆,尤其是南岛和侗傣语人群,以及马来人和印尼人。
▪Haplogroup O2b (SRY465, M176) 分布在日本,韩国,满洲里和东南亚。
o Haplogroup O3 (M122) 整个东亚地区,包括东南亚,南太平洋群岛和波利尼西亚。
▪O3a1c-002611:汉族常见类型,占汉族15%以上▪O3a2b-M7:苗瑶族群特征类型,通常占汉族5%以下▪O3a2c1-M134:汉族30%左右,广泛分布于东亚、东南亚▪O3a2c1a-M117:汉族和藏缅语族的特征类型,汉族15%以上详细树形图:见O倍群文件夹Haplogroups K2b1, M & S目前没有发现底层k2b1*的例子。
携带亚型k2b1的人主要是在巴布亚人,密克罗尼西亚人、波利尼西亚人和澳大利亚土著人。
主要的两个亚单倍群:∙Haplogroup S (B254) also known as K2b1a: 见于巴布亚新几内亚。
∙Haplogroup M (P256) also known as K2b1b: 见于新几内亚和美拉尼西亚。
Haplogroup P (K2b2)单倍群 P(p295)有两个主要分支:P1(p-m45)和极为罕见的P2(p-b253)。
P1*和P2、P* 仅在菲律宾吕宋岛同时发现,特别是P*和P1*在吕宋岛的Aeta人之间较显著的发现率。
同时,P1*在西伯利亚东部和中亚生活的人中也较普遍,它也在东南亚和南亚低频发现。