Cadmium inhibits the induction of high-affinity nitrate uptake
- 格式:pdf
- 大小:463.14 KB
- 文档页数:12

菜豆多聚泛肽基因在重金属胁迫下的表达_英文_ () 植物学报 1999 , 41 10:1052,1057Acta B ota nica Si nicaΞ菜豆多聚泛肽基因在重金属胁迫下的表达柴团耀张玉秀( )中国科学技术大学研究生院生物部北京 100039( ) 摘要差别筛选 HgCl胁迫的菜豆 Phaseolus vulgaris L . 幼苗叶片 cDNA 库 , 分离出两个重金属胁迫相应基因 2 ( ) PvS R5 和 PvS R51 Phaseolus vulgaris stress- related gene片段。
cDNA 和氨基酸序列分析表明 PvS R5 和 PvS R51 分别编码一种多聚泛肽。
Northern blot 分析表明多聚泛肽是组成性表达蛋白 ,主要在根中表达 ,叶片和茎中表达较少 ; Hg 、Cd 、Cu 和 Zn 等重金属、高温和水杨酸能强烈地刺激其在叶片中的表达 ,而受伤几乎没有影响。
推测多聚泛肽在抵抗重金属胁迫和提高植物的抗逆性方面有重要作用。
关键词重金属 ,多聚泛肽 ,菜豆Expression Analysis of Polyubiquitin Genes from BeanΞ in Response to Heavy MetalsCHAI Tuan- Yao ZHANG Yu- Xiu( )Department of Biology , Graduate School of University of Science and Technology of China , The Chinese Academy of Sciences , Beijing 100039( Abstract Using differential screening of a leaf cDNA library prepared from a bean cultivar Phaseolus vul2) garis L . cv. Saxaexposed to HgCl, the authors have isolated and characterized two heavy metal- regulated 2( ) cDNA fragments , designated as PvS R5 and PvS R51 Phaseolus vulgaris stress- related gene. The sequencesof the cDNA inserts and homological analysis showed that both PvS R5 and PvS R51 encode a polyubiquitin re2spectively. The polyubiquitin genes were constitutively expressed in roots but weakly expressed in stems andleaves. Northern blot analysis revealed a low level of transcripts of polyubiquitin in unstressed bean leaves , butthe gene expression was strongly stimulated by heavy metals , elevated temperature and salicylic acid , whereaswounding had almost no effect . These suggested that polyubiquitin might play important roles in resistance toheavy metals and various environmental stresses.Key words Heavy metals ,Polyubiquitin , Phaseolus vulgarisUbiquitin is a 76 amino acid protein highly con2 mal protein , the ubiquitin- dependent pathway controls the served in all eukaryotes. The ubiquitins are encoded by levels of many key cell regulators , i . e . phytochrome andcyclin. So it plays major roles in various biological pro2 two different gene families , polyubiquitin genes and ubiq2 1 ,2 ) ( uitinextension protein genes UbEP . Polyubiquitin cesses, including DNArepair , transcription , signaltransduction , the cell cycle control , apoptosis and stress genes contain several direct repeats of the ubiquitin codingunit . UbEP genes contain a single ubiquitin coding unit responses. The expression of polyubiquitin genes can be 3 ,4 fused in frame to the coding region for a small protein as2 enhanced by HgClin maize and tobacco , we report 2sociated with ribosomes. Ubiquitin monomer in the cyto2 here the identification and characterization of polyubiqui2 plasm can be covalently attached to other proteins via a tin cDNA from bean and gene expression in response toheavy metals. Furthermore , the mechanism of plant resis2 multistep enzymatic process. The linkage occurs at specif2tance to heavy metals is discussed. ic lysine residues of the acceptor protein with formation ofan isopeptide bond between the carboxyl group of the C- 1 Materials and Methods εterminal glycine of ubiquitin and- NHgroup s of the ly2 2 sine side- chains of the target protein. The conjugation of 111Plant materials and stress conditions ubiquitin to protein may alter their stability or serve as a ( ) Bean Phaseolus vulgaris L . cv. Saxaseeds were recognition signal for proteolysis by the 26 S proteo2 surface- sterilized with a 2 . 5 % calcium hypochloride solu2 1 some .In addition to removing the denatured or abnor2 tion for 10 min , rinsed several times with distilled water() Ξ Supported by the“863”Grant for Youth and the NationalNatural Science Foundation of China No . 39870078. Received : 1999- 01- 14 Revised : 1999- 05- 16and then imbibed in sterile water for 16 h. Germination signal with the control probe . The two clones were sub2 ?after which the young plants were trans2 occurred at 22 ( ) cloned into pBluescript KS + plasmid vector and se2 ferred onto a liquid culture medium containing KNO2 3 quenced by the dideoxy method.( ) mmol/ L , Ca NO2 . 5 mmol/ L , MgSO1 mmol/ L , 3 2 4 113Northern blot analysisKHPO1 mmol/ L and Fe 2 . 8 mg/ L , Mn 0 . 55 mg/ L , 2 4 RNA samples were prepared and analyzed according 6 Zn 0 . 65 mg/ L , Cu 0 . 06 mg/L , B 0 . 32 mg/ L , Mo 0 . to standard protocols, by electrophoresis on 1 . 2 % a2 02 mg/ L . Plants were grown in a growth chamber with a garose-formaldehyde gels , transferring onto Hybond N photoperiod of 16 h at 22 ? during the day and 18 ? membranes , and hybridizing for 24 hin the presence of 6 during the night and a photosynthetic photon flux density ( ×SSC 1 ×SSC : NaCl 0 . 15 mol/ L , CHNaO0 . 15 6 537 32 - 2 - 1 ) μmol/ L- 50 % formamide at 42 ? to P- cDNA probes. When the two primary leaves of 150 mol m? s? .Hybridization was followed by three washes for 15 min were well expanded , plants were stressed by adding one ofeach at 42 ?in 2 ×SSC. Finally , the membrane was ex2the following metals : HgCl, CdCl, CuSOor ZnSOat a 2 2 4 4μposed to X- ray film for autoradiography at - 80 ?.final concentration of 100mol/ L . For other experiments() plants were grown in plots in soilunder the same light- 2 Results dark and temperature conditions. Various types of abiotic PvSR51 encode a polyubiquitin re2 2 . 1 PvSR5 and stress were applied asfollows : leaves were sprayed with a spectively ( ) 0 . 2 % W/ VHgClor CdCl, or 10 mmol/ L salicylic 2 2 analysis showed that PvS R5 clone con2 Sequence ( ) acid SA solution. For heat- shock treatment , plants ( ) tained a 790 bp insert Fig. 1A. Northern blot revealed were placed at42 ? for 4 h , while wounding was per2 7 that PvS R5 mRNA has a size of 1 200 nucleotides, in2 formed by dusting the leaves with celite and gentle rub2 dicating that the cDNA was not full- length. PvS R5 cDNA bing. Leaf tissue was harvested at various time points after contains more than two repeats of ubiquitin coding unit stress treatment . and lacks the 5′end coding sequence , leucine which is 2 bean cD NA li12 Construction and screening of a 1unique non- ubquitin residue at the end of the last ubiqui2 brary tin repeat . Total RNA extraction was done using the phenol/ PvS R51 cDNA is also partial fragment that lacks the 5 chloroform/ isoamylalcohol technique. Polyadenylated 5′coding region.It encodes more than four ubiquitin re2mRNAs were obtained by chromatographing total RNA peats , the terminal repeat contains another additional ( ) twice on oligo- dTcellulose as described by Sambrook ( )amino acid , phenylalanine Fig. 1B. Repeats of 228 nu2 6 et al . Double- stranded cDNA was synthesized from cleotides encoding ubiquitin monomers were aligned below polyadenylated RNA isolated from bean plantsharvested 6 the first complete one taken as an arbitrary reference . Ho2 h after spraying with mercuric choloride solution using the mologies were found to range between 80 % to 90 %. Pharmacia Biotech kit . The cDNA library was constructed There are no homologies in the3′untranslated region be2 λin the bacteriophagegt 10 cloning vector . tween PvS R5 and PvS R51 genes. Therefore , PvS R51 en2 The cDNA clones were placed at a density of about codes a polyubiquitin that isdifferent from PvS R5 . ( ) 1 000 plaques/ dish diameter 9 cm. Duplicate sets of 2 . 2 Expression of PvSR5 gene in various tissues of nitrocellulose filters containing recombinant phage plaques young bean plants 32 were screened with [ P - labelled cDNA probes , pre2 PvS R5 gene is highly expressed in root but weakly pared of using polyadenylated RNAs extracted from mer2 ( ) expressed in stems and leaf tissue Fig. 2. The same curic chloride- treated or untreated bean leaves. Differen2 ( pattern was observed on PvS R51 gene results not tial screening permitted the isolation of clones that were ) shown, demonstrating that polyubiquitin genes are con2 only expressed in mercury- treated plants or that were ex2 stitutively expressed in different tissues. pressed at higher levels in stressed plants than in control PvS R5 gene expres2 Effect of heavy metals on 2 . 3 plants. Both types of clones were considered to be heavy- sion in primary leaves( metal stress- related. 32 positive clones Phaseolus vul2 Theexpression of polyubiquitin gene was studied in ) garis stress- related gene , PvS R were isolated by screen2 response to several metal stresses. More than one hy2 ing of the cDNA library , in which PvS R5 and PvS R51 bridization signals are observed in Fig. 3 , as PvS R5 clones gave a strong hybridization signal with the cDNA probe contains partialubiquitin coding sequence that is probe obtained from mercury- treated plants and a weak植物学报 1054 41 卷() ()Fig. 1 Nucleotide sequence and deduced amino acid sequence of PvS R5 cDNA Aand PvS R51 cDNA B A. The initiation amino acid of the ubiquitin repeats is boxed , asterisk indicates the stop codon. GenBank access number for PvS R5 is U77940. B. The stop codon is indicated by an asterisk and the putative polyadenylation signal is underlined.Nucleotide sequences of the ubiq2 uitin repeats are aligned. Identical nucleotides are indicated by dots. Amino acid sequences are given in the one- letter code below the last u2 biquitin repeat . GenBank access number for PvS R51 is U77939.levels in the leaves , but with a different kinetics of induc2tion from that obtained with sprayed leaves. When mer2curic chloride was absorbed by the roots , PvS R5 mRNAstarted to accumulate at 9 h after the onset of treatment ,( ) reached a maximum at 48 h and then declined Fig. 3C.The transcript level continued to increase from 9 h afterthe onset of treatment until a maximum was reached 48 h( ) later when cadmium was absorbed by the roots Fig. 3D. Root- absorbed copper also stimulated the PvS R5 expres2 Fig. 2 PvS R5 gene expression in various tissues of Phaseolus vul2 garis seedlings sion , the transcripts reached a maximum after 24 h , ( μ) Total RNAs 10 g/ laneextracted from the various tissues of 12( ) whereas zinc had alittle effect Fig. 3 E , F. day-old plants were separated on formaldehyde- agarose gels , trans2 2 . 4 PvSR5 gene regulation by otherf orms of a biotic ferred onto membranes and hybridized with PvS R5 cDNA. R. Root ;S. Stem ; L . Primary leaves. stress Spraying with SA resulted in a large increase in tran2 highly homologous to the other ubiquitin gene . As shown script levels , reached a maximum at 3 h and then de2 in Fig.3A and 3B , the amounts of PvS R5 transcripts in2 ) ( ) (clined Fig. 4A. High temperature 42 ?also stimu2 creased rapidly , reached a maximum at3 h and then de2 ( ) lated strongly PvS R5 gene expression Fig. 4B, PvSR5 creased gradually when the leaves were sprayed with mer2 ( mRNA reached a maximum at 1 h during the stress 4 curic chloride or cadmium chloride solution. When the ) h, and then decreased after plants were returned to nor2 mercuric chloride or cadmium chloride was added to the mal temperature condition. Wounding had almost no effect liquid culture medium instead of being sprayed onto the ( ) PvS R5 gene expression Fig. 4C. on plants , PvS R5 mRNA also accumulated above the basalFig. 3 Northern blots analysis showing the levels of PvS R5 mRNA in bean leaves treated with various metals () () The time course of accumulation of PvS R5 transcripts after the seedlings were treated byvaporization of HgClAand CdClB, or by root- 2 2 ( ) () ( ) ( ) absorption of HgClC, CdClD, CuSOE, and ZnSOF. 2 2 4 4encode a polyubiquitin respectively. Both PvS R5 and 3 Discussion PvS R51 cDNA are incomplete at their 5′end probablySequence analysis showed that PvS R5 and PvS R51 because of inverted repeats in coding regions of ubiquitin genes contained more than two repeats of the ubiquitin mRNAs forming snap back loop structures which are self- 4 coding unit . The additional amino acid residues of both primed during cDNA synthesis. PvS R5 mRNA prefer2 genes encoded are different , PvS R5 terminates at entially accumulated in the root of bean seedling indicat2 leucine , whereas the phenylalanine is found at the C- ter2 ing that polyubiquitin gene has tissue- specific expression. minal of PvS R5 , suggesting that PvS R5 and PvS R51植物学报 1056 41 卷stimulate the expression of PvS R5 , but Cu and Zn had a( ) little effect Fig. 3, demonstrating that Hg and Cd ions are more toxic than Cu and Zn to living cell .PvS R5 gene responds not only to heavy metals , butalso to other stresses such as elevated temperature , 13 12 and virus infection, indicating that the ubiqui2 UVtin pathway has important roles in eliminating the damagedproteins induced by various stresses and maintaining thecell structure and function. Transgenic rice experimentsshowed that polyubiquitin gene expression was limited tothe region exposed to heat stress and/ or wounding , oraround the necrotic lesion induced by pathogen infection , 14 but rather not regulated systemically. This implied thatpolyubiquitins have key roles in the processes of the localdefence reaction. SA is a natural signal in the inductionof defense responses , which accumulates upon ozone or 15 Fig. 4 Northern blot analysis showing the time course of PvS R5 UV treatment , as well as pathogen attack. Exogenous accumulation in bean leaves under various abiotic stresses SA can positively regulate the gene expression of PvS R5 , The arrow on panel B indicates the transfer of the stressed plants to leading to the conclusion that SA might be involved in the normal growth conditions and the time points behind the arrows the duration of recovery. A. Salicylic acid ; B. Heat stress ; C. pathway of polyubiquitin mRNA synthesis in response to Wounding. 16 stresses. Chen et al . proposed that systemically ac2quired resistance signaling is mediated by an accumulation PvS R5 transcripts was detected in un2 A low level ofof HO, because SA specifically binds to catalase and ( ) stressed bean leaves Fig. 3, but the gene expression was 22 2 + 2 + inhibits its activity , leading to an elevation of HOlevel . ( 22 strongly stimulated by heavy metals Hg, Cdand 2 + Furthermore , HOinduces expression of defense- related 22 ) Cu. The datum is consistent with the responses of genes. A rapid and transient release of active oxygen polyubiquitin genes to mercuric stress in maize and tobac2 9 3 ,4 () species AOScan be induced by heavy metals, UV , , suggesting that polyubiquitin may playan impor2 co 16 17 pathogen infection, SA and heat stress. These re2tant role in resistance to heavy metals. Heavy metal ions sults suggest that PvS R5 gene expression was possibly in2 can bind to protein sulfhydryl group s , leading to deficien2volved in the regulation of AOS in response to stresses , cy of essential ions and destruction of the enzyme struc2 8 ,9 and there may be a common signal transduction pathway ture . Other toxicity mode is oxidative damage by free 9 ,10for various stresses in the regulation of PvS R5 gene ex2 radicals generated by metal redox cycling . Eventual2 pression. ly , the two processes result in accumulating huge amount of denatured and damaged proteins by metals in the cell .References In eukaryotes , an important selective proteolysis pathway for the elimination of abnormal proteins that are generated Belknap W R , Garbarino J E. The role of ubiquitin in plant 1 senescence and stress responses. Trends Plant Sci , 1996 , 1 : under normal or stress conditions is ATP- dependent and 2 331,335 mediated by ubiquitin system. The ubiquitin monomers generated by UbEP genes act as a molecular“chaperone” Tanaka K. Proteasones : Structure and biology. J Biochem , 21998 , 123 : 195,204in facilitating ribosome assembly. In contrast , the polyu2 Didierjean L , Frendo P , Nasser W , Geneviève G , Marivet J , 3 biquitin genes would provide monomers for general utiliza2 Burkard G. Heavy metal- responsive genes in maize : Identifi2 1 ,2 tion inubiquitin- mediated processes in cell. In cation and comparison of theirexpression upon various forms of yeast , the expressions ofpolyubiquitin and ubiquitin- con2 abiotic stress. Planta , 1996 , 199 : 1,8jugating enzyme genes are activated in response to cadmi2 4 Genschik P , Parmentier Y , Durr A , Marbach J , Criqui M C.um exposure and the mutants deficient in specific ubiqui2 Ubiquitin genes are differentially expressed in protoplast- de2rived cultures of Nicotiana sylvestris and in response to varioustin- conjugating enzymes are hypersensitive to cadmium , stresses. Plant Mol Biol , 1992 , 20 : 897,910 proposed that cadmium resistance is meditated partially by 11 Ragueh F , Lescure N , Roby D , Marco Y. Gene expression in 5 ubiquitin pathway. PvS R5 mRNA levels are increased Nicotiana tabacum in response to compatible and incompatible upon treatment with metals , implying that enormous ubiq2 isolates of Pseudomonas solanacearum . Physiol Mol Plant uitin monomers are required for degrading the abnormal Pathol , 1989 , 35 : 23,33 proteins produced by metals. Hg and Cd can strongly 6 Sambrook J , Fritsch E F , Maniatis T. Molecular Cloning : A() ()Laboratory Manual . 2nd ed. New York : Cold Spring Harbor 生物物理学报,1998 , 14 : 767,771 in Chinese() ( ) 13 Zhang Y- X张玉秀, Chai T- Y柴团耀. Effects of alfalfa Laboratory Press , 1989.) ( ) ( 7 柴团耀 , Zhang Y- X 张玉秀 , Burkard G. Chai T- Y mosaic virus infection on the expression of stress- responsiveHeavy metal- responsive genes in bean : cloning of cDNAs and ()genes in bean. Acta Hort Sin 园艺学报,1998 , 25 : 399,(()gene expression analysis. Acta Phytophysiol Sin 植物生理学 401 in Chinese ) ()14 Takimoto I , Christensen A H , Quail P H , Uchimiya H , Toki 报, 1998 , 24 : 399,404 in ChineseS. Non- systemic expression of a stress- responsive maize polyu2 8 Van Assche F , Clijsters H. Effects of metal on enzyme activity in plants. Plant Cell Environ , 1990 , 13 : 195,206 () biquitin gene Ubi-1in transgenic rice plants. Plant Mol Bi2 ( ) ( ) Zhang Y- X 张玉秀 , Chai T- Y 柴团耀 , Burkard G. ol , 1994 , 26 : 1007,1012 9 (Heavymetal tolerance mechanisms in plants. Acta B ot Sin 植 Ec Key- Kaltenbach H , Kiefer E , Grosskopf E , Ernst D , San2 15 ) ()物学报, 1999 , 41 : 453,457 in Chinese dermann H. Differential transcript induction of parsley patho2 González A , Steffen K L , Lynch J P. Light and excess man2genesis- related proteins and of a small heat shockprotein by o2 10 ganese implications for oxidative stress in common bean. Plant zone and heat shock. Plant Mol Biol , 1997 , 33 : 343,350 Physiol , 1998 , 118 : 493,50816 Chen Z , Silva H , Klessig D F. Active oxygen species in the J ungmann J , Reins H A , Schobert C , J entsch S. Resistance to 11 induction of plant systemic acquired resistance by salicylic cadmium mediated by ubiquitin- dependent proteolysis. N a2 acid. Science , 1993 , 262 : 1883,1886 ture , 1993 , 361 : 369,371 17Dat J F , Lopez- Delgado H , Foyer C H , Scott I M. Parallel () () (Chai T- Y柴团耀, Zhang Y- X张玉秀, Zhang Z- D 张正 changes in HOand catalaseduring thermotolerance induced 12 22 ) 东. Effects of elevated ultraviolet radiation on the stress- re2 by salicylic acid or heat acclimation in mustard seedlings. sponsive gene expression of bean seedling. Acta Biophys Sin Plant Physiol , 1998 , 116 : 1351,1357。

文章编号:1673-887X(2023)02-0083-03水环境中镉污染处理的研究进展吴泉,罗词安(上饶幼儿师范高等专科学校,江西上饶334000)摘要镉存在于自然界中的硫镉矿中,属于一种有毒的金属元素,具有降解难、毒性低、稳定性高、可蓄积的特征,水、土壤、动植物中都分布着一些镉,该重金属会对人的健康造成很大的危害。
当镉进入到人体后,往往需要较长的时间才能排出体外,通常人肾皮质的镉生物学半衰期为10~30年。
近年来我国各地时常发生镉污染事故,给当地居民的正常生产生活造成了极为严重的影响,我国政府和社会各界也非常关注镉的生产安全及其对环境影响。
文章主要阐述水环境中镉污染处理技术。
关键词镉污染;水环境;研究进展中图分类号X523文献标志码A doi:10.3969/j.issn.1673-887X.2023.02.025Progress in the Treatment of Cadmium Pollution in Water EnvironmentWu Quan,Luo Ci'an(Shangrao Preschool Teachers College,Shangrao334000,Jiangxi,China)Abstract:Cadmium,which exists in cadmium sulfide ore in nature,is a toxic metal element,which has the characteristics of diffi‐cult degradation,low toxicity,high stability and accumulation,and some cadmium is distributed in water,soil,animals and plants, this heavy metal will cause great harm to human health.When cadmium enters the human body,it often takes a long time to be ex‐creted from the ually,the biological half-life of cadmium in human renal cortex is10~30years.In recent years,cadmium pollution accidents often occur in various parts of China,which have a very serious impact on the normal production and life of local residents.The government and all sectors of society are also very concerned about the production safety of cadmium and its impact on the environment.This paper mainly described the treatment technology of cadmium pollution in water environment.Key words:admium pollution,water environment,research progress1镉污染的概述如今,美国毒物和疾病登记署(ATSDR)将镉列入第6位危害人类健康的有毒物质,且在全球性危险化学物质中,联合国环境规划署也将镉列为首位的有毒重金属;镉也是国际职业卫生重金属委员会和世界卫生组织中重点研究的环境、食品污染物。
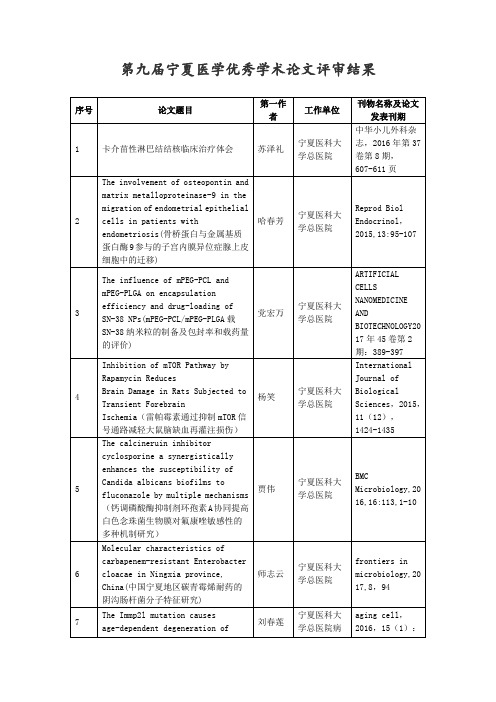
第九届宁夏医学优秀学术论文评审结果序号论文题目第一作者工作单位刊物名称及论文发表刊期1 卡介苗性淋巴结结核临床治疗体会苏泽礼宁夏医科大学总医院中华小儿外科杂志,2016年第37卷第8期,607-611页2 The involvement of osteopontin andmatrix metalloproteinase-9 in themigration of endometrial epithelialcells in patients withendometriosis(骨桥蛋白与金属基质蛋白酶9参与的子宫内膜异位症腺上皮细胞中的迁移)哈春芳宁夏医科大学总医院Reprod BiolEndocrinol,2015,13:95-1073 The influence of mPEG-PCL andmPEG-PLGA on encapsulationefficiency and drug-loading ofSN-38 NPs(mPEG-PCL/mPEG-PLGA载SN-38纳米粒的制备及包封率和载药量的评价)党宏万宁夏医科大学总医院ARTIFICIALCELLSNANOMEDICINEANDBIOTECHNOLOGY2017年45卷第2期:389-3974 Inhibition of mTOR Pathway byRapamycin ReducesBrain Damage in Rats Subjected toTransient ForebrainIschemia(雷帕霉素通过抑制mTOR信号通路减轻大鼠脑缺血再灌注损伤)杨笑宁夏医科大学总医院InternationalJournal ofBiologicalSciences,2015,11(12),1424-14355 The calcineruin inhibitorcyclosporine a synergisticallyenhances the susceptibility ofCandida albicans biofilms tofluconazole by multiple mechanisms(钙调磷酸酶抑制剂环孢素A协同提高白色念珠菌生物膜对氟康唑敏感性的多种机制研究)贾伟宁夏医科大学总医院BMCMicrobiology,2016,16:113,1-106 Molecular characteristics ofcarbapenem-resistant Enterobactercloacae in Ningxia province,China(中国宁夏地区碳青霉烯耐药的阴沟肠杆菌分子特征研究)师志云宁夏医科大学总医院frontiers inmicrobiology,2017,8,947 The Immp2l mutation causesage-dependent degeneration of刘春莲宁夏医科大学总医院病aging cell,2016,15(1):cerebellar granule neuronsprevented by antioxidant treatment(抗氧化治疗对Immp21突变小鼠的小脑颗粒神经元凋亡的保护作用)167-1768 The therapeutic response of CDDO-Mein the esophageal squamous cellcarcinoma (ESCC) cells is mediatedby CaMKIIα(CaMKIIα参与调控CDDO-Me治疗食管鳞癌效应研究)王艳阳宁夏医科大学总医院美国转化研究杂志2016;8(4),1695-17079 Enhanced encapsulation andbioavailability of breviscapine inPLGA microparticles by nanocrystaland water-soluble polymer templatetechniques(基于纳米结晶与凝胶模板技术提高灯盏花素-PLGA微粒包封率及生物利用度)王虹宁夏医科大学总医院Eur J PharmBiopharm,2017,115:177-18510 Period2 downregulation inhibitsglioma cell apoptosis by activatingthe MDM2-TP53 pathway(Period2下调通过激活MDM2-TP53信号通路抑制胶质瘤细胞凋亡)牛占锋宁夏医科大学总医院Oncotarget,Vol. 7, No. 1911 Antibodies against C1q Are aValuable Serological Marker forIdentification of Systemic LupusErythematosus Patients with ActiveLupus Nephritis(抗C1q抗体可作为鉴别诊断系统性红斑狼疮肾炎的血清学标志之一)池淑红宁夏医科大学总医院DiseaseMarkers,2015,450351:1-1112 Wnt signaling as potentialtherapeutic targetIn lung cancer(Wnt信号作为肺癌潜在治疗的靶点)杨佳丽宁夏医科大学总医院Expert Opinionon TherapeuticTargets,2016,20(8):999-101513 Wnt5a Increases Properties of LungCancer Stem Cells and Resistance toCisplatin through Activation ofWnt5a/PKC Signaling Pathway(Wnt5a通过激活Wnt5a/PKC信号途径增加肺癌干细胞的特性及顺铂耐药性)杨佳丽宁夏医科大学综医院Stem cellinternational2016,1690896,16 pages14 Dendritic and langerhans cellsrespond to Aβ peptidesdifferently:implication for ADimmunotherapy(树突状细胞和朗格罕式细胞对β-淀粉样蛋白作用的反应不同:对阿尔兹海默病免疫治疗的意义)成江宁夏医科大学总医院Oncotarget,2015,6(34):35443-3545715 Enhanced skin permeation ofglabridin using eutecticmixture-based nanoemulsion(基于低共熔物纳米乳提高光甘草定皮肤渗透性研究)刘晨宁夏医科大学总医院Drug Deliv AndTranslRes,2017,7:325-33216 AT-101 enhances gefitinibsensitivity in non-small cell lungcancer with EGFR T790M mutations(AT-101 增加吉非尼对EGFR T790M突变型非小细胞肺癌的治疗敏感性)赵仁宁夏医科大学总医院BMCCANCER,2016,16:49117 Effects of glucose administeredwith lidocaine solution on spinalneurotoxicity in rats(葡萄糖与利多卡因注射液联合使用对大鼠神经毒性的影响)马汉祥宁夏医科大学总医院INTERNATIONALJOURNAL OFCLINICAL ANDEXPERIMENTALMEDICINE,2015,8(11):20638-2064418 MuRF2 regulates PPARγ 1 activity toprotect against diabeticcardiomyopathy and enhance weightgain induced by a high fat diet(MuRF2通过调节PPARγ1活性抑制高脂饮食诱导的糖尿病心肌病和体重增加)何军宁夏医科大学总医院CardiovascDiabetol (心血管糖尿病学)2015,14:9719 A nanocarbon paste electrodemodified with nitrogen-dopedgraphene for square wave anodicstripping voltammetricdetermination of trace lead andcadmium(氮掺杂石墨烯修饰纳米碳糊电极的构建及对痕量铅镉的方波阳极溶出伏安检测)刘新胜宁夏医科大学MicrochimicaActa,183(2)20 Salinomycin exerts anti-angiogenicand anti-tumorigenic activities byinhibiting vascularendothelialgrowth factor receptor2-mediated angiogenesis盐霉素通过VEGFR2/STAT3通路抑制胃癌血管生成和肿瘤生长的作用机制研究陈静宁夏医科大学Oncotarget.2016,7(18):26580-2659221 Sophoridine-loaded PLGAmicrospheres for lung targeting:preparation ,in vitro , and in vivoevaluation(载槐定碱PLGA肺靶向微球的制备及体内外评价)王文苹宁夏医科大学Drug Delivery;922 CD44 receptor targeting andendosomal pH-sensitive dualfunctional hyaluronic acid micellesfor intracellular paclitaxeldelivery(肿瘤胞内pH触发式释药的CD44受体靶向聚合物胶束用于胞内紫杉醇高效递送)刘艳华宁夏医科大学MolecularPharmaceutics23 基于跨理论模型的社区家庭访视护理对老年高血压患者服药遵从行为的影响李胜玲宁夏医科大学中华护理杂志,2016,,51(5):629-63424 Non-enzymatic sensing of uric acidusing a carbon nanotubeionic-liquid paste electrodemodified with poly(β-cyclodextrin)(基于聚β-环糊精修饰碳纳米管-离子液体糊电极的尿酸无酶传感分析)李永红宁夏医科大学MicrochimicaActa,182(11-12)25 Electrochemical determination ofbisphenol A at ordered mesoporouscarbon modified nano-carbon ionicliquid paste electrode(基于介孔碳修饰纳米碳离子液体糊电极的双酚A电化学检测)李永红宁夏医科大学Talanta , 2016 ,148 :362-36926 煤尘致人胚肺成纤维细胞纤维化中胸腺分化抗原-1 DNA 甲基化的变化刘志宏宁夏医科大学环境与职业医学(08)27 职业慢性心理应激与血清热休克蛋白70及肿瘤坏死因子-α表达水平的关联研究宋辉宁夏医科大学中华预防医学杂志,2016,50卷5期:423-42828 Highly sensitive detection ofmercury ion based on T-richDNAmachine using portable glucosemeter 基于便携式血糖仪的富含T碱基DNA机器用于汞离子的高灵敏检测姚惠琴宁夏医科大学Sensors andActuators B29 枸杞多糖联合去细胞异种神经支架修复大鼠坐骨神经缺损的实验研究赵飞宁夏人民医院中华显微外科杂志,2015年12月,第38卷第6期30 宁夏地区带蒂皮瓣远端蒂覆盖软组织缺损延迟愈合的多因素分析赵飞宁夏人民医院中华显微外科杂志,2016年10月,第39卷第5期31 自制C/S-J型胆道自行脱落支架在逆行性胰胆管造影中的临床应用杨勇宁夏人民医院中华肝胆外科杂志,2016年5月,第22卷第5期32 行动研究法(action research)在培养护士科研能力中的应用性研究葛玉荣宁夏人民医院中华护理杂志,2016年1月,第51卷第1期,33 血管损伤对新生内膜增生的影响袁吉祥宁夏人民医院中国循环杂志,2016年12月,第31卷第12期,34 宁夏地区食源性与人源沙门氏菌耐药性与血清型对比研究魏琼宁夏疾病预防控制中心《中国抗生素杂志》2016年9月第41卷第9期35 宁夏致泻性大肠埃希菌的流行特征及耐药现状研究张燕飞宁夏疾病预防控制中心中国抗生素杂志,2015年11月第40卷第11期36 Molecular epidemiology of hepatitisC infections in Ningxia, China:genotype, phylogeny and mutationanalysis 宁夏丙型肝炎分子流行病学研究吴忠兰宁夏疾病预防控制中心VirologyJournal, 2016 ,13 (1) :17237 线粒体通透性转换孔在脂肪乳逆转布比卡因大鼠心肌毒性中的作用杨立斌石嘴山市第一人民医院中华麻醉学杂志,2015年9月第35卷第9期38 农村首发精神分裂症患者睡眠障碍现况及其与社会人口学特征和临床特征之间的关系The Prevalence of SleepDisturbance and TheirSocio-demographic and ClinicalCorrelates in First-episodeIndividuals with Schizophrenia inRural China)马欣荣自治区宁安医院Perspectives inpsychiatriccare39 宁夏地区结核分枝杆菌耐药情况调查王晓林宁夏第四人民医院中华结核和呼吸杂志, 2015 , 38(10) :738-74040 回族患者在斋月发生上消化道出血的临床分析黄李雅宁夏医科大学总医院中华急诊医学杂志,2016年第25卷第8期,1041-1043页41 NF-KBp65对子宫内膜异位症腺上皮细胞OPN,MMP9表达及细胞侵袭性的影响哈春芳宁夏医科大学总医院现代妇产科进展,2016年第25卷第7期,518-522页42 抗N-甲基-D天门冬氨酸受体抗体与系统性红斑狼疮认知功能障碍的相关性研究竺红宁夏医科大学总医院中华风湿病学杂志 2016年第20卷第5期,312-316页43 多参数磁共振成像诊断和鉴别诊断外周带早期前列腺癌和前列腺炎李鹏宁夏医科大学总医院中华医学杂志,2016年第96卷第37期,2973-2977页44 骨代谢等生化指标在不同孕期的代谢特点及相关性研究席向红宁夏医科大学总医院中华医学杂志,2017年第97卷第13期,1015-1018页45 Reosseointegration FollowingRegenerative Therapy ofTissue-Engineered Bone in a CanineModel of ExperimentalPeri-Implantitis(骨结合复合组织工程化骨的再生疗法治疗犬实验性种植体周围炎的研究)孙小娟宁夏医科大学总医院ClinicalImplant Dentistand RelatedResearch 2016年4月18卷0246 病椎固定治疗胸、腰椎结核的疗效分析施建党宁夏医科大学总医院中华骨科杂志,2016,36(11):681-69047 Application of a modifiedthoracoabdominal approach thatavoids cutting open the costalportion of diaphragm duringanterior thoracolumbar spinesurgery避免切开膈肌的改良胸腹联合入路在胸腰段脊柱前路手术中的应用施建党宁夏医科大学总医院Eur Spine J 2016Dec 27 doi:10.1007/s00586-016-4917-2.[Epub ahead ofprint]48 三维斑点追踪技术评价肥厚型心肌病MYBPC3基因截短突变患者左室收缩功能和同步性的早期改变纳丽莎宁夏医科大学总医院中国超声医学杂志,2017年第33卷第3期,228-230页49 急性心肌梗死患者介入治疗术后心理弹性及影响因素分析姚丽宁夏医科大学总医院护理学杂志,2016年第31卷第23期,14-17页50 脊髓κ受体在瑞芬太尼诱发切口痛大鼠术后中枢敏化中的作用:在体脊髓电生理法邓立琴宁夏医科大学总医院中华麻醉学杂志,2016年第36卷第12期,1448-1452页51 Expression of heparin-bindingepidermal growth factor in theendometrium is positivelycorrelated(子宫内膜中HB-EGF可以提高IVF-ET妊娠结局)宋梦玲宁夏医科大学总医院Int J Clin ExpPathol,2016,9(8):8280-828552 血流感染鲍氏不动杆菌耐药性分析周文杰宁夏医科大学总医院中华医院感染学杂志,2016年第26卷第21期,4831-4834页53 火把花根对急性肺损伤大鼠肾组织紧密连接蛋白claudin一5及ZO一1表达的影响马希刚宁夏医科大学总医院中华麻醉学杂志,2016年第36卷第1期,101-104页54 中老年回汉族人群的胰岛素抵抗及β细胞功能与脂代谢等关系的对比研究姜敏宁夏医科大学总医院中国医学科学院学报 2016年第38卷第2期:192-197页55 The polymorphic CAG repeat in exon1of androgen receptor is associatedwith level of HDL-cholesterol andhypertension in Chinese middle-agedto elderly men(雄激素受体基因第一外显子CAG重复多态性与中国中老年男性高密度脂蛋白水平和高血压的相关性)姜敏宁夏医科大学总医院ClinicalEndocrinology(临床内分泌学)2017年4月6日首次在线发表,纸质版在印刷中56 脓毒症患者肠屏障功能损伤及其相关因素王晓红宁夏医科大学总医院中华医学杂志,2016年第96卷第44期,3568-3572页57 宁夏南部山区甲状腺结节和甲状腺功能异常的流行病学调查王婷宁夏医科大学总医院中华健康管理学杂志,2017年第11卷第2期,172-176页58 脑胶质瘤术后IMRT中弥散张量成像示踪视觉通路纤维束应用研究赵超云宁夏医科大学总医院中华放射肿瘤学杂志2016.25(4):315-319页59 Berberine protects the heart fromischemia/reperfusion injury byattenuating endoplasmic reticulumstress via the JAK2/STAT3 signalingpathway(小檗碱通过JAK2/STAT3信号通路抑制内质网应激对大鼠心肌缺血再灌注损伤的保护作用研究)王云宁夏医科大学总医院ActaPharmacologicaSinica (2016)37:1-1460 Involvement of histone methylationin macrophage apoptosis andunstable plaque formation inmethionine-inducedhyperhomocysteinemic ApoE-/- mice(组蛋白甲基化参与蛋氨酸饮食诱发丛广志宁夏医科大学总医院Life Sciences,2017,173:135-144高同型半胱氨酸血症ApoE-/-小鼠斑块不稳定和巨噬细胞凋亡)61 Modulation of FABP4 hypomethylationby DNMT1 and its inverse interactionwith miR-148a/152 in the placenta ofpreeclamptic rats and HTR-8 cells(在妊娠高血压疾病大鼠胎盘组织和滋养细胞中DNMT1与miR-148a/152相互作用调控FABP4 DNA低甲基化)张慧萍宁夏医科大学总医院Placenta. 2016,46:49-62.62 High-throughput sequencing of 16SrDNA amplicons characterizesbacterial composition inbronchoalveolar lavage fluid inpatients withventilator-associated pneumonia(16S rDNA扩增的高通量测序分析呼吸机相关性肺炎患者肺泡灌洗液的细菌组成)杨晓军宁夏医科大学总医院Drug Design,Development andTherapy 2015:94883–489663 Rapamycin Reduced Ischemic BrainDamage in Diabetic Animals IsAssociated with Suppressions ofmTOR and ERK1/2 Signaling(雷帕霉素抑制mTOR及ERK1/2信号通路缓解糖尿病缺血性脑损伤)刘萍宁夏医科大学总医院国际生物科学杂志2016; 12(8),1032-104064 Primary Pulmonary Artery Sarcoma onDual-Time Point FDG PET/CT Imaging(双时相 FDG PET/CT显像在原发性肺动脉肉瘤中的应用)李娟宁夏医科大学总医院ClinicalNuclearMedicine,2016,41(8):456-45865 Comparison of Genetic Variants inCancer-Related Genes betweenChinese Hui and Han Populations(宁夏回汉族人群间肿瘤相关基因遗传变异分布的对比研究)杨文君宁夏医科大学总医院Plos ONE2015,10(12)66 Emerging Role and TherapeuticImplication of Wnt SignalingPathways in Autoimmune Diseases Wnt/β-catenin(Wnt信号通路在自身免疫性疾病中的新作用和治疗意义)石娟宁夏医科大学总医院Journal ofImmunologyResearch,2016,9392132,18pages67 Distinct Roles of Wnt/β-CateninSignaling in the Pathogenesis ofChronic Obstructive PulmonaryDisease and Idiopathic PulmonaryFibrosis(Wnt信号通路在慢性阻塞性肺疾病和特发性肺纤维化发病机制中石娟宁夏医科大学总医院Mediators ofInflammation,2017,3520581,16pages的分子机制)68 High-methionine diets accelerateatherosclerosis by HHcy-mediatedFABP4 gene demethylation pathwayvia DNMT1 in ApoE-/- mice (高蛋氨酸饮食诱导高同型半胱氨酸血症经DNMT1/ FABP4 DNA低甲基化通路促进ApoE-/-鼠动脉粥样硬化发生)张慧萍宁夏医科大学总医院FEBS Letter,2015,589:3998-400969 转化生长因子-β1和白细胞介素-10单核苷酸多态性与复发性口腔溃疡易感性的研究张敬宁夏医科大学总医院华西口腔医学杂志,2016年第34卷第1期,27-31页70 Characteristics oftemporomandibular joint in patientswith temporomandibular jointcomplaint(颞下颌关节紊乱患者的颞下颌关节的特征)郭晓倩宁夏医科大学总医院Int J Clin ExpMed,2015,8(9):16057-1606371 宁夏同心县回族癫痫流行病学调查王旭宁夏医科大学总医院中风与神经疾病杂志,2016年第33卷第1期,27-31页72 多层螺旋CT全肝灌注对肝脏常见肿瘤血流状态的评价陈勇宁夏医科大学总医院中华肿瘤杂志,2015年第37卷第12期,904-908页73 Microemulsions basedonpaeonol-menthol eutectic mixturefor enhanced transdermaldelivery:formulation developmentand in vitro evaluation(基于丹皮酚-薄荷脑低共熔物纳米乳胶用于提高经皮渗透的处方筛选及体内评价)刘晨宁夏医科大学总医院Arif CellsNanomedBiotechnol,1-774 银川市医保试行按病种分值结算效果探析杜永强宁夏医科大学总医院中华医院管理杂志,2016年第32卷第9期,659-662页75 双任务情境下孤独症儿童执行功能研究方建群宁夏医科大学总医院中国儿童保健,2016年第24卷第12期,1245-1248页76 Mi R-126 Suppresses theGlucose-StimulatedProliferation via IRS-2 in INS-1 β陶虹宁夏医科大学PLOS ONE2016,11(2):e0149954Cells(miR-126通过IRS-2抑制糖刺激引起的INS-1 β细胞增殖)77 Hyperhomocysteinemia in ApoE-/-Mice leads to overexpression ofenhancer of zeste homolog 2 viamiR-92a regulation (miR-92a靶向调控EZH2介导同型半胱氨酸致动脉粥样硬化的机制研究)杨晓玲宁夏医科大学Plos one78 Oxymatrine attenuatedhypoxic-ischemic brain damage inneonatal rats via improvingantioxidant enzyme activities andinhibiting cell death(氧化苦参碱通过增加抗氧化酶活性和抑制细胞凋亡减轻新生大鼠缺血缺氧性脑损伤)余建强宁夏医科大学NeurochemistryInternational;8979 The anticonvulsant andneuroprotective effects ofoxysophocarpine onpilocarpine-induced convulsions inadult male mice氧化槐果碱对匹罗卡品诱发小鼠癫痫的抗癫痫及神经保护作用汪静宁夏医科大学Cell MolNeurobiol;3780 Protective effect ofgentiopicroside against dextransodium sulfate induced colitis inmice(龙胆苦胆对葡聚糖硫酸钠诱导的小鼠溃疡性结肠炎的保护作用)赵玉萍宁夏医科大学InternationalImmunopharmacology;3981 Comparative studies on theproperties of glycyrrhetinicacid-loaded PLGA microparticlesprepared by emulsion and templatemethods(采用乳液法和凝胶模板法制备载甘草次酸PLGA微球特性的比较研究)王文苹宁夏医科大学InternationalJournal ofPharmaceutics;49682 Protective effect ofsophoraflavanone G onstreptozotocin (STZ)-inducedinflammation in diabetic rats(苦豆子中Sophoraflavanone G对链脲佐菌素(STZ)诱导的糖尿病大鼠降糖及并发症的保护作用)王汉卿宁夏医科大学Biomedicine &Pharmacotherapy;8983 Neuroprotective effect andmechanism of MU-Xiang-You-Fang on赵启鹏宁夏医科大学Journal ofEthnopharmacolocerebral ischemia-reperfusioninjury in rats(回药木香油方抗脑缺血再灌注损伤的作用及机制研究)gy,19284 Lycium barbarum polysaccharidesattenuatesN-methy-N-nitrosourea-inducedphotoreceptor cell apotosis in ratsthrough regulation of poly(ADP-ribose ) polymerase andcaspase expression (枸杞多糖通过调节PARP和Caspase的表达来改善MNU诱导的大鼠感光细胞凋亡)朱亚飞宁夏医科大学Journal ofEthnopharmacology,19185 Anti-Proliferative Effect ofTriterpenoidal Glycosides from theRoots of Anemone vitifolia throughaPro-Apoptotic Way白长财宁夏医科大学molecules,2017,22(4):64286 Construction and characterizationof osteogenic and vascularendothelial cell sheets from ratadipose-derived mesenchymal stemcells(SD大鼠脂肪间充质干细胞向成骨细胞、血管内皮细胞诱导分化构建双细胞膜片的研究)张华林宁夏医科大学Tissue andCell,2016年,第48卷,第5期,488-49587 Bilayered PLGA/Wool KeratinComposite Membranes SupportPeriodontal Regeneration in BeagleDogs(双层PLGA/羊毛角蛋白复合膜用于Beagle犬牙周组织再生的研究)张华林宁夏医科大学ACSBiomaterialsScience &Engineering,2016年,第2卷第12期,2162-217588 银川市儿童监护人脊灰疫苗知晓率及其影响因素分析杨婵宁夏医科大学中华疾病控制杂志,2016,20(8):805-80889 基于三维质量结构模式的社区产后家庭访视护理质量评价体系构建研究刘国莲宁夏医科大学中国全科医学,2016,19(19):2246-225190 Inhibition of Connective TissueGrowth Factor AttenuatesParaquat-Induced Lung Fibrosis in aHuman MRC-5 Cell Line(CTGF基因沉默对百草枯致MRC-5细胞纤维化的影响)黄敏宁夏医科大学EnvironmentalToxicology,2016 , 31(11) :162091 百草枯诱导人神经胚胎干细胞凋亡中黄敏宁夏医科大中华劳动卫生职microRNA 表达特征及功能学业病 , 2017 , 35(1) :19-2492 百草枯对人胚胎神经干细胞增殖过程中的PKC/ERK信号通路分子表达的影响蔡倩宁夏医科大学环境与职业医学 , 2017 , 34(1) :42-4893 Lycium barbarumL.Polysaccharide(LBP) reducesGlucose Uptake via Down-Regulationof SGLT-1 In Caco2 Cell (枸杞多糖通过下调SGLT-1受体抑制了Caco2细胞对葡萄糖的摄取)蔡慧珍宁夏医科大学Molecules,22卷2期94 不同严重程度社区获得性肺炎患者凝血与纤溶指标的研究白淑荣宁夏人民医院中华医院感染学杂志,2016年10月,第26卷第19期95 宁夏地区血液透析患者生存状态的质性研究葛玉荣宁夏人民医院中国实用护理杂志,2016年5月,第33卷第15期96 冥想训练对瓣膜性心脏病老年患者术后认知功能障碍的改善作用陈玉梅宁夏人民医院中华行为医学与脑科学杂志,2016年12月,第25卷第12期,97 Reduced inflammatory cellrecruitment and tissue damage inspinal cord injury by acellularspinal cord scaffoldseeded with mesenchymal stem cells9(脊髓脱细胞支架复合间充质干细胞减轻脊髓损伤后炎性细胞浸润和组织损伤)汪玉海宁夏人民医院EXPERIMENTALAND THERAPEUTICMEDICINE,2017年1月,第13卷第1期,98 (Selective Hemihepatic VascularOcclusion Versus Pringle Maneuverin Hepatectomy for Primary LiverCancer)选择性半肝血流阻断与Pringle手法在原发性肝癌切除术中的应用研究李明皓宁夏人民医院WORLD J SURGONCOL (世界外科肿瘤杂志),2017,(9):2399 2012-2016年鲍氏不动杆菌的临床分布与耐药性分析李怡宁夏人民医院中华医院感染学杂志,2017年4月,第27卷第8期100 Tongue acupuncture to treatpost-stroke dysphagia. (舌针治疗脑卒中后吞咽困难)高焕民宁夏人民医院Int J Clin ExpMed 2015;8(8)101 沉默Cdk5或Cdk5激活剂p35的基因表达与足细胞凋亡的相关性研究;KnockdownofExpressionofCdk5orp35(a Cdk5Activator)ResultsinPodocyteApoptosis郑亚莉宁夏人民医院Plos one2016,Jul,15;11(8):1371-84.102 宁夏地区医疗照射频率水平调查分析孙彦玲宁夏疾病预防控制中心中华放射医学与防护杂志 ,2016 , 36(10) :757-760103 2013-2014年手足口病患者柯萨奇病毒A10型宁夏分离株VP1区基因特征分析马江涛宁夏疾病预防控制中心中华预防医学杂志 , 2015 , 49(12) :1056-1060104 某企业工作场所锰水平和工人职业健康体检结果分析孙伟宁夏疾病预防控制中心中华劳动卫生职业病杂志105 综合ICU患者肺部感染肠杆菌科病原菌耐药性及防控措施宇文慧宁夏第五人民医院中国消毒学杂志2015年12期106 成人与儿童下呼吸道感染患者的病原菌分布及耐药性分析李建华宁夏第五人民医院中华医院感染学杂志2015年第20期107 家庭访视护理对社区老年糖尿病患者自我效能的影响游书秋银川市第一人民医院实用医学杂志2016年6月第32卷12期108 品管圈护理模式应用偏瘫患者实施健康教育的效果分析张惠霞石嘴山第一人民医院医学信息, 2015(z2) :161-161109 分析舒适护理在门诊无痛人流术中的应用效果张惠霞石嘴山第一人民医院现代养生月刊 ,2016(7) :254-254110 宁夏地区公共场所控烟现状及消费者吸烟行为和控烟态度调查刘凯宁夏卫生和计划生育监督局《中华疾病控制杂志》2015年19卷第9期111 急性肠黏膜损伤导致躯体穴区敏化的电生理学特征许建峰宁夏医科大学总医院针刺研究,2015年第40卷第3期,180-185页112 KIF14过表达与胃肠癌患者预后的关系陈萍宁夏医科大学总医院中国全科医学,2016年第19卷第8期,931-940页113 乳腺癌组织中耐药相关基因MDR1、BCRP和LRP表达的研究陈萍宁夏医科大学总医院宁夏医科大学学报,2016年第38卷第1期,13-17页114 Storage time of intraoperativetransfused allogeneic red blood顾继伟宁夏医科大学总医院PLOS ONE,2017cells is not associated with new-onset postoperative atrial fibrillation in cardiac surgery (心脏外科术后新发房颤与术中输注同种异体红细胞的存储时间的相关性研究)115 Impact of chrysosplenetin on thepharmacokinetics and anti-malarialefficacy of artemisinin againstPlasmodium berghei as well as invitro CYP450 enzymatic activitiesin rat liver microsome(猫眼草黄素对青蒿素药代动力学、抗伯氏疟原虫活性以及对大鼠肝微粒体CYP450体外活性影响研究)魏世杰宁夏医科大学总医院MalariaJournal,Volume14:432,November 2015,页码:1-13116 Regioselective synthesis,stability and release behaviors ofthe secondary 5-fluorouracil aceticacid/b-cyclodextrin conjugate forcolon delivery(5-氟尿嘧啶乙酸-β-环糊精共轭物的选择性合成、稳定性及其结肠释药性能的研究)魏世杰宁夏医科大学总医院Journal ofInclusionPhenomena andMacrocyclicChemistry.Volume 84, December2015,页码:43-47117 Matrine amelioratesisoproterenol-induced acutemyocardial ischemia throughregulation of growth factors andRhoA/ROCK1 pathway(苦参碱调控细胞因子与RhoA/ROCK信号通路抑制异丙肾上腺素致大鼠心肌肥厚作用及机制研究)党宏万宁夏医科大学总医院InternationalJournal ofClinical andExperimentMedicine 2016年9卷第11期,页码:21961-21969118 经后入路手术治疗先天性颅颈交界区不稳定性疾病刘诤宁夏医科大学总医院中华外科杂志,2016年第54卷第5期,391-393页119 基于PDCA的自我管理干预模式对老年经皮冠状动脉介入治疗术术后患者健康行为及用药依从性的影响姚丽宁夏医科大学总医院中国实用护理杂志,2016年第32卷第25期,1931-1937页120 早期预警评分联合标准化沟通模式在食管癌患者围手术期管理中的应用张曦宁夏医科大学总医院中国实用护理杂志,2017年第33卷第12期,928-932页121 The role of p38MAPK activation inspinal dorsal horn inremifentanil-induced postoperative邓立琴宁夏医科大学总医院NeurologicalResearch,2016,38(17):929hyperalgesia in rats(脊髓背角p38MAPK活化在瑞芬太尼诱发大鼠术后痛觉过敏中的作用)-936122 血清及卵泡液中肝素结合性表皮生长因子水平与多囊卵巢综合征关系的研究宋梦玲宁夏医科大学总医院实用妇产科杂志,2015年第31卷第7期,508-511页123 温针灸对兔膝骨性关节炎关节软骨尿激酶型纤溶酶原活物系统的影响武永利宁夏医科大学总医院中华中医药杂志,2017年第32卷第2期,735-738页124 调强放疗计划中射野角度归零与实际角度的剂量验证比较孔伟宁夏医科大学总医院中国医学物理学杂志,第32卷第6期,892页-896页125 25羟维生素D在原发性胆汁性肝硬化中的诊断价值苏荣宁夏医科大学总医院中国免疫学杂志,2016年第32卷第12期,1830-1836页126 男性儿童与成人系统性红斑狼疮的临床分析苏荣宁夏医科大学总医院中国全科医学,2016年第19卷第9期,1095-1098页127 Prevalence of Genes of OXA-23Carbapenemase and AdeABC EffluxPump Associated with MultidrugResistance of Acinetobacterbaumannii Isolates in the ICU of aComprehensive Hospital ofNorthwestern China(中国西北地区综合医院重症监护病房分离的耐碳青霉烯类鲍氏不动杆菌AdeABC药物外排泵基因及耐碳青霉烯酶基因OXA-23的研究)贾伟宁夏医科大学总医院InternationalJournal ofEnvironmentalResearch andPublicHealth,2015,12,10079-10092128 烧伤科患者肠杆菌科细菌耐药性与基因型分布师志云宁夏医科大学总医院中华医院感染学杂志,2016,26(12):2660-2663129 Clinical and genomic responses toultra-short course chemotherapy inspinal tuberculosis(超短程化疗治疗脊柱结核的临床疗效和基因表达谱的变化的研究)牛宁奎宁夏医科大学总医院EXPERIMENTALAND THERAPEUTICMEDICINE,2017,13:1681-1688130 Anti-tumor Necrosis Factor TherapyIncreased Spineand Femoral NeckBone Mineral Density of Patients李海波宁夏医科大学总医院TheJournalofRheumatology2015,(8):with Active Ankylosing Spondylitiswith Low BoneMineral Density(肿瘤坏死因子-α拮抗剂治疗增加了伴有低骨密度的活动性强直性脊柱炎患者脊柱和股骨颈的骨密度)1413-1417131 磁敏感加权成像在肺癌脑转移瘤诊断中的应用价值王晓东宁夏医科大学总医院中国医学计算机成像杂志2016年第22卷第3期,198-203页132 基于第二版前列腺影像报告和数据系统的多参数MRI诊断外周带前列腺癌的价值陈志强宁夏医科大学总医院中国医学影像技术,2016年第32卷第9期,1414-1418页133 Water-soluble coenzyme Q10 reducesrotenone-induced mitochondrialfission(水溶性辅酶Q10降低鱼藤酮诱导的线粒体分裂)李海宁宁夏医科大学总医院Neurochemicalresearch.2017.42(2):1096-1103134 双源CT灌注缺损评分评价急性肺栓塞严重程度的临床应用朱力宁夏医科大学总医院中国医学影像技术,2015年第31卷第10期,95-99页135 The Expression of NOX4 in SmoothMuscles of Small Airway Correlateswith the Disease Severity of COPD(COPD小气道平滑肌细胞NOX4表达与其疾病严重程度的相关性研究)刘鲜艳宁夏医科大学总医院BioMed ResearchInternational2016.08.30136 血清降钙素原与烧伤脓毒症患者血细菌培养类别的关系沈江涌宁夏医科大学总医院中华烧伤杂志,2015年第31卷第4期,304-305页137 集束化口腔护理方案在防治鼻咽癌放化疗患者口腔黏膜炎中的应用夏莉娟宁夏医科大学总医院中国实用护理杂志,2017年第33卷第10期,758-763页138 ABCB1基因多态性对乳腺癌紫杉类药物化疗疗效的影响刘新兰宁夏医科大学总医院西安交通大学学报(医学版),2016年第37卷第3期,330-334页139 骨桥蛋白在胸椎黄韧带骨化中的定位表达及意义袁海峰宁夏医科大学总医院宁夏医学杂志,2015年第37卷第11期,964-966页140 SPECT/CT显像对布鲁氏杆菌脊柱炎的诊断价值李艳梅宁夏医科大学总医院中华核医学与分子影像杂志,2016年第36卷第6期,516-520页141 鼻用糖皮质激素结合鼻腔冲洗治疗变应性鼻炎的疗效观察高小平宁夏医科大学总医院临床耳鼻咽喉头颈外科杂志,2016年第30卷第9期,702-706页142 树脂粘接技术在保存牙体修复中的应用研究朱琳虹宁夏牙科大学总医院宁夏医学杂志,2015年第37卷第7期,598-600页143 宁夏农村惊厥性癫痫患者死因分析晁丽娜宁夏医科大学总医院中国神经免疫学和神经病学杂志,2017年第24卷第2期,118-122页144 对比分析容积CT剂量指数与体型特异性的剂量评估在估算腹部CT扫描辐射剂量中的差异陈勇宁夏医科大学总医院中华放射医学与防护杂志,2016年第36卷第1期,74-77页145 围手术期小剂量地塞米松对甲状腺肿瘤切除术后咽喉痛的预防作用张慧文宁夏医科大学总医院上海交通大学学报,2016年第36卷第6期,870-874页146 经颅多普勒超声对颈动脉内膜剥脱术围手术期脑血流的评估与应用陈景云宁夏医科大学总医院宁夏医科大学学报,2016年第38卷第10期,1130-1134页147 探讨高敏肌钙蛋白I联合常规心肌标志物对微小心肌损伤的诊断价值张成磊宁夏医科大学总医院中国卫生检验杂志,2016年第26卷第21期,3143-3146页148 医保按病种分值付费对医院管理的影响及对策杜永强宁夏医科大学总医院中华医院管理杂志,2015年第31卷第9期,668-671页149 贝前列素钠对肺源性心脏病肺动脉高压患者新功能及血气指标的影响马宣宁夏医科大学总医院宁夏医学杂志,2016年第38卷第11期,1016-1018页150 CTEPH患者血浆NT-proBNP与右心漂浮导管测定血流动力学的关系马宣宁夏医科大学总医院临床肺科杂志,2017年第22卷第5期,897-899页151 Muscle ring finger-3 protectsagainst diabetic cardiomyopathyinduced by a high fat diet(MuRF3抑制高脂饮食诱导的糖尿病心肌病)何军宁夏医科大学总医院EndocrineDisorders (内分泌疾病)2015,15:36。

姜黄素在肺部疾病中的研究进展张金兰;李朝霞;黄支隆【摘要】目的姜黄素(curcumin)是姜黄发挥药理作用的主要活性成分,它从姜科姜黄属植物姜黄的根茎中提取而来,由于其色泽及味道独特,长期以来被人们当作一种食品添加剂,如咖喱粉.近年来大量研究已证实姜黄素具有抗炎、抗氧化、抗血管生成、抗纤维化、减少气道粘液分泌以及防癌抗癌等药理作用,且毒副作用小.姜黄素越来越受关注,但系统阐述姜黄素在肺部疾病中应用的文章并不多,本文将综述姜黄素在肺部疾病中的应用,为进一步开发姜黄素在肺部疾病中的应用提供理论依据.【期刊名称】《贵州医药》【年(卷),期】2019(043)001【总页数】4页(P42-45)【关键词】姜黄素;肺部疾病;研究进展【作者】张金兰;李朝霞;黄支隆【作者单位】贵州医科大学,贵州贵阳 550004;贵州医科大学,贵州贵阳 550004;贵州医科大学,贵州贵阳 550004【正文语种】中文【中图分类】R5631 姜黄素的生物学特性1.1 姜黄素的结构姜黄素的分子式为C21H20O6,其化学结构如图1所示,主链为不饱和酯及芳香族基团,故而在水中的溶解度很低,是植物界少有的具有二酮色素的二酮类化合物,1870年首次从姜黄中分离出来,1910年被发现其双阿魏酰甲烷的化学结构。
姜黄素为橙黄色结晶粉末,味稍苦,是一种相对分子质量较低的脂溶性多酚类化合物,不溶于水和乙醚,溶于乙醇、丙二醇,易溶于冰醋酸和碱溶液。
图1 姜黄素的化学结构1.2 姜黄素的生物学作用现有的研究已发现姜黄素具有抗炎、抗氧化、抗病毒、抗肿瘤、调脂、抗凝、抗纤维化、抗动脉粥样硬化等广泛的药理活性,且毒性低、不良反应小。
近年来,姜黄素得到了众多研究者的青睐,越来越多的研究表明,姜黄素具有广泛的治疗特性,在多种疾病中具有良好的临床应用前景。
2 姜黄素在肺部疾病中的作用2.1 抗炎炎症是很多肺部疾病的诱因,也是影响肺部疾病预后不良的独立危险因素,姜黄素能作用于炎症反应相关的许多分子靶标来调控炎症反应。

细胞凋亡一词来源于希腊语, 原指树叶或花的自然凋落。
由英、美三位科学家在1972年首次提出,于2002年获得诺贝尔医学奖。
细胞凋亡是指机体在一定生理、病理条件下为维持内环境稳定, 通过基因控制而使细胞主动有序的死亡, 表现一系列形态和生化方面的特征。
细胞凋亡是细胞正常的死亡,它涉及一系列基因的激活、表达以及调控等作用,不造成炎症和自体的损伤, 是细胞为了更好地适应生存环境而主动争取的死亡过程。
因这一死亡过程严格受到程序的控制, 又称细胞程序性死亡(Progammed cell death,PCD)【1】。
一.环境因子与PCD1 病原体与植物PCD植物受病原物(真菌、细菌和病毒)感染时往往会发生过敏性细胞死亡,即过敏反应(hypersensitive response,HR):一方面死亡细胞可以将入侵寄主体内的病原病原微生物限制在感染点周围,阻止其进一步扩展而实现局部抗病性;另一方面也可进一步激活植物抗病防卫的相关信号通路,使植物获得对此后入侵的多种类型病原物的系统抗病性[2]。
现有研究结果表明,植物HR 中的细胞死亡往往表现出PCD 的典型特征。
例如,Wakabayashi 等[3]发现,生菜感染细菌后,细胞线粒体会增大,出现类似动物PCD 的症状,细胞质内空泡化增加,使细胞丧失功能。
Olszak等[4]用细菌侵染拟南芥突变体acdll 后发现,拟南芥AtFMO基因蛋白的表达既可以激活细胞的防御系统,又可以促进细胞发生PCD 现象,但并不是细胞发生PCD 现象所必需的。
2 低氧与植物PCD由于淹水或水培中微生物和植物根的呼吸,降低了植物根际氧的浓度而出现了低氧或无氧状态,导致根部细胞发生PCD现象。
He 等[5]研究表明,Ca2+和蛋白质磷酸化与植物通气组织的形成有关:在正常供氧条件下Ca2+通道抑制剂和蛋白质磷酸酯酶抑制剂都能促进玉米根中细胞的死亡,而在低氧的根中Ca2+螯合物EGTA 和蛋白激酶抑制剂都能防止细胞死亡。

台盼蓝染色细胞存活率检测试剂盒产品编号 产品名称包装 C0011台盼蓝染色细胞存活率检测试剂盒100次产品简介:碧云天生产的台盼蓝染色细胞存活率检测试剂盒(Trypan Blue Staining Cell Viability Assay Kit ),是利用正常的健康细胞能够排斥台盼蓝,而丧失细胞膜完整性的细胞可以被台盼蓝染色研制而成。
严格来说,台盼蓝染色检测的是细胞膜的完整性,通常认为细胞膜丧失完整性,即可认为细胞已经死亡。
台盼蓝染色后,通过显微镜下直接计数或显微镜下拍照后计数,就可以对细胞存活率进行比较精确的定量。
台盼蓝染色后的HeLa 细胞请参考图1。
图1. 细胞台盼蓝染色效果图。
胰酶消化下来的HeLa 细胞,1000g 离心1分钟,弃上清,用适量细胞重悬液重悬后,用等体积台盼蓝染色液(2X)染色。
箭头所示为台盼蓝染色的死亡细胞。
注:本HeLa 细胞经过一定的细胞坏死诱导处理。
台盼蓝染色只需3-5分钟即可完成,并且操作非常简单。
本试剂盒足够检测100个细胞样品。
包装清单:产品编号 产品名称 包装 C0011-1 台盼蓝染色液(2X) 10ml C0011-2细胞重悬液 100ml —说明书1份保存条件:4ºC 保存,一年有效。
注意事项:本试剂盒提供的两种溶液都是无菌的,使用时最好在超净台内进行,避免细菌污染。
本产品长时间存放可能会出现少量颗粒状沉淀,可在37ºC 水浴约10分钟以充分溶解沉淀。
沉淀完全溶解后即可正常使用。
台盼蓝对人体有毒,操作时请特别小心,并注意有效防护以避免直接接触人体或吸入体内。
本产品仅限于专业人员的科学研究用,不得用于临床诊断或治疗,不得用于食品或药品,不得存放于普通住宅内。
为了您的安全和健康,请穿实验服并戴一次性手套操作。
使用说明:1. 收集细胞:对于贴壁细胞先用胰酶和/或EDTA 消化下细胞。
对于悬浮细胞,则可以直接收集细胞。
把收集的细胞在1000-2000g 离心1分钟,弃上清,用1毫升或根据细胞的量用适当细胞重悬液重新悬起细胞。
C-band LNB for earth station C波段卫星地⾯站⾼频头 C-Brite C-布莱特超级铁素体不锈钢 C-clamp set C型夹钳 C-frame hydraulic press C型机架液压机 C-frame hydraulic straightening and mounting press C型机架校直压装液压机 C-reactive protein and C-reactive protein test plate C反应蛋⽩及其检测板 C-scanner C型超声波扫描器 C9 aromatic solvent 碳九芳烃溶剂 CA alloy CA铜铝镍硅耐蚀合⾦ Cab-O-sil 硅⽯粉 cab-over-engine truck 驾驶室在发动机上部的货车 cabal glass 钙硼铝玻璃 Caball 卡巴尔牌汽车 Caballero 骑⼠牌汽车 cabana cloth 运动服织物 cabbage ball 椰菜 cabbage lettuce 结球莴苣 cabbage mustard 芥蓝 cabbage oil 菜油 cabbage palm 棕榈属叶纤维 cabbage 圆⽩菜 cabbaging press 包装压榨机 cabeca 卡⽐萨丝 Cabernet 解百衲葡萄酒 cabesa 卡⽐沙⽺⽑ cabicidin 杀真菌素 cabin cableway 吊舱索道 cabin hook 窗钩 cabin ration feeding machine 仓式定量给料机 cabin ropeway 吊舱索道 cabinet drier 烘箱 cabinet file 半圆锉 cabinet for television set 电视机柜 cabinet loudspeaker 箱式扬声器 cabinet mirror 柜镜 cabinet oven 柜式烤炉 cabinet panel 配电盘 cabinet plane ⽊⼯刨 cabinet rack 机箱架 cabinet scraper ⽊⼯刮⼑ cabinet speaker 箱式扬声器 cabinet ventilator 箱式通⽓器 cabinet water-proof switch 柜式防⽔开关 cabinet with dragon design 云龙神台 cabinet with mirror door 带镜⽴柜 cabinet 机壳 cabinet-type hatcher 柜式孵卵器 cabinet-type switch 柜式开关 cabinet-working machinery 细⽊⼯机械 Cabisan 酚铵杀菌剂 cabistan 卡⽐斯顿地毯 cable excavator 电缆管道挖沟机 cable armour cutter 电缆钢带剥切⼑ cable bearer 电缆⽀架 cable boring tool 电缆表⾯穿孔器 cable box 电缆箱 cable braiding machine 编绳索机 cable brake 缆式制动器 cable branch box 电缆分线箱 cable calibration amplifier 电缆校正放⼤器 cable car 缆车 cable change-over box 电缆转换箱 cable channel 电缆管道 cable chock 导缆器 cable clamp collar 电缆连接卡头 cable clip 电缆夹 cable compass 电缆罗盘 cable compound 电缆绝缘胶 cable conntecting plug 电缆联接插头 cable conveyer 缆索运输机 cable cord tyre 帘布轮胎 cable cord 阔条灯芯绒 cable coupling box 电缆接线盒 cable cover lead cutter 电缆破铅⽪⼑ cable covering machine 电缆包编机 cable crane 缆索式起重机 cable cutter 电缆剪 cable deviation equalizer 电缆偏差均衡器 cable diameter gauge 缆径测量仪 cable drag scraper 缆索拖铲 cable drilling outfit 索钻设备 cable drum jack 电缆卷盘架 cable drum trailer 电缆卷筒拖车 cable drum 电缆卷筒 cable duct tube 电缆导管 cable fault detector 电缆故障检测器 cable fault finder 电缆故障测定仪 cable fault locator 电缆故障定位仪 cable fault search set 电缆事故检查器 cable flaw detector 电缆探伤仪 cable foot engine ⾜踏缆车 cable for ship 船⽤电缆 cable gear 敷电缆机 cable grease 电缆脂 cable grip 电缆钳 cable guide 电缆引导管 cable gum 电缆胶 cable hanger 电缆吊架 cable head 电缆头 cable hoist 缆索绞车 cable hoisting pulley 钢索起重滑车 cable in cotton 棉纱包电线 cable in lead 铅包电缆 cable in plastics 塑料电线 cable in rubber 橡⽪电线 cable input plug 电缆输⼊插头 cable insulating paper 绝缘电缆纸 cable iron 钢绳铁 cable isolating paper 电缆绝缘纸 cable isolator 电缆隔离器 cable jack 电缆盘千⽄顶 cable joint 电缆接头 cable knife 电缆⼑ cable lac 电缆胶漆 cable lay wire rope 缆式钢丝绳 cable layer 电缆敷设船 cable laying ship 电缆敷设船 cable laying truck 电缆敷设车 cable length switch 电缆长度转换开关 cable lift 钢索起重机 cable lifter 起锚机 cable line tester 电缆路径测试仪 cable mark detector 电缆记号探测器 cable marking paper 电缆标记⽤纸 cable mattings 亚⿇厚帆布 cable monitor 电缆监控器 cable net ⼤眼窗纱 cable optical fiber 成缆光纤 cable paper 电缆纸 cable picking and laying machine 电缆收放机 cable rack 电缆架 cable reel 电缆盘 cable relay 电缆继电器 cable release 快门线 cable rig 缆索凿井机 cable ring 电缆圈 cable semicontinuous casting machine 钢丝绳半连续铸造机 cable sender 电缆传送机 cable sheath 电缆包⽪ cable ship 铺设电缆船 cable shoe 电缆终端套管 cable silk 饰边粗丝线 cable sleeve 电缆套管 cable stand 电缆架 cable tank 电缆槽 cable tensiometer 电缆拉⼒计 cable tension indicator 钢绳张⼒指⽰器 cable terminal rack 电缆转接架 cable terminating equipment 电缆终端设备 cable tester 电缆测试器 cable testing set 电缆测试器 cable thread 三股缝纫线 cable tool drill 钢绳冲击式钻机 cable trailer 电缆拖车 cable tram 缆道电车 cable transmitter ⽔线电报发报机 cable transporting truck 电缆运输车 cable tray 电缆槽 cable trunk 电缆管道 cable TV control system 闭路电视控制系统 cable vessel 电缆船 cable winch 钢索绞车 cable wire 钢丝绳 cable working vehicle 电缆⼯程车 cable wrapping paper 电缆包装纸 cable yarn 电缆纱线 cable 电缆 cable-actuated excavator 缆控挖⼟机 cable-driven car 钢丝绳牵引车 cable-driven ingot buggy 钢丝绳牵引的送锭车 cable-dump truck 钢索⾃卸卡车 cable-former 成缆机 cable-laid rope 多股钢丝绳 cable-operated mucker 抓⽃式装岩机 cable-spinning equipment 钢缆绞车 cable-suspended feeder 钢索悬吊式给料机 cable-towed machine 钢索牵引机 cabled netting twine 结绳 cabled yarn 缆线 cablet 细缆 cableway bucket 索道吊⽃ cableway carriage 架空索道车 cableway tractor 索道牵引机 cabline patchouli ⼴藿⾹ cabling machine 搓绳机 cabo negro 棕榈属叶纤维 cabochon 凸⾯宝⽯ cabots 卡博茨平布 cabretta leather ⽺⽪⾰ cabriolet 活顶⼩轿车 Cabstar 卡星牌汽车 cabtyre cable 橡⽪绝缘软电缆 cabtyre cord 橡⽪绝缘软线 cabtyre flexible wire 橡套软线 cabtyre wire 厚橡胶绝缘软性线 cabulla fibre 中美洲西沙尔⿇ cabuya 卡布耶叶纤维 cacao bean cleaner 可可⾖清选机 cacao bean separator 可可⾖分离器 cacao bean sorting machine 可可⾖分级机 cacao bean 可可⾖ cacao oil 可可油 cacao pigment 可可壳⾊ cacharado 亚⿇平布 cache buffer memory 超⾼速缓冲存储器 cache card ⾼速缓冲存储卡 cache 超⾼速缓冲存储器 cachemire de soie ⾼级塔夫绸 cachemirette 仿开⼠⽶斜纹绒 cachenez 围⼱ cachmire shawl ⽺绒披⼱ cack ⼩孩平底鞋 cacoclasite 钙铝黄长⽯ cacodylic acid 卡可基酸 cacotheline 硝基马钱⼦碱 cactinomycin 放线菌素C cactus needle 仙⼈掌式唱针 cactus 仙⼈掌 CAD program of microwave circuit 微波电路计算机辅助设计软件 Cad-Trete 福钙混剂 caddis 卡迪斯斜纹呢 caddow 花式⽑圈布 Caddy 卡迪牌汽车 caddy 茶叶罐 Caddy 卡迪牌汽车 Caddy 氯化镉 cade oil 杜松油 cadet blue cloth 军校灰蓝⾊布 cadet cloth 军校制服呢 cadet 短指⼿套 cadexomer iodine 卡地姆碘 cadger ⼩型注油器 Cadillac 卡迪拉克牌汽车 cadillo 肖梵夫花韧⽪纤维 cadillon 卡迪翁斜纹呢 cadion 镉试剂 cadmium acetate ⼄酸镉 cadmium amalgam 镉汞合⾦ cadmium bearing alloy 镉轴承合⾦ cadmium borate 硼酸镉 cadmium bromide 溴化镉 cadmium carbonate 碳酸镉 cadmium cell 镉电池 cadmium chloride 氯化镉 cadmium copper 镉铜 cadmium filter 镉过滤器 cadmium fluoride photon spectrometer 氟化镉光⼦谱仪 cadmium fluoride 氟化镉 cadmium gold 镉⾦装饰合⾦ cadmium ingot 镉锭 cadmium iodide 碘化镉 cadmium laser 镉激光器 cadmium magnesium alloy 镉镁合⾦ cadmium mercury 镉汞 cadmium metal ⾦属镉 cadmium nickel alloy 镉镍合⾦ cadmium niobate 铌酸镉 cadmium nitrate 硝酸镉 cadmium orange 镉桔黄 cadmium oxalate 草酸镉 cadmium oxide 氧化镉 cadmium photocell 镉光电池 cadmium red 镉红 cadmium salt 镉盐 cadmium selenide 硒化镉 cadmium silicate 硅酸镉 cadmium silver 镉银合⾦ cadmium solder 镉焊料 cadmium standard cell 镉标准电池 cadmium sulfide cell 硫化镉电池 cadmium sulfide detector 硫化镉探测器 cadmium sulfide 硫化镉 cadmium telluride 碲化镉 cadmium test voltmeter 镉棒测试专⽤伏特计 cadmium titanate 钛酸镉 cadmium tungstate 钨酸镉 cadmium vapor arc lamp 镉汽弧灯 cadmium yellow 镉黄 cadmium zirconate 锆酸镉 cadmium 镉 cadmium-copper wire 镉铜线 cadmium-covered detector 敷镉检测器 cadmium-doped germanium detector 锗掺镉探测器 cadmium-mercury excimer laser 镉-汞受激准分⼦激光器 cadmium-nickel storage battery 镉镍蓄电池 cadmium-nickel storage cell 镉镍蓄电池 cadmium-plated steel 镀镉钢 cadmium-selenide detector 硒-镉探测器 cadmium-selenium red 隔硒红 cadmium-telluride gamma-ray spectrometer 碲化镉γ射线谱仪 cadmium-telluride high temperature infrared fire detector 碲化镉⾼温红外⽕情探测器 cadmium-zinc eutectic alloy 镉锌共晶合⾦ Cadola 卡多拉牌⼿表 Cadon 卡唐多叶形耐纶66长丝 caen 卡昂哔叽 caesalpinia ⽯莲⼦ caesium atomic clock 铯原⼦钟 caesium bicarbonate 碳酸氢铯 caesium bromide 溴化铯 caesium carbonate 碳酸铯 caesium chloribe 氯化铯 caesium chromate 铬酸铯 caesium discharge tube 铯放电管 caesium fluoride 氟化铯 caesium hydroxide 氢氧化铯 caesium iodide 碘化铯 caesium laser 铯激光器 caesium nitrate 硝酸铯 caesium photocell 铯光电管 caesium sulfate 硫酸铯 caesium vapour laser 铯蒸⽓激光器 caesium vapour rectifier 铯蒸发整流管 caesium 铯 caesium-oxygen cell 充⽓铯光电管 cafedrine 咖⿇黄碱 cafergot tablet 加⾮葛⽚ caffard 卡法德呢 caffeine citrate 柠檬酸咖啡因 caffeine sodium benzoate 安钠咖 caffeine 咖啡因 Cafina 克飞纳牌⼿表 Cafor 卡福牌⼿表 caftan 卡夫坦长⾐ café curtain 褶裥窗帘 cage antenna 笼形天线 cage disintegrator 笼式粉碎机 cage goniometer 笼形测向器 cage hoist 罐笼提升机 cage motor ⿏笼式电动机 cage positioned ball valve 笼式定位球调节阀 cage rotor ⿏笼式转⼦ cage type airless shot blasting machine ⿏笼式抛丸清理机 cage type induction motor ⿏笼式感应电动机 cage type multiplier 笼形倍增器 cage-lifter 笼式升降机 Caiana cotton 凯阿那棉 cairngorm 烟晶 cajun fibre 卡琼强韧叶纤维 cake basket 饼篮 cake box 糕饼盒 cake breaker 滤饼破碎机 cake colouring ⾷物染⾊料 cake crusher 碎饼机 cake dish 糕点碟 cake finish machine 丝饼后处理机 cake fodder breaker 油饼饲料粉碎机 cake fork 点⼼叉 cake knife 糕饼⼑ cake machine 糕点⽣产设备 cake mold 蛋糕模⼦ cake pan 饼铮 cake plate 饼碟 cake salver 糕点托盘 cake spinning machine 丝饼纺丝机 cake stand 饼架 cake tea 茶砖 cake tin 饼模 cake tongs 饼钳 cake tray 饼碟 cake turner 翻饼铲 cake washing machine 丝饼⽔洗机 cake winder 丝饼络丝机 cake-cooky wrapping machine 糕饼裹包机 cake-spun viscose rayon 纺成丝饼的粘胶⼈造丝 cake-wrapper paper 糕点包装纸 caking coal 粘结煤 caking glass wool 粘结玻璃棉 Cal-cop 10 铜铵合剂 Cala-Line 卡拉-莱恩聚丙烯或聚⼄烯薄膜纤维 calabar bean 卡拉巴⾖ calabar skin 灰⽩⽑⽪ calabash gourd 葫芦 calabrere 卡勒布雷尔棉布 calabria cotton 卡拉布⾥亚棉 Calacloud 卡拉克劳德醋酯短纤 caladaris 卡勒达⾥条⼦平布 Calais 卡莱斯牌汽车 calamacho 意⼤利丝缎 calamanco 卡拉曼科亚⿇布 calamatta silk 卡拉马塔⽣丝 calamine 炉⽢⽯ Calamite 灭螨特 calamus oil 菖蒲油 calamus 菖蒲 calan tablet 卡兰⽚ Calan 开兰牌⼿表 calandria 排管式堆容器。
REGULAR ARTICLEExogenous nitric oxide improves antioxidative capacity and reduces auxin degradation in roots of Medicago truncatula seedlings under cadmium stressJin Xu&Wenying Wang&Hengxia Yin&Xiaojing Liu&Hong Sun&Qin MiReceived:19March2009/Accepted:21April2009/Published online:6May2009#Springer Science+Business Media B.V.2009Abstract The effects of nitric oxide(NO)on cadmium toxicity in Medicago truncatula seedlings were studied by investigating root growth and uptake of antioxidants, IAA and ions.Exposure to cadmium reduced root growth and NO accumulation,and increased the production of reactive oxygen species(ROS)in roots. Supplementation with NO improved root growth and reduced ROS accumulation in roots.The NO-scavenger cPTIO,the nitrate reductase(NR)inhibitor tungstate, and the NO synthase(NOS)inhibitor L-NAME all inhibited the accumulation of NO in roots and reversed the effects of NO in promoting the root growth and accumulation of proline and glutathione.Application of NO reduced auxin degradation by inhibiting the activity of IAA oxidase.Exogenous NO also enhanced the uptake of K+and Ca2+.These results suggest that NO improves cadmium tolerance in plants by reducing oxidative damage,maintaining the auxin equilibrium and enhancing ion absorption.Keywords Nitric oxide.IAA oxidase.Proline. Glutathione.Heavy metal tolerance.Medicago truncatulaIntroductionNitric oxide(NO),a free radical gas,is an important signaling molecule involved in developmental,hormonal and environmental responses of plants(Neill et al.2002; Durner and Klessig1999).NO accumulation in roots mediates auxin-induced gravitropic bending(Hu et al. 2005),lateral root formation(Correa-Aragunde et al. 2004),adventitious root growth(Tewari et al.2008)and root hair development(Lombardo et al.2006).NO also improves plant tolerance to both abiotic and biotic stresses(Hsu and Kao2004;Sun et al.2007;Zhang et al.2008;Vital et al.2008;Yang et al.2006).Numerous investigations have indicated that up-regulation of the antioxidant system is an early response of plants to environment stress(Vital et al. 2008).Plants have two classes of antioxidativePlant Soil(2010)326:321–330DOI10.1007/s11104-009-0011-4Responsible Editor:Juan Barcelo.J.Xu(*):X.LiuCenter for Agricultural Resources Research, Institute of Genetics and Developmental Biology, Chinese Academy of Sciences,Huaizhong RD286,Shijiazhuang050021,Chinae-mail:xujin@W.Wang:H.Yin:Q.MiDepartment of Biology,Qinghai Normal University, Wusi west RD38,Xining,ChinaH.Sun(*)School of Pharmacy,Hubei University of Traditional Chinese Medicine, Huangjiahu west RD1,Wuhan430061,Chinae-mail:shkshk1969@systems(Xu et al.2009):enzyme antioxidants,which can convert superoxide radicals into hydrogen peroxide, water and oxygen,and non-enzymatic antioxidants, which can directly detoxify oxygen free radicals and are intracellular non-enzymatic reactive oxygen species (ROS)scavengers.Enzyme antioxidants include super-oxide dismutase(SOD,EC 1.15.1.1),ascorbate peroxidase(APX,EC1.11.1.11)and catalase(CA T, EC1.11.1.6).Examples of non-enzymatic antioxidants are proline,ascorbic acid,and glutathione.In plants,NO inhibits the activity of CA T and APX(Clark et al. 2000).NO also upregulates the expression of enzymes involved in proline biosynthesis(Δ1-pyrroline-5-car-boxylate synthetase,or P5CS;Zhang et al.2008)and glutathione biosynthesis(gamma-glutamylcysteine syn-thetase,orγ-ECS;Innocenti et al.2007),suggesting that NO mediates the intracellular antioxidant system in plants.Cadmium(Cd)is one of the most toxic elements that negatively affects plant growth and development (Macek et al.2002).Many studies have attempted to clarify the mechanism of Cd toxicity in plants(Sanita di Toppi and Gabbrielli1999).Early studies showed that Cd inhibited nitrate reductase(NR)activity in the shoots;as a result,cadmium reduced nitrate absorption and transport from root to shoot(Hernandez et al. 1996).Accumulated Cd in plants seems to compete for the same transmembrane carriers as nutrients,such as potassium(K),calcium(Ca),magnesium(Mg),iron (Fe),manganese(Mn),copper(Cu)and zinc(Zn) (Rivetta et al.1997;Sanita di Toppi and Gabbrielli 1999).In this study,we examined the Cd-induced oxidative damage in the roots of M.truncatula seed-lings and showed that NO has a protective effect on stress symptoms.Our results indicate that(i)NO reduced Cd-induced oxidative damage by increasing the production of proline and total glutathione;(ii) exogenous NO reduces Cd accumulation and enhances the absorption of K+and Ca2+under Cd stress;and(iii) NO supplementation maintains auxin equilibrium in plants by reducing the activity of IAA oxidase. Materials and methodsPlant material and culture conditionsSeeds of M.truncatula were first treated with70% ethanol for30s,rinsed five times with sterile distilled water,and then allowed to germinate on double-layer filter paper wetted with distilled water.Germinated M. truncatula seedlings were transferred to Hoagland solution.The seedlings were grown for1day in Hoagland solution and subsequently transferred to fresh Hoagland solution that contained various con-centrations of CdSO4(0,50,100,200,250,or 300µM)with or without NO donor or NO eliminator treatments for48h.To determine whether NO was able to mediate the physiological responses of the M.truncatula seedlings to cadmium-induced stress, the NO donor sodium nitroprusside(SNP,100μM), the NO scavenger2-(4-carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide(cPTIO, 100μM),the NO synthetase(NOS)inhibitor N G-nitro-L-Arg-methyl ester(L-NAME,100μM),and the nitrate reductase(NR)inhibitor tungstate(100μM) were used.Cultures were maintained at22±3°C under a16-h photoperiod with a photosynthetic photon flux (PPF)density of45μmol m-2s-1provided by cool white fluorescent lamps.Determination of growth and ion accumulations Relative growth rate(RGR)was determined by the method described previously(Errabii et al.2007).For analysis of ion content,seedlings were oven-dried at 65-70°C for5-7d.50mg of dried plant tissues were ground up and digested in1ml of concentrated nitric acid for2-3d at room temperature.Samples were then boiled for1-2h until completely digested.After adding4ml of millipore-filtered deionized water and briefly centrifuging,the Cd,K and Ca contents of each sample were determined by atomic absorption spectrophotometer(HITACHI-170-10,Japan).Each experiment was repeated at least three times with six explants.The significance of the results were verified by Duncan’s test(P<0.05).Determination of proline accumulation and total glutathione contentThe free proline content was determined according to Bates et al.(1973).Total glutathione contents were measured according to a method modified from Anderson et al.(1995).Seedlings were ground and centrifuged for10min at10,000g at4°C.Reagent1 (110mM Na2HPO4,40mM NaH2PO4,15mM EDTA,0.3mM5,5’-dithiobis(2-nitrabenzoic acid),and0.04%BSA),Reagent2(1mM EDTA,50mM imidazole solution,and0.02%BSA),5%Na2HPO4 (pH7.5),1.5U glutathione reductase(GR),0.5ml of extract and9mM NADPHNa4were combined into a 3ml reaction.The reaction mixture was measured at 412nm at25°C.The significance of the results were verified by Duncan’s test(P<0.05).Detection of the accumulation of NO,ROS and H2O2 NO was monitored by incubating roots from3-day-old seedlings with15μM of the fluorescent probe4,5-diaminofluorescein diacetate(DAF-2DA)in20mM HEPES–NaOH(pH7.5).Thereafter,the roots were washed twice for5min with fresh buffer and viewed under a Leica laser scanning confocal microscope (excitation490nm;emission515nm).For ROS level detection,roots were washed and stained with25µM of5-(and-6)-chloromethyl-2′,7′-dichlorodihydrofluorescein diacetate,acetylester (CM-H2DCFDA)for25min.Roots were then washed thoroughly and viewed under a Leica laser scanning confocal microscope(excitation488nm; emission525nm).For localizing H2O2produced by M.truncatula roots,treated roots were immersed in1mg/ml of3-diaminobenzidine(DAB)-HCl(pH3.8)for5h and cleared by boiling in alcohol(95%,v/v)for5min. Photos were taken using a Carl Zeiss Imaging System.Six roots were analyzed in each set of experiments.Detection of plasma membrane integrityThe loss of plasma membrane integrity was evaluated using Evans blue staining(Baker and Mock1994).M. truncatula roots were stained in0.25%(v/v)of Evans blue solution for15min at room temperature,and subsequently rinsed three times with distilled water for10min.Root tips(0.5cm)were excised and soaked in N,N-dimethylformamide for1h at room temperature.The optical density was measured spectrophotometrically at600nm.Determination of IAA content and the activity of IAA oxidaseFor each sample,0.5g of fresh seedlings were ground and extracted for12h with80%(v/v)aqueous methanol containing1%polyvinylpyrolidone and1mM2,6-di-tert-butylpara-cresol at4°C.The samples were purified, and then endogenous IAA content was determined by an indirect enzyme-linked immunosorbent assay (ELISA)technique(Yang et al.2001).0.5g of fresh seedlings were ground in5ml of chilled phosphate buffer(0.06M,pH6.0),and then centrifuged for20min at4,000g at4°C.The supernatants were collected and IAA oxidase activity was assayed using6ml reaction mixtures containing 4ml of phosphate buffer(0.06M,pH6.0),0.5ml of gel fraction,0.3ml of MnCl2(1mM),0.6ml of DCP (1mM),and0.6ml of IAA(4mM).The enzyme activities were expressed as the percentage of IAA destroyed(the negative control without an enzyme fraction has100%IAA),which was calculated from the absorbance of Salkowski’s reagent at530nm (Faivre-Rampant et al.2000).Four biological repli-cates were analyzed for each treatment.The signifi-cance of the results were verified by Duncan’s test (P<0.05).ResultsNO supplementation improved the seedling growth and decreased the cadmium accumulationTo select an appropriate concentration for Cd exposure, we first investigated the effects of different concentra-tion of Cd on growth of M.truncatula seedlings.As shown in Fig.1,the effects of Cd on root growth varied with the different Cd concentrations used.Root growth decreased markedly as Cd concentratio in-creased.Seedlings exposed to200μM Cd exhibited substantial growth reduction,and root growth de-creased to21%of normal.After48h of exposure to 50µM Cd,root growth decreased to57%of normal compared to untreated control plants;hence,50μM was chosen as a sublethal dosage in the experiment.To examine the accumulation of NO,ROS and H2O2in Cd-stressed seedlings,we applied a DAF-2 DA fluorescence probe,CM-H2DCFDA fluorescence probe and DAB staining to detect in vivo of NO,ROS and H2O2levels,respectively.As shown in Fig.2,Cd stress decreased the production of NO and enhanced the accumulation of ROS and H2O2in roots of M. truncatula seedlings after48h of treatment.We also used Evans blue staining as an indicator for measuringplasma membrane integrity.The results of Evans blue staining indicated that Cd toxicity strongly reduced plasma membrane integrity,and therefore resulted in cell death in roots under Cd stress.In order to investigate the role of NO in cadmium tolerance of M.truncatula seedlings,the NO donor SNP and the NO scavenger cPTIO were applied to determine root growth and cadmium accumulation under cadmium stress.As shown in Figs.2,3and4, supplementation with100μM SNP improved plasma membrane integrity,promoted root growth and decreased Cd uptake in Cd-stressed M.truncatula seedlings.To confirm that the stimulative effect of SNP treatment was the result of NO accumulation,100μM of the NO scavenger cPTIO was applied alone or with pared to untreated control plants,the endogenous NO level was reduced by28%by cPTIO(Fig.2a),suggesting that the DAF-2DA-dependent fluorescence was related to the amount of endogenous NO.Supplementing with cPTIO inhibited root growth and increased the Cd accumu-lation under Cd stress;in addition,the inhibitory effects of cPTIO could be reversed by supplementing with SNP(Fig.2a).Application of the NR inhibitor tungstate and the NOS inhibitor L-NAME inhibited the production of NO in roots and reversed the protective effects of SNP on M.truncatula seedlings (Figs.2,3and4).Exogenous NO improves the antioxidative capacity by increasing the productions of proline and total glutathioneCd toxicity increased the accumulation of ROS and H2O2in roots of M.truncatula seedlings.Upon addition of the NO donor SNP,the production of ROS and H2O2decreased markedly(Fig.2).Since supplementation with SNP decreased the activities of the enzyme antioxidants CAT and SOD in M. truncatula seedlings(data not shown),we examined the levels of the non-enzyme antioxidants proline and glutathione.Our results indicated that NO supple-mentation increased the accumulation of proline and glutathione by134%and135%compared to Cd-treated seedlings,whereas application of the NO scavenger cPTIO,the NOS inhibitor L-NAME and the NR inhibitor tungstate reduced the levels of proline and glutathione(Fig.5).NO improved K and Ca uptakes in Cd-stressedM.truncatula seedlingsIon balance inside the cell is closely related to plant adaptation to environmental stress.Therefore,we compared the K and Ca levels of M.truncatulata seedlings under Cd stress.Figure6showed that K and Ca contents decreased by69.7%and74.8%,respec-tively,in M.truncatula seedlings after48h of Cd treatment.After SNP treatment,K and Ca levels were increased by129%and150%respectively compared to Cd-treated seedlings.Inhibition of NO accumula-tion by cPTIO,L-NAME and tungstate reversed the effects of NO on the ion absorption of M.truncatula seedlings.NO counteracts the inhibitory effects of cadmium toxicity on the IAA content by reducing the activity of IAA oxidase in M.truncatula seedlings Because NO improved seedling growth under Cd toxicity,we next examined the effects of NO on the IAA content of M.truncatula seedlings.Cd stress reduced IAA content by41.6%in M.truncatula seedlings and the IAA content was further reduced by 22.6%when supplemented with50μM Cd plus 100μM of cPTIO(Fig.7).Chaoui and Ferjani(2005) found that Cd-reduced growth could be associated with elevated IAA oxidase activity in peaseedlings.Therefore,we hypothesized that there appears to be a relationship between NO accumulation and the activity of IAA oxidase in plants subjected to cadmium stress.To test whether NO accumulation altered the activity of IAA oxidase,the NO donor SNP and NO inhibitors cPTIO,L-NAME and tungstate were used.Supplementing with 100μM SNP reduced the activity of IAA oxidase by 71%compared to Cd-treated seedlings,whereas,the NO inhibitor cPTIO,L-NAME and tungstate increased the activity of IAA oxidase by 125%,120%and 125%,respectively (Fig.8).DiscussionIn the present work,we provide evidence for the involvement of NO in cadmium tolerance.NO reduces the activity of IAA oxidase and increasesthe accumulation of proline and glutathione,resulting in improved seedling growth under cadmium stress.NO has also been found to provide a protective effect against Cd toxicity in rice leaves (Hsu and Kao 2004),sunflower leaves (Laspina et al.2005)and wheat roots (Singh et al.2008).Although Cd can induce an increase in scavenging enzyme activity,NO supple-mentation inhibits this effect (Beligni and Lamattina 2002;Singh et al.2008).Rodríguez-Serrano et al.(2009)found that Cd 2+treatment reduces the activity of Mn-SOD,Fe-SOD and Cu,Zn-SOD,however,semi-quantitative RT-PCR analysis indicate that Cd up-regulated the expression of Fe-SOD but down-regulated the Mn-SOD and Cu,Zn-SOD in pea plants,suggesting that Mn-SOD is regulated at transcriptional level,while Cu,Zn-SOD and Fe-SOD are regulated at both transcriptional and postranscriptional level under Cd stress.Though NO accumulation suppresses the activities of antioxidative enzymes,such as SOD,CATFig.2In situ fluorescence and histochemical localiza-tion showing the effects of the NO donor SNP and the NO scavengers cPTIO,tungstate,L-NAME on Cd-induced (a)NO production,(b)ROS accumulation,(c)H 2O 2level,and (d)loss of membrane integrity inMedicago truncatula roots.At each treatment,there are nine roots in each photo-graph in the followingorder:control,Cd (50μM),Cd (50μM)+SNP(100μM),Cd (50μM)+cPTIO (100μM),Cd(50μM)+cPTIO (100μM)+SNP (100μM),Cd (50μM)+tungstate (100μM),Cd (50μM)+tungstate(100μM)+SNP (100μM),Cd (50μM)+L-NAME (100μM),Cd (50μM)+L-NAME(100μM)+SNP (100μM)and APX,application of NO increases the production of non-enzymatic antioxidants,such as proline and glutathione.Proline and glutathione can directly detoxify oxygen free radicals,and glutathione is an important antioxidant and redox buffer when plants are under stress.Siripornadulsil et al.(2002)reported thatproline reduces cadmium stress not by sequestering cadmium but by reducing cadmium-induced free radical damage,thus maintaining a more reducing environment in the cell.NO thus both maintains intracellular antioxidative capacity and reduces oxida-tive damage.Supplementation with NO protectes plants against membrane damage due to lipid perox-idation and improves the activities of membrane transporters which exclude excess or toxic heavy metal ions from the cells of roots(Beligni and Lamattina 2002;Singh et al.2008),thereby reducing Cd accumulation in M.truncatula seedlings.Supply with NO donor SNP induces the accumulations of proline and glutathione in the seedlings of M.truncatula. Proline can decrease heavy metal toxicity by chelating metal ions(Farago and Mullen1979).Our previous study indicated that proline pretreatment increases cadmium tolerance in callus and regenerated shoots of Solanum nigrum(Xu et al.2009).High glutathione levels facilitate cell proliferation in roots and hence improve root growth under Cd stress(Mendoza-Cozatl and Moreno-Sanchez2006).These results indicated that exogenous NO enhanced the production of proline and glutathione and reduced the H2O2accumulation, thereby improving Cd tolerance in M.truncatula Fig.5Effects of100μM SNP,cPTIO,L-NAME or tungstate on the accumulation of proline and glutathione in Medicago truncatula seedlings after48h of exposure to50μM Cd.Each value is the mean of three replicates.V ertical bars represent±standard error.Different letters indicate significant differences,5%level, Duncan’s mutiple rangetestseedlings.Future study will analysis the levels of the other superoxides such as hydroxyl radical (OH .)and superoxide anion (O 2.-),and further elucidate the effects of different superoxides on the growth of Cd-stressed seedlings.There are conflicting results reported regarding the impact of Cd on NO accumulation.We found that Cd 2+treatment reduced NO accumulation in roots of Medicago seedlings.This result is consistent with the findings of Rodriguez-Serrano et al.(2006),Barroso et al.(2006)and Rodríguez-Serrano et al.(2009),and are contrary to the reports by Bartha et al.(2005),Kopyra et al.(2006),Groppa et al.(2008)and Besson-Bard et al.(2009)that Cd 2+mediates NO production in pea (Pisum sativum ),India mustard (Brassica juncea ),wheat (Triticum aesti-vum ),and Arabidopsis (Arabidopsis thaliana )roots and in Glycine max cell suspensions.The possibility is that we investigated the NO accumulation after 48h of Cd 2+treatment and not during the early stages of Cd 2+exposure,and endogenous NO might be produced within hours in seedlings and then reduced significantly in Cd-stressed seedlings.Our study suggested that Cd stress decreased NO level ,thereby promoting the accumulation of ROS and inducing oxidative damage.NO synthetase (NOS)and nitrate reductase (NR)are the most studied enzymatic sources of NO in plants (Graziano and Lamattina 2007).L-NAME,anFig.6Effects of 100μM SNP,cPTIO,L-NAME or tungstate on the uptake of K +and Ca 2+in Medicago truncatula seedlings under 50μM Cd stress for 48h.Each value is the mean of three replicates.Vertical bars represent ±standard error.Differ-ent letters indicate significant differences,5%level,Duncan ’s mutiple rangetestFig.8Effects of 100μM of SNP,cPTIO,L-NAME or tungstate on the activity of IAA oxidase of Medicago truncatula seedlings under 50μM of Cd stress for 48h.Each value is the mean of three replicates and vertical bars represent ±standard error.Different letters indicate significant differences,5%level,Duncan ’s mutiple range testL-arginine analogue,has been used to inhibit NOS in plants(Neill et al.2002).NR is a molybdenum-containing enzyme that catalyzes the formation of NO through nitrite reduction and tungstate is a molybdate analogue that suppresses the formation of an active NR in vivo(Deng et al.1989;Graziano and Lamattina2007).Both tungstate and L-NAME inhibited the NO accumulation in Cd-treated roots of M.truncatula seedlings,implying that Cd reduced the NO accumulation through both the NR and NOS pathways.Similar results were also observed in Cd-treated A.thaliana plants(Besson-Bard et al.2009)and pea plants(Rodríguez-Serrano et al.2009).At high concentration of Cd,supplementaion with NOS inhib-itor L-NAME suppressed NO accumulation both in roots and leaf dises of A.thaliana,suggesting that NO accumulation in Cd-treated seedlings might be catalyzed through a L-Arg-dependent route.Besson-Bard et al. (2009)found that NO production was not suppressed in the NR-null double mutant nia1nia2.However,we found that supplementation with NR inhibitor tungstate reduced NO accumulation significantly in Cd-treated Medicago seedlings.Therefore,further experiments will determine the source of NO in Cd-stressed plants.Cd toxicity inhibits the accumulations of K+and Ca2+in plants,disturbing intracellular ion homeostasis and exerting a toxic effect on plants(Rivetta et al. 1997).Zhao et al.(2004)reported that NO serves as a signaling molecule in the induction of salt tolerance by increasing the K+/Na+ratio,which is dependent on increased plasma membrane(PM)H+-A TPase activity in reed callus.Our study indicates that NO supple-mentation increases the absorption of Ca2+and K+in plants.Ca was a vital signaling molecular in plants responding to the stresses.NO was proved to interact with Ca2+channel in plants(Neill et al.2002; Wendehenne et al.2004).Cd2+treatment reduced the accumulation of Ca2+.Rodríguez-Serrano et al.(2009) found that exogenous supply of Ca2+decreased the accumulation of Cd2+in pea plants,suggesting that the competition between both elements for the same transporters.Rodríguez-Serrano et al.(2009)also found that Cd-induced down-regulation of Cu,Zn-SOD and reduction in activity of Cu,Zn-SOD could be reverted by Ca supply;Ca supply restored NO accumulation in Cd-treated plants by modulating NOS activity in plants,thereby reducing ROS accumulation and membrane damages.These results indicated that the increased Ca level by NO treatment is important for improving Cd tolerance in M.truncatula seedlings.K+ is an essential element that plays vital roles in various aspects of plant cell growth and metabolism and is needed in large quantities(Zhu et al.1998).Appropri-ate concentration of K is advantageous for the accumulation of divalent cations(Kahn and Hanson 1957).These results indicated that NO ameliorates the effects of Cd toxicity on NO-mediated intracellular ion equilibrium.Further research is needed to elucidate the molecular details regarding the exact cascade of changes that occur and genes or proteins expressed in response to NO in enhancing the ion uptake under Cd toxicity.Previous studies indicate that NO mediates auxin-induced processes in roots(Gouvea et al.1997;Hu et al.2005;Lombardo et al.2006).Auxin application to roots resulted in localized NO production during lateral root and root hair formation.These results suggest that NO is an important molecule operating downstream of auxin through a linear signaling pathway during root growth and development(Correa-Aragunde et al. 2007).Cd stress suppressed IAA accumulation by increasing the activity of IAA oxidase.In this study, we found that NO supplementation enhances the level of IAA in roots by reducing the activity of IAA oxidase.These results indicate that NO modulates auxin signaling pathway through mediating auxin-induced processes,and maintaining the auxin equilib-rium by reduing the activity of IAA oxidase in plants. To the best of our knowledge,our results are the first to show that NO reduces the activity of IAA oxidase,thus reestablishing the level of IAA as an adaptation to Cd stress.Further experiments will determine whether NO supplementation alters the expression of genes in-volved in auxin biosynthesis,metabolism and transport in plants.Acknowledgement The research was supported by the Knowledge Innovation Program of the Chinese Academy of Sciences(Grant no.0707013603).ReferencesAnderson MD,Prasad TK,Stewart CR(1995)Changes in isozyme profiles of catalase,peroxidase,and glutathione reductase during acclimation to chilling in mesocotyls of Maize seedlings.Plant Physiol109:1247–1257Baker CJ,Mock NM(1994)An improved method for monitoring cell death in cell suspension and leaf discassays using Evans blue.Plant Cell Tissue Org39:7–12.doi:10.1007/BF00037585Barroso JB,Corpas FJ,Carreras A,Rodriguez-Serrano M, Esteban FJ,Fernandez-Ocana A,Chaki M,Romero-Puertas MC,Valderrama R,Sandalio LM,del Rio LA (2006)Localization of S-nitrosoglutathione and expression of S-nitrosoglutathione reductase in pea plants under Cd stress.J Exp Bot57:1785–1794.doi:10.1093/jxb/erj175 Bartha B,Kolbert Z,Erdei L(2005)Nitric oxide production induced by heavy metals in Brassica juncea L.Czern.and Pisum sativum L.Acta Biol Szegediensis49:9–12 Bates LS,Waldren RP,Tear ID(1973)Rapid determination of free proline for water stress studies.Plant Soil39:205–207.doi:10.1007/BF00018060Beligni MV,Lamattina L(2002)Nitric oxide interferes with plant photooxidative stress by detoxifying reactive oxygen species.Plant Cell Environ25:737–748.doi:10.1046/ j.1365-3040.2002.00857.xBesson-Bard A,Gravot A,Richaud P,Auroy P,Duc C, Gaymard F,Taconnat L,Renou JP,Pugin A,Wendehenne D(2009)Nitric oxide contributes to cadmium toxicity in Arabidopsis by promoting cadmium accumulation in roots and by up-regulating genes related to iron uptake.Plant Physiol149:1302–1315.doi:10.1104/pp.108.133348 Chaoui A,Ferjani EE(2005)Effects of cadmium and copper on antioxidant capacities,lignification and auxin degradation in leaves of pea(Pisum sativum L.)seedlings.C R Biologies328:23–31.doi:10.1016/j.crvi.2004.10.001 Clark D,Durner J,Navarre DA,Klessig DF(2000)Nitric oxide inhibition of tobacco catalase and ascorbate peroxidase.Mol Plant Microbe Interact13:1380–1384.doi:10.1094/ MPMI.2000.13.12.1380Correa-Aragunde NM,Graziano ML,Lamattina L(2004) Nitric oxide plays a central role in determining lateral root development in tomato.Planta218:900–905.doi:10.1007/s00425-003-1172-7Correa-Aragunde N,Lanteri ML,Garcia-Mata C,Have A, Laxalt AM,Graziano M,Lamattina L(2007)Nitric oxide functions as intermediate in Auxin,Abscisic acid,and lipid signaling pathways.Plant Cell Monogr5:1861–1370 Deng M,Moureaux T,Caboche M(1989)Tungstate,a molybdate analog inactivating nitrate reductase,dereg-ulates the expression of the nitrate reductase structural gene.Plant Physiol91:304–309.doi:10.1104/pp.91.1.304 Durner J,Klessig DF(1999)Nitric oxide as a signal in plants.Curr Opin Plant Biol2:369–372.doi:10.1016/S1369-5266(99)00007-2Errabii T,Gandonou CB,Essalmani H,Abrini J,Idaomar M, Senhaji NS(2007)Effects of NaCl and mannitol induced stress on sugarcane(Saccharum sp.)callus cultures.Acta Physiol Plant29:95–102.doi:10.1007/s11738-006-0006-1 Faivre-Rampant O,Kevers C,Gaspar T(2000)IAA-oxidase activity and auxin protectors in nonrooting rac mutant shoots of tobacco in vitro.Plant Sci153:73–80.doi:10.1016/S0168-9452(99)00255-1Farago ME,Mullen WA(1979)Plants which accumulate metals.IV.A possible copper-proline complex from the roots of Armeria maritima.Inorg Chim Acta32:93–94.doi:10.1016/S0020-1693(00)91627-XGouvea CMCP,Souza JF,Magalhaes CAN,Martins IS(1997) NO releasing substances that induce growth elongation inmaize root segments.Plant Growth Regul21:183–187.doi:10.1023/A:1005837012203Graziano M,Lamattina L(2007)Nitric oxide accumulation is required for molecular and physiological responses to iron deficiency in tomato roots.Plant J52:949–960.doi:10.1111/j.1365-313X.2007.03283.xGroppa MD,Rosales EP,Iannone MF,Benavides MP(2008) Nitric oxide,polyamines and Cd-induced phytotoxicity in wheat roots.Phytochemistry69:2609–2615.doi:10.1016/j.phytochem.2008.07.016Hernandez LE,Carpena-Ruiz R,Garate A(1996)Alterations in the mineral nutrition of pea seedlings exposed to cadmium.J Plant Nutr19:1581–1598.doi:10.1080/ 01904169609365223Hsu YT,Kao CH(2004)Cadmium toxicity is reduced by nitric oxide in rice leaves.Plant Growth Regul42:227–238.doi:10.1023/B:GROW.0000026514.98385.5cHu X,Neill S,Tang Z,Cai W(2005)Nitric oxide mediates gravitropic bending in soybean roots.Plant Physiol 137:663–670.doi:10.1104/pp.104.054494Innocenti G,Pucciariello C,Gleuher ML,Hopkins J,de Stefano M,Delledonne M,Puppo A,Baudouin E,Frendo P(2007)Glutathione synthesis is regulated by nitric oxide in Medicago truncatula roots.Planta225:1597–1602.doi:10.1007/s00425-006-0461-3Kahn JS,Hanson JB(1957)The effect of calcium on potassium accumulation in corn and soybean roots.Plant Physiol 32:312–316.doi:10.1104/pp.32.4.312Kopyra M,Stachon-Wilk M,Gwozdz EA(2006)Effects of exogenous nitric oxide on the antioxidant capacity of cadmium-treated soybean cell suspension.Acta Physiol Plant28:525–536.doi:10.1007/s11738-006-0048-4 Laspina NV,Groppa MD,Tomaro ML,Benavides MP(2005) Nitric oxide protects sunflower leaves against Cd-induced oxidative stress.Plant Sci169:323–330.doi:10.1016/j.plantsci.2005.02.007Lombardo MC,Graziano M,Polacco J,Lamattina L(2006) Nitric oxide functions as a positive regulator of root hair development.Plant Signal Behav1:28–33Macek T,Mackova M,Pavlikova D,Szakova J,Truksa M, Singh Cundy A,Kotrba P,Yancey N,Scouten WH(2002) Accumulation of cadmium by transgenic tobacco.Acta Biotechnol22:101–106.doi:10.1002/1521-3846(200205) 22:1/2<101::AID-ABIO101>3.0.CO;2-N Mendoza-Cozatl DG,Moreno-Sanchez R(2006)Control of glutathione and phytochelatin synthesis under cadmium stress.J Theor Biol238:919–936.doi:10.1016/j.jtbi.2005.07.003Neill SJ,Desikan R,Clarke A,Hurst RD,Hancock JT(2002) Hydrogen peroxide and nitric oxide as signalling mole-cules in plants.J Exp Bot53:1237–1247.doi:10.1093/ jexbot/53.372.1237Rivetta A,Negrini N,Cocucci M(1997)Involvement of Ca2+-calmodulin in Cd2+toxicity during the early phases of radish(Raphanus satious L.)seed germina-tion.Plant Cell Environ20:600–608.doi:10.1111/ j.1365-3040.1997.00072.xRodriguez-Serrano M,Romero-Puertas MC,Zabalza A,Corpas FJ,Gomez M,del Rio LA,Sandalio LM(2006)Cadmium effect on oxidative metabolism of pea(Pisum sativum L.) roots:imaging of reactive oxygen species and nitric oxide。
1.indication 缺点2.test specimen 试样3.bar 棒材4.stock 原料5.billet 方钢,钢方坯6.bloom 钢坯 ,钢锭7.section 型材8.steel ingot 钢锭9.blank 坯料,半成品10.cast steel 铸钢11.nodular cast iron 球墨铸铁12.ductile cast iron 球墨铸铁13.bronze 青铜14.brass 黄铜15.copper 合金16.stainless steel 不锈钢17.decarburization 脱碳18.scale 氧化皮19.anneal 退火20.process anneal 进行退火21.quenching 淬火22.normalizing 正火23.Charpy impact text 夏比冲击试验24.fatigue 疲惫25.tensile testing 拉伸试验26.solution 固溶办理27.aging 时效办理28.Vickers hardness 维氏硬度29.Rockwell hardness 洛氏硬度30.Brinell hardness 布氏硬度31.hardness tester 硬度计32.descale 除污,除氧化皮等33.ferrite 铁素体34.austenite 奥氏体35.martensite 马氏体36.cementite 渗碳体37.iron carbide 渗碳体38.solid solution 固溶体39.sorbite 索氏体40.bainite 贝氏体41.pearlite 珠光体42. nodular fine pearlite/ troostite屈氏体43.black oxide coating 发黑44.grain 晶粒45.chromium 铬46.cadmium 镉47.tungsten 钨48.molybdenum 钼49.manganese 锰50.vanadium 钒51.molybdenum 钼52.silicon 硅53.sulfer/sulphur 硫54.phosphor/ phosphorus 磷55.nitrided 氮化的56.case hardening 表面硬化,表面淬硬57.air cooling 空冷58.furnace cooling 炉冷59.oil cooling 油冷60. electrocladding /plating电镀61.brittleness 脆性62.strength 强度63.rigidity 刚性 ,刚度64.creep 蠕变65.deflection 挠度66.elongation 延长率67.yield strength 折服强度68.elastoplasticity 弹塑性69.metallographic structure 金相组织70.metallographic test 金相试验71.carbon content 含碳量72.induction hardening 感觉淬火73.impedance matching 感觉淬火74. hardening and tempering调质75.crack 裂纹76.shrinkage 缩孔,松散77.forging 锻(件)78.casting 铸(件)79.rolling 轧(件)80.drawing 拉(件)81.shot blasting 喷丸(办理)82.grit blasting 喷钢砂(办理)83.sand blasting 喷砂(办理)84.carburizing 渗碳85.nitriding 渗氮86.ageing/aging 时效87.grain size 晶粒度88.pore 气孔89.sonim 夹砂90.cinder inclusion 夹渣91. lattice 晶格92. abrasion/abrasive/rub/wear/wearing resistance (property)耐磨性93. spectrum analysis 光谱解析94. heat/thermal treatment热办理95.inclusion 夹杂物96.seGREgation 偏析97.picking 酸洗,酸浸98.residual stress 节余应力99.remaining stress 节余应力100.relaxation of residual stress 除掉节余应力101.stress relief 应力开释A R steel 高温度锰钢abnormal steel失常钢;非正常钢aluminium-steel cable钢铝电缆aluminized steel涂铝钢aluminum nickel steel铝镍钢anchored steel trestle锚固式钢栈桥angle steel ruler 钢角尺anisotropic steel各向异性钢片anisotropy silicon steel各向异性硅钢片annealed sheet steel 退火薄钢板annealed steel 退火钢 ; 韧钢anti-creeping angle steel防爬角铁anticorrosive aluminium-coated steel wire防蚀镀铝钢丝arc-furnace steel电炉钢arc-welded steel pipe电弧焊接钢管area of steel 钢筋断面面积 ; 钢筋截面面积area of structural steel型钢截面积Armco aluminized steel阿姆柯渗铝钢armco aluminized steel表面浸镀铝钢Armco stabilized steel阿母柯稳固化钢Armco steel 阿姆柯软钢armco steel 不硬化钢austenilic Ni-Cr stainless steel 奥氏体镍铬不锈钢austenitic alloy steel奥氏体合金钢austenitic chromium-nickel steel奥氏体铬-镍钢austenitic clad steel奥氏体复合钢austenitic heat-resistance steel奥氏体耐热钢austenitic manganese steel 奥氏体锰钢 ; 哈德菲尔德锰钢austenitic stainless steel 奥氏体不锈钢austenitic steel奥氏体钢automatic steel易切削钢automatic(al) steel易切削钢axle shaft steel 车轴钢axle steel 车轴钢钢铁业的英语词汇 1Carbon Steel Plate 碳素钢板 ; 碳素钢板和型材Carbon Tool Steel 碳工具钢 ; 碳素工具钢Carbon-Containing Alloy Steel含碳合金钢Carbon-Free Stainless Steel 无碳不锈钢Carbon-Free Steel 无碳钢Carburizing Steel 碳钢Case-Hardened Steel 表面渗碳硬化钢; 渗碳钢Bolts & Nuts For Steel Slotted角钢螺丝Cast Alloy Steel Piston 合金铸钢活塞Cast Iron Electrode With Steel Core钢芯铸铁焊条Cast Stainless Steel 锻造不锈钢Cast Steel 铸钢件Cast Steel Plate 铸钢板Cast(Carbon) Steel 铸钢Cast-In Steel Bush 铸入钢套Casement Section Steel 窗框钢Cast-Steel 铸钢Cast-Steel Crossing 铸钢辙叉Cast-Steel Pipe 铸钢管Cast-Steel Separator 铸钢隔板 ; 铸钢横梁Cast-Steel Shot 钢砂Cast-Steel Yoke 铸钢轭Cellular Steel Floor 格型钢楼板Cement-And-Grouted Steel Bolt钢筋沙浆锚杆Cemented Steel 表面硬化钢Cemented Templet Steel 渗碳样板钢Centra Steel 球状石墨铸钢Centrifugal Steel 离心铸钢Centrifuge(D) Steel 离心锻造钢Channel Steel 槽钢Channel-Section Steel 槽形钢Channeled-Steel Wheel Rim 槽钢制轮辋Checkered Steel Plate 花纹钢板Chemical Capped Steel 化学封顶钢Chilled Cast Steel 冷硬铸钢Chilled Hardened Steel 淬火钢Chilled Steel 淬火钢 ; 淬硬钢Chisel Steel 凿钢 ; 凿子钢Chrome Hardened Steel 铬硬化钢Chrome Steel 铬钢Chrome-Carbon Steel 铬碳钢Chrome-Manganese Nitrogen Steel铬锰氮钢Chrome-Manganese-Silicon Alloy Steel铬锰硅合金钢Chrome-Molybdenum Steel铬钼钢Chrome-Nickel Steel 铬镍钢Chrome-Plated Steel Nozzle 镀铬钢喷管Chrome-Silicon Steel 铬硅钢Chrome-Tungsten Steel 铬钨钢Chrome-Vanadium Steel 铬钒钢Chromemanganese Steel 铬锰钢Chromium Nickel Steel 高强度耐蚀铬镍钢Chromium Stainless Steel 铬不锈钢Chromium Steel 铬钢Chromium-Molybdenum Steel铬钼钢Chromium-Nickel Steel 铬镍钢Chromium-Plated Mild Steel Animal Chain镀铬软钢动物链条Chromium-Tungstenvanadium Steel铬钨钒钢Chromiummanganese Steel 铬锰钢Chromiumnickel Austenite Stainless Steel铬镍奥氏体不锈钢Chromiumplated Thin-Walled Steel Cylinder Liner钢质薄壁镀铬汽缸衬套Chromized Steel 渗铬钢Circular Steel Support 环形金属支架Clad Sheet Steel 包层薄钢板Clad Steel 包层钢 ; 双层钢 ; 双金属钢Clad Steel Plate 复合钢板Cladding Steel 包覆钢Cladding Steel Sheet 复合钢板Clean Steel 纯净钢 ; 干净钢Closed Die Steel 闭锻模钢Coarse-Grained Steel 粗晶粒钢Coated Steel 镀层钢板Coating Steel Pipe 镀锌钢管Cobalt High Speed Steel 钴高速钢Cobalt-Chromium Steel钴铬钢Coiled Steel 成卷带钢Cold Forging Steel 冷锻钢Cold Pressed Steel 冷压钢Cold Reduced Steel 马口铁卷板Cold Rolled Ribbed Steel Bar 冷轧竹节钢筋Cold Rolled Sheet Steel 冷轧薄钢板Cold Twisted Steel Bar 冷扭钢筋Cold-Banded Steel Pipe 冷箍钢管Cold-Drawn Steel 冷拔钢 ; 冷拉钢Cold-Drawn Steel Pipe 冷拉钢管Cold-Drawn Steel Tube 冷拉钢管Cold-Finished Steel 冷加工精整钢Cold-Pressed Sheet Steel 冷压薄钢板Cold-Pressed Steel Plate 冷压钢板Cold-Pressing Quality Steel 冷弯钢Cold-Roll Steel Sheets 冷轧钢板Cold-Rolled (Cold Rolled) Steel 冷轧钢Cold-Rolled Carbon Spring Steel 冷轧弹簧碳钢Cold-Rolled Carbon Steel 冷轴碳钢Cold-Rolled Steel Strip 冷轧带钢Cold-Shaping Steel 冷变形钢Cold-Short Steel 冷脆钢Cold-Steel 利器Cold-Strip Steel 冷轧带钢Cold-Strip Steel Rolling Mill冷轧带钢机;带钢冷轧机Cold-Worked Steel 冷作钢Cold-Working Steel 冷作钢Coldshort Steel 冷脆钢Color Stainless Steel 彩色不锈钢Colored Stainless Steel 彩色不锈钢Column With Steel Hooping螺旋箍筋柱Combined Steel And Concrete Column钢骨混凝土柱Comminuted Steel Shot钢粉Common Straight Carbon Steel 一般碳素钢Complex Alloy Steel 多元合金钢 ; 合金钢碳素结构钢Carbon Construction Steel碳素工具钢Carbon Tool Steel合金结构钢Alloy Construction Steel合金工具钢Alloy Tool Steel高速工具钢High Speed Tool Steel弹簧钢Spring Steel不锈耐热钢Stainless & Heat Resisting Steel轴承钢Bearing Steel工字钢Ibeam一般圆钢Round Steel槽钢Channel扁钢Flat Steel Bar等边角钢Equal Angle Steel不等边角钢Unequal Angle Steelh型钢Hbeam钢坯Billet,Bloom钢筋混凝土用变形钢筋High Strength Deformed Steel Bar 窗框钢Window Sash中厚钢板Steel Plate热轧钢板(卷)Hot Rolled Steel Coil冷轧钢板(卷)Cold Rolled Steel Sheel or Coil镀锌钢板(卷)Galvanized Steel Coil钢带Steel Strip线材Wire Rod Coil无缝钢管Seamless Steel Pipe锅炉管Boiler Tube石油套管Pipe For Oil Field高压管Pressure Pipe高压气瓶用管Tube for High-Pressure Vessel地质钻探用管Seamless Steel Pipe For Geological Drill In 镀锌焊管Galvanized Welded Pipe石油钻探杆Drill Pipe For Oil Field铸铁管Cast Iron Pipe重轨Heavy Rail轻轨Light Rail炼钢生铁Basic Pig Iron锻造生铁foundry Pig Iron一般圆钢钉Round Nails for General Uses镀锌底碳钢丝Galvanized Low Carbon Steel Wire.钢丝绳Steel Wire Rope钢绞线Stranded Wire预应力混凝土钢丝Steel Wire For Prestressed Concrete 窗纱Window Screen车轮Wheel Tyre轮筛Wheel Circleiron and steel industry钢铁工业ironworks铁厂foundry锻造车间steelworks, steel mill钢厂coking plant炼焦厂electrometallurgy电冶金学powder metallurgy粉末冶金学blast furnace 鼓风炉mouth, throat炉口hopper, chute 料斗stack 炉身belly 炉腰bosh 炉腹crucible 炉缸slag tap 放渣口taphole 出铁口 ,出渣口pig bed 铸床mould 铸模(美作 :mold)tuyere, nozzle 风口ingot mould锭模(美作:ingot mold) floor平台hearth炉底charger 装料机ladle 铁水包 ,钢水包dust catcher 除尘器washer 清洗塔converter转炉hoist 卷扬机compressor 压缩机tilting mixer可倾式混铁炉regenerator蓄热室heat exchanger 热交换器gas purifier 煤气净化器turbocompressor涡轮压缩机burner烧嘴cupola 化铁炉 ,冲天炉emptier排空装置trough铁沟渠,排渣沟skip 料车rolling mill轧机,轧钢机blooming mill初轧机roller辊bed 底座rolling-mill housing轧机机架drawbench拔管机,拉丝机drawplate拉模板shaft furnace竖炉refining furnace精髓炉reverberatory furnace反射炉hearth furnace床式反射炉firebrick lining耐火砖衬retort反应罐muffle马弗炉roof, arch 炉顶forge 锻造press 压锻pile hammer打桩锤drop hammer落锤die 拉模blowlamp吹炬(美作 :blowtorch) crusher 破碎机iron ore铁矿石coke 焦炭bauxite 铁钒土alumina铝cryolite冰晶石flux 熔剂limestone flux石灰石溶剂haematite赤铁矿(美作 :hematite) gangue 脉石cast iron 铸铁cast iron ingot铸铁锭slag 炉渣soft iron软铁pig iron 生铁wrought iron熟铁iron ingot铁锭puddled iron搅炼熟铁round iron圆铁scrap iron 废铁steel 钢crude steel 粗钢mild steel, soft steel软钢,低碳钢hard steel 硬钢cast steel 坩埚钢 ,铸钢stainless steel 不锈钢electric steel电工钢,电炉钢high-speed steel 高速钢moulded steel铸钢refractory steel热强钢,耐热钢alloy steel 合金钢plate, sheet薄板corrugated iron瓦垅薄钢板tinplate, tin马口铁finished product成品,产品semifinished product半成品,中间产品ferrous products铁制品coiled sheet 带状薄板bloom 初轧方坯metal strip, metal band铁带,钢带billet坯锭,钢坯shavings 剃边profiled bar异型钢材shape, section 型钢angle iron 角钢frit烧结wire线材ferronickel镍铁elinvar 镍铬恒弹性钢ferrite铁氧体,铁醇盐cementite渗碳体,碳化铁pearlite珠光体charging, loading装料,炉料fusion, melting, smelting熔炼remelting再融化,重熔refining 精髓casting 出铁to cast 出铁tapping 出渣 ,出钢 ,出铁to insufflate, to inject注入heating 加热preheating预热tempering回火temper回火hardening 淬水annealing 退火reduction还原cooling 冷却decarbonization, decarburization脱碳coking 炼焦slagging, scorification造渣carburization渗碳case hardening 表面硬化cementation渗碳fritting, sintering烧结puddling搅炼pulverization粉化,雾化nitriding渗氮alloy 合金floatation, flotation浮选patternmaking制模moulding成型(美作 :molding) calcination 煅烧amalgamation汞齐化rolling轧制drawing拉拔extrusion 挤压wiredrawing拉丝stamping, pressing 冲压die casting 拉模锻造forging 锻造turning车削milling铣削machining, tooling加工autogenous welding, fusion welding氧炔焊arc welding 电弧焊electrolysis 电解trimming清理焊缝blowhole气孔钢铁业的英语词汇 2Abnormal Steel失常钢;非正常钢Aluminum-steel Cable钢铝电缆Aluminized Steel 涂铝钢Aluminum Nickel Steel铝镍钢Anchored Steel Trestle 锚固式钢栈桥Angle Steel Ruler 钢角尺Anisotropic Steel各向异性钢片Anisotropy Silicon Steel各向异性硅钢片Annealed Sheet Steel 退火薄钢板Annealed Steel 退火钢 ; 韧钢Anti-Creeping Angle Steel 防爬角铁Anticorrosive Aluminum-Coated Steel Wire防蚀镀铝钢丝1.Arc-Furnace Steel 电炉钢Arc-Welded Steel Pipe 电弧焊接钢管Area Of Steel 钢筋断面面积 ; 钢筋截面面积Area Of Structural Steel 型钢截面积Armco Aluminized Steel阿姆柯渗铝钢Armco Aluminized Steel表面浸镀铝钢Armco Stabilized Steel 阿母柯稳固化钢Armco Steel 阿姆柯软钢Armco Steel 不硬化钢Austenitic Ni-Cr Stainless Steel 奥氏体镍铬不锈钢Austenitic Alloy Steel奥氏体合金钢Austenitic Chromium-Nickel Steel奥氏体铬-镍钢Austenitic Clad Steel 奥氏体复合钢Austenitic Heat-Resistance Steel 奥氏体耐热钢Austenitic Manganese Steel奥氏体锰钢;哈德菲尔德锰钢Austenitic Stainless Steel 奥氏体不锈钢Austenitic Steel 奥氏体钢Automatic Steel易切削钢Automatic(Al) Steel易切削钢Axle Shaft Steel 车轴钢Axle Steel 车轴钢Bain tic Steel 贝氏体钢Balanced Steel 半冷静钢Ball Bearing Steel Strip 滚珠轴承钢带Ball Race Steel 滚珠轴承圈钢Ball-Bearing Steel 滚珠轴承钢Bamboo Steel 竹节钢筋Band Steel 带钢Banding Steel 带钢 ; 箍钢Bar Steel 棒钢 ; 条钢Basic Bessemer Steel 碱性贝氏转炉钢Basic Bessemer Steel 碱性转炉钢Basic Bessemer Steel Converter 碱性转炉钢转炉Basic Converter Steel 碱性转炉钢Basic Electric Furnace Steel 碱性电炉钢Basic Open Hearth Steel 碱性平炉钢Battened Steel Column双肢钢Beam Steel Plate 大梁钢板Beam With Compression Steel双筋梁Bearing Quality Steel 滚珠钢Bearing Steel 轴承钢Bent Steel 挠曲钢筋Bessemer Low Carbon Steel 酸性转炉低碳钢Bessemer Mild Steel 贝色麦钢Bessemer Steel 贝色麦钢 ; 酸性钢Best Plough Steel Wire 铅淬火高强度钢丝Billet Steel 坯段钢 ; 钢坯 ; 短条钢Binary Steel 二元合金钢Black Mild Steel Carriage Bolt And Nut黑铁马车螺丝闩Black Mild Steel Fish Bolt And Nut黑铁鱼尾螺丝闩Black Mild Steel Flat Head Rivet 黑铁扁头铆钉Black Mild Steel Flat Head Tin men Rivet黑铁号头铆钉Black Mild Steel Hexagonal Nut 黑铁六角螺丝帽Black Mild Steel Pan Head Rivet 黑铁锥头铆钉Black Mild Steel Round Head Rivet 黑铁圆头铆钉Black Mild Steel Square Nut 黑铁四方螺丝帽Black Steel Pipe 黑钢板Black Steel Sheet 黑钢皮Black Tin Steel 黑钢板Blister Steel 泡钢 ; 泡面钢Abnormal Structure Steel失常组织钢Abrasion-Resistant Steel 耐磨钢Blue Steel 蓝钢Bluing Of Steel 钢加蓝Boiler Steel 锅炉钢 ; 锅炉钢板Boiler Steel Plate 锅炉钢板Boiling Steel 沸腾钢Bolting Steel 螺栓钢Blue Planished Steel 发蓝薄钢板Bloom Steel 初轧钢 ; 钢坯Boron Steel 硼钢Brass Fitting Of Steel Window钢窗铜配件Brass-Plated Mild Steel Round Bead Wood Screw 镀黄铜圆头软钢木螺丝Brass-Plated Mild Steel Round Head Machine Screw 镀黄铜圆头软钢机器螺丝Brass-Plated Steel Wire 镀黄铜钢丝Brearley Steel 布里阿雷不锈钢; 布氏高铬钢Bridge Steel 桥梁钢Bright And Black Mild Steel Flat Head Tinmen Rivet 号头铆钉Bright Mild Steel Cotter Pin 光明软钢开尾销Bright Mild Steel Countersunk Head Wood Screw 光明软钢平头木螺丝Bright Mild Steel Hexagonal Nut 光明软钢六角螺丝帽Bright Mild Steel Hexagonal Nuts 六角光螺帽; 六角螺丝帽Bright Mild Steel Hexagonal Pressed Nut光明软钢六角冲压螺丝帽Bright Mild Steel Round Head Wood Screw光明软钢圆头木螺丝Bright-Drawn Free Cutting Steel光拉自由切削钢Bright-Drawn Steel 光明拉拔钢材Bright-Finished Steel 光明精整钢Bristol Steel Belt Lacing 布瑞斯涛钢带接头Bronze Steel 镀青铜钢Building Of Steel Frame Construction钢框架房屋Bulb Steel 球扁钢Bulb-Rail Steel 球头丁字钢Burned Steel 过烧钢Calorized Steel 涂铝钢 ; 渗化钢Calorizing Steel 铝化钢Capped Steel 加盖钢 ; 半冷静钢Carbon And Low-Alloy Steel Vessels 碳钢及低合金钢容器Carbon Content Of Steel 钢含碳量Carbon Molybdenum Steel钼碳钢Carbon Steel 碳钢 ; 碳素钢Carbon Steel Covered Electrode 碳钢焊条Carbon Steel Plate2. 中文名英文名质量描述6063 6063 Extrusion-pntd 镀漆6061 6061 Extrusion-pntd 镀漆生铝件Tense 含铁〈 2% 生杂铝Mixed Cast 含铁〈 2% 旧铝片Old Sheet-Taint/Tabor 干净新铝片Tough-Taboo 干净切碎铝Shredded Aluminum 干净汽车切片铝Shredded Auto Aluminum 含铝 70% 带铁铝Irony Aluminum 需注明含量356 轮毂铝356 Auto Wheels 干净铝水箱Aluminum Radiator 干净带皮铝线Acsr/Ins 需注明含量铜铝水箱Al/Cu Radiator 干净易拉罐UBC-Baled/Densified 干净铝变压器Aluminum Transformer铝壳马达Aluminum Case Motor1#光明铜线#1 Bare Bright 干净1#铜#1 Candy 含铜 98% 2#铜#2 Birch/Cliff 含铜 94-96% 1#火烧线#1 Burnt Wire 含铜〉 97% 1#铜米#1 Cu-chop1#铜线缆#1 Insulated Wire 含铜 72%2#铜线缆#2 Insulated Wire 含铜 50-55% 1#2#混杂铜线缆#1#2 Mixed Wire 含铜 65% 黄杂铜Yellow Brass-Honey 干净干净铜水箱Ocean Auto Radiator 干净(牛粪 )切碎电机Shredded Pickings 含铜 18-20% 混杂电机Electric Motors 含铜 8-12% 304 炉料304 Sabot 18-8304 可利用料304 Reusable 板或杆316 可利用料316 Reusable 板或杆3.1、烧结 sintering粉末或压坯在低于主要组分熔点的温度下的热办理,目的在于经过颗粒间的冶金联合以提升其强度。
ORIGINAL ARTICLECadmium inhibits the induction of high-affinity nitrate uptake in maize (Zea mays L.)rootsCecilia Rizzardo •Nicola Tomasi •Rossella Monte •Zeno Varanini •Fabio F.Nocito •Stefano Cesco •Roberto PintonReceived:28May 2012/Accepted:17July 2012ÓSpringer-Verlag 2012Abstract Cadmium (Cd)detoxification involves gluta-thione and phytochelatins biosynthesis:the higher need of nitrogen should require increased nitrate (NO 3-)uptake and metabolism.We investigated inducible high-affinity NO 3-uptake across the plasma membrane (PM)in maize seedlings roots upon short exposure (10min to 24h)tolow Cd concentrations (0,1or 10l M):the activity and gene transcript abundance of high-affinity NO 3-trans-porters,NO 3-reductases and PM H ?-ATPases were analyzed.Exposure to 1mM NO 3-led to a peak in high-affinity (0.2mM)NO 3-uptake rate (induction),which was markedly lowered in Cd-treated roots.Plasma membrane H ?-ATPase activity was also strongly limited,while internal NO 3-accumulation and NO 3-reductase activity in extracts of Cd treated roots were only slightly lowered.Kinetics of high-and low-affinity NO 3-uptake showed that Cd rapidly (10min)blocked the inducible high-affin-ity transport system;the constitutive high-affinity transport system appeared not vulnerable to Cd and the low-affinity transport system appeared to be less affected and only after a prolonged exposure (12h).Cd-treatment also modified transcript levels of genes encoding high-affinity NO 3-transporters (ZmNTR2.1,ZmNRT2.2),PM H ?-ATPases (ZmMHA3,ZmMHA4)and NO 3-reductases (ZmNR1,ZmNADH:NR ).Despite an expectable increase in NO 3-demand,a negative effect of Cd on NO 3-nutrition is reported.Cd effect results in alterations at the physiologi-cal and transcriptional levels of NO 3-uptake from the external solution and it is particularly severe on the inducible high-affinity anion transport system.Further-more,Cd would limit the capacity of the plant to respond to changes in NO 3-availability.Keywords ATPase ÁCadmium ÁNitrate reductase ÁNitrogen acquisition ÁNitrate transporters NRT Abbreviations Cd Cadmium GDH Glutamate dehydrogenaseNO 3-Nitrate NR Nitrate reductaseElectronic supplementary material The online version of this article (doi:10.1007/s00425-012-1729-4)contains supplementary material,which is available to authorized users.C.Rizzardo ÁF.F.NocitoDipartimento di Scienze Agrarie e Ambientali,Universita`degli Studi di Milano,Via Celoria 2,20133Milan,Italy e-mail:cecilia.rizzardo@unimi.it F.F.Nocitoe-mail:fabio.nocito@unimi.itN.Tomasi ÁR.Monte ÁR.Pinton (&)Dipartimento di Scienze Agrarie e Ambientali,Universita`degli Studi di Udine,Via delle Scienze 208,33100Udine,Italy e-mail:roberto.pinton@uniud.it N.Tomasie-mail:nicola.tomasi@uniud.it R.Montee-mail:rossella.monte@uniud.itZ.VaraniniDipartimento di Biotecnologie,Universita`degli Studi di Verona,Villa Lebrecht,Via della Pieve 70,37029San Floriano,VR,Italye-mail:zeno.varanini@univr.itS.CescoFacolta`di Scienze e Tecnologie,Libera Universita `di Bozen-Bolzano,Piazza Universita`1,39100Bozen-Bolzano,Italy e-mail:stefano.cesco@unibz.itPlantaDOI 10.1007/s00425-012-1729-4PM Plasma membraneSO42-SulfateIntroductionMultiple evidence suggests that coping with cadmium(Cd) toxicity should require increased nitrate(NO3-)uptake and metabolism.Firstly,Cd detoxification in higher plants mainly occurs through phytochelatins,which are N-and S-containing peptides(Clemens2006).Their synthesis leads to a depletion of the glutathione pool and to a de-repression of sulfate(SO42-)uptake(Nocito et al.2002), which in turn is known to be linked to NO3-uptake and metabolism,as described in maize cells or in barley and spinach plants(Clarkson et al.1989,1999;Prosser et al. 2001).Secondly,synthesis of phytochelatins requires glu-tamate,but exposition to Cd inhibits glutamine synthetase and glutamine oxoglutarate aminotransferase so that both leaves and roots of Cd-treated plants tend to accumulate ammonium and deplete their glutamate pool.As a conse-quence,an increase in glutamate synthesis via glutamate dehydrogenase(GDH)activity has been observed in dif-ferent plant species such as barley,bean,maize and rice (Boussama et al.1999a,b;Gouia et al.2000,2003;Astolfiet al.2004;Lee et al.2010):NO3-could help avoiding ammonium toxicity and favor glutamate synthesis(Britto and Kronzucker2002).Finally,the severity of Cd toxicity also depends on the plant N-status and an adequate rate of NO3-uptake may consequently facilitate Cd detoxifica-tion:N-deficient barley plants treated with Cd show dif-ferent metabolite pools(e.g.lower phytochelatins content), enzymatic activity and gene transcription levels if com-pared to N-sufficient plants treated with Cd(Finkemeier et al.2003).It has also been recently suggested that Cd tolerance may involve a preferential accumulation of NO3-in Arabidopsis roots(Li et al.2010).Despite the expectable increase in NO3-demand,a general negative effect of Cd on NO3-nutrition in higher plants has been reported(Sanita`di Toppi and Gabbrielli 1999).Cd exposure lowers NO3-accumulation and assimilation both in the root and in the shoot of bean and tomato(Ouariti et al.1997;Gouia et al.2000).At the enzymatic level,Cd decreases protein amount and activity of nitrate reductase(NR)in bean and maize,probably as a consequence of a general depression in protein synthesis (Boussama et al.1999b;Gouia et al.2003).However,it is often difficult to separate the direct effect of Cd itself from the secondary effect caused by the cellular response to a toxic accumulation of the heavy metal,since experimental conditions frequently include high Cd concentrations(up to the millimolar range)or prolonged exposures of the plants to the heavy metal(some days)as well as additional stress factors,e.g.,S-deficiency(Astolfiet al.2004).Notwithstanding the contradiction between the expected importance of NO3-uptake for Cd detoxification and the proved negative effect of Cd on NO3-nutrition,the effect of Cd exposure on the mechanisms of the anion’s uptake across the plasma membrane(PM)by NO3-transporters has not been studied in detail.Indeed,a decrease in NO3-depletion from the external solution,as well as an inhibi-tion of NR at the physiological level,has been described in barley or Pisum sativum(25–50l M Cd for up to72h or 10day;Hernandez et al.1997;Boussama et al.1999a),but the activity of NO3-transporters has not been monitored. At the molecular level,it has been observed in Arabidopsis that Cd(5or50l M Cd for up to30h)can rapidly alter the transcript levels of genes encoding NO3-transporters(e.g. AtNRT1.1,AtNRT2.1,AtNRT2.2),as well as NO3-reduc-tases(AtNR1,AtNR2)and PM proton pumps(PM H?-ATPases; e.g.AtAHA2,AtAHA5,AtAHA10,AtAHA11; Herbette et al.2006).NO3-uptake across the root PM represents a complex process with some peculiar characteristics shared by dif-ferent plant species,such as:(1)a localized rapid induction of high-affinity transporters activity upon supply of the anion,observed at both the transcript and the protein level (Hole et al.1990;Miller et al.2007;Wirth et al.2007),(2) a systemic negative feedback on high-affinity transporters exerted by the intermediate products of NO3-assimilation (e.g.nitrite,ammonium,glutamine,asparagine,arginine; Fraisier et al.2000;Vidmar et al.2000;Loque et al.2003), and(3)a dependence of NO3-transport on the electro-chemical gradient generated by the activity of the PM H?-ATPases(Miller and Smith1996).The induction of high-affinity NO3-transport is there-fore considered one of thefirst steps of the complex response to external NO3-with the anion acting not only as a nutrient,but also as a signal eliciting the rapid gene expression of transporters and metabolism enzymes(Krouk et al.2010).High-affinity NO3-transporters have also been suggested to play a role in root morphology(Little et al.2005;Remans et al.2006)and plant growth(Orsel et al.2004;Katayama et al.2009)in Arabidopsis and rice.Thus,it appears interesting to assess whether Cd might affect the induction of high-affinity NO3-transport across the PM.In the present work,5-day-old maize seedlings were exposed to1mM NO3-for up to24h(induction)in the presence or absence of low Cd concentrations(1or 10l M Cd)and the induction of high-affinity NO3-uptake was monitored;concomitantly,uptake kinetics,NO3-accumulation and reduction rate,and PM H?-ATPase activity were measured.The transcript levels of the main genes involved in NO3-uptake and reduction were also analyzed.PlantaMaterials and methodsPlant material and growth conditionsMaize seeds(Zea mays L.cv.Cecilia;Pioneer Hi-Bred Italia Srl,Pieve Delmona,CR,Italy)were germinated over an aerated0.5mM CaSO4solution at27°C in the dark.After 3days,seedlings were transferred into an aerated solution containing0.5mM CaSO4(day/night photoperiod16/8,light intensity220l mol photons m-2s-1,temperature(day/ night)25/20°C,RH70–80%).After2days,seedlings were transferred for NO3-uptake induction to a nutrient solution (NS)containing(mM)KNO31,NH4H2PO40.025,CaSO4 0.4,KH2PO40.087,MgSO40.1,KCl0.005,FeSO40.01, H3BO30.0025,MnSO40.0002,ZnSO40.0002,CuSO4 0.00005,H2MoO40.00005and with0(induced),1or10l M CdSO4for0,4,8,12or24h.In the NS for non-induced plants,KNO3was replaced by K2SO40.5mM. Measurement of net high-affinity NO3-uptakeand calculation of kinetic parametersRoots of intact seedlings were immersed in40mL of a con-stantly agitated and aerated solution containing0.5mM CaSO4and0.2mM uptake was measured as NO3-depletion from the solution per unit of time(Cataldo et al.1975),removing samples(0.2mL)for NO3-determi-nation every2min for10min,span time during which uptake had a linear trend.Aliquots of0.2mL were mixed thoroughly with0.8mL of5%(w/v)salicylic acid in concentrated H2SO4.After20min incubation at room temperature,19mL of2M NaOH was added.Samples were cooled to room temperature and NO3-concentration was determined spec-trophotometrically by measuring the absorbance at412nm.Kinetic parameters of the high-affinity NO3-uptake sys-tem(V max and K m)were calculated in the0.15–0.5concen-tration range.Uptake rates were measured as described above except that the uptake solution contained0.125,0.15,0.2,0.3, 0.5,1,2,5or10mM KNO3.Kinetic parameters were cal-culated after subtracting the linear component of the uptake rate calculated as the slope in the0.2–0.5concentration range. The results were obtained using the linearization of Linewe-aver–Burk.The linearizations of Hanes–Woolf and Woolf–Augustinsson–Hofstee were used for comparison(Segel 1976)and gave lower absolute values for V max and K m,but confirmed the differences between treatments.These kinetic parameters are not to be attributed to a single transporter,but refer to the overlapping activities of different transporters. NO3-reductaseNR was extracted from leaf tissues grinded in a mortar with liquid nitrogen.The extraction buffer(50mM potassium phosphate buffer,pH7.5,1mM ethylenediaminete-traacetate,1mM dithiothreitol(DTT),1l Mflavin adenine dinucleotide,10l M leupeptin and10l M chimostatin were then added to the tissue powder(0.04mL mg-1FW). The homogenate was centrifuged at4°C for30min at 12,500g.NR activity was measured immediately in the supernatant.The reaction mixture consisted of10mM phosphate buffer,pH7.5,supplemented with10mM KNO3and0.1mM NADH.The reaction was terminated after15min at28°C in the dark,by addition of an equal volume of sulfanilamide[1%(w/v)in1N HCl]and then naphthylethylene-diamine dihydrochloride[0.01%(w/v)] to the reaction mixture and the absorbance at540nm was measured.Determination of NO3-and Cd contentRoots were rinsed three times in distilled water and blotted with paper towels,frozen in liquid nitrogen and stored at -80°C until use.Leaves were collected and immediately frozen and stored.For NO3-content,300mg tissue was homogenized in ice cold deionised water(10mL g-1FW). The homogenate wasfiltered through four cheesecloth layers and transferred into2mL tubes,then centrifuged at 13,000g for15min.NO3-concentration was determined in200l L aliquots of the supernatants with the same pro-cedure described for NO3-uptake assay,except that for each sample a blank was prepared,omitting the salicylic acid from the H2SO4solution to subtract basal noise.For Cd content,as described in Zuchi et al.(2009),root and shoot tissues were oven-dried at80°C,ashed at 550°C,dissolved in1N HCl and analyzed by inductively coupled plasma atomic emission spectrometry(Varian, Torino,Italy).Isolation of plasma membranesPlasma-membrane vesicles were isolated from root samples as described in Tomasi et al.(2009)with slight modifications: 2g FW root tissue was homogenized with a mortar and pestle in4mL freshly prepared ice-cold extraction medium: 250mM sucrose,2mM MgSO4,2mM adenosine50-tri-phosphate,10%(v/v)glycerol,10mM glycerol-1-phos-phate,0.16%(w/v)BSA,2mM ethylene glycol tetraacetic acid,2mM DTT,5.7%(w/v)choline-iodide,1mM phen-ylmethylsulfonylfluoride,20l g mL-1chymostatin,25mM MES-1,3-bis[tris(hydroxymethyl)-methyloamino]propane (BTP)pH7.6and0.5g-1FW polyvinylpolypyrrolidone (PVPP).Homogenates werefiltered through four layers of cheesecloth and the suspensions were subjected to differen-tial centrifugation steps in an Eppendorf microcentrifuge at 2°C:12,700g for3min(pellets discarded),12,700g for 25min(pellets recovered).Microsomes,gently resuspendedPlantain400l L of homogenization medium(extraction medium without PVPP)were loaded onto a discontinuous sucrose gradient made by layering700l L sucrose solution (1.13g cm-3)on a300l L sucrose(1.17g cm-3)cushion and then centrifuged at12,700g for1h.The sucrose solutions were prepared in5mM MES-BTP pH7.4and contained all of the protectants present in the homogenization medium. Vesicles migrating to the1.13/1.17g cm-3interface were collected,diluted with1.8mL homogenization medium and centrifuged at14,000g for30min.Pellets were resuspended in a100l L medium containing250mM sucrose,10%(v/v) glycerol,1mM DTT,50l g mL-1chymostatin and2mM MES-BTP pH7.0,immediately frozen in liquid nitrogen and stored at-80°C.Measurement of PM H?-ATPase activityPM H?-ATPase hydrolytic activity was measured at38°C in a0.6mL reaction medium(50mM MES-BTP pH6.5, 5mM MgSO4,100mM KNO3,600l M Na2MoO4, 1.5mM NaN3,5mM ATP-BTP pH 6.5,0.01%(w/v) polyoxyethylene20cetyl ether(Brij58),with or without 100l M V2O5).The reaction was started by adding the membrane vesicles containing0.5l g of total protein;after 30min,the reaction was stopped and color developed as previously described by Santi et al.(1995).Inorganic phosphate was quantified spectrophotometrically at705nm as described by Forbush(1983).Protein content was determined as described by Bradford(1976),using BSA as standard,after solubilizing membrane vesicles with0.5M NaOH(Gogstad and Krutnes1982).The activity is expressed in l mol P mg protein h-1subtracting the quantity produced in the enzyme assay in presence of vanadate. Transcript levels analysisAt harvesting times,root samples were collected,imme-diately frozen in liquid nitrogen and conserved at-80°C until further processing.RNA extractions were performed using the Invisorb Spin Plant RNA kit(Stratec Molecular, Berlin,Germany).1l g of total RNA(checked for quality and quantity using a spectrophotometer,followed by electrophoresis in agarose gel)of each sample was retro transcribed using1pmol of Oligo d(T)23VN(Sigma Aldrich,Milano,Italy),15U Prime RNase Inhibitor(Ep-pendorf,Hamburg,Germany)and10U M-MulV RNase H-for1h at42°C(Finnzymes,Helsinki,Finland).After RNA digestion with1U RNase A(USB,Cleveland,OH, USA)for1h at37°C,transcript levels analyses were performed by adding0.1l L of the cDNA to FluoCycleTM sybr green(20l Lfinal volume;Euroclone,Pero,MI,Italy) in a DNA Engine Opticon Real-Time PCR Detection (Biorad,Hercules,CA,USA).Primers(T m=58°C)were the following:ZmNRT2.1 (AJ344451),GATCGACGATCACCTATACCTC and GTG CTCCGTTGACATGAG(PCR efficiency69%);ZmNRT 2.2(AY659965),CCTACCTTTACGTGTATGCCTTG and GATGTGCCAACGATATTCATC(PCR efficiency83%); ZmMHA1(U09989),CGAGAACAAGACGAGCTTCA and CAGTGGAGATGCTCGACAAA(PCR efficiency 75%);ZmMHA2(X85805),TCCGACTGTTGTTTGTCG AG and CACCGACTCCATCCTCATCT(PCR efficiency 71%);ZmMHA3(AJ441084),GCCAAGAGACGAGCTG AGAT and CACCGTGTAGTTCTGCTGGA(PCR effi-ciency84%);ZmMHA4(AJ539534),CGGTGATGTGAT TGGAGACA and CGGTGATGTGATTGGAGACA(PCR efficiency93%);ZmNR1(AF153448),CCAGCCGACTT GCCAGCGTAA and GCATGGCCTATGTTATCTGCTG CTC(PCR efficiency85%);ZmNADH:NR(M27821), GGTCTTTGGAGGTGGAGGTGCTG and CTCTGGCT GCGTATTCAAACTCTCGT(PCR efficiency85%); ZmST1.1(AF355602),AAGTGGAATCCATGCTTTGG and CTGAGCGGAGCTTCTGGAT(PCR efficiency74%). As housekeeping genes,ZmPolyU(polyubiquitin,S94466, GTACCCTCGCCGACTACAAC and ATGGTCTTGCCA GTCAAGGT,PCR efficiency83%),and ZmRPL17(ribo-somal protein L17,AF034948,AAAGTCTCGCCACTCC AATG and ACGTCCAAGCCTTTCACATC,PCR effi-ciency90%)were used.Triplicates were performed on three independent experiments;analyses of real-time result were performed using Opticon Monitor2software(Biorad)and R (version2.9.0;/)with the qPCR package(version1.1-8;http://www.dr-spiess.de/qpcR.html). Efficiencies of amplification were calculated following the authors’indications(Ritz and Spiess2008). Transmembrane topology predictionPredictions have been carried out at the PSIPRED Protein Structure Prediction Server of the University College of London(/psipred/)using MEM-SAT3with the default settings and the sequences retrieved from NCBI(/protein)for the following proteins:ZmNRT2.1(CAC87729.2),ZmNRT2.2 (AY659965.1),ZmNRT1.2(AAY40798.1)and ZmNAR2.1 (AAY40796.1).Statistical analysisComputation of the graphical representation and statistical validation(ANOVA and Student’s t test;P\0.05)were performed on data belonging to each time point(not between different time points)using SigmaPlot11.0(Sy-stat software,Point Richmond,CA,USA).Transcript levels data were illustrated considering the differences inPlantathe PCR efficiency of amplification and using the mean transcript level of the housekeeping genes ZmPolyU and ZmRPL17in roots of control non-induced plants at time zero as reference.ResultsThe data presented have been obtained using maize seed-lings exposed to0,1or10l M Cd during a24-h induction for NO3-uptake(1mM NO3-).In our experimental conditions Cd exposure did not produce any apparent symptoms of toxicity.As expected(Nocito et al.2002),a typical detoxification response was activated by all Cd-treated plants,as evidenced by decreased glutathione pools, increased non-protein thiols concentrations and higher SO42-uptake capacity(Online Resource1:Suppl.Figs. S1–S3).At the end of the exposure period,Cd concentra-tion was higher in the roots as compared to the shoots(184 vs.52l g g-1DW and470vs.102l g g-1DW,in plants treated with1and10l M Cd,respectively).Cd effect on the induction of high-affinity NO3-transporters’activity,NO3-accumulationand reductionInduced plants,when treated with1mM NO3-,gradually developed a greater net high-affinity NO3-uptake rate, measured at0.2mM NO3-,with a peak after12h of treatment and a subsequent de-induction phase(Pinton et al.1999;Santi et al.2003);the increase in net high-affinity NO3-uptake was not observed in plants not sup-plied with NO3-(control non-induced),which maintained their basal uptake rate all along the experimental period (Fig.1).On the other hand,the presence of Cd in the nutrient solution strongly affected the induction of root high-affinity NO3-uptake:only a slight induction of the high-affinity net NO3-uptake rate was observed in plants supplied with both1mM NO3-and1l M Cd(indu-ced?1l M Cd);the presence of10l M Cd strongly impaired induction of the high-affinity NO3-uptake (Fig.1).Conversely,no significant alteration was observed in the constitutive net high-affinity NO3-uptake rate(non-induced?1or10l M Cd;Fig.1).NO3-supply also produced a gradual increase in the NR activity of the roots,which was slightly,although signifi-cantly,less pronounced in the presence of Cd(Fig.2a).In induced Cd-treated plants,root NO3-content was similar to that of the control-induced plants and started to decline after 8h of treatment with the highest Cd concentration(10l M Cd)(Fig.2b).NO3-accumulation in the roots was faster and more pronounced than in the shoots:NO3-content was about twofold higher in the roots than in the shoot after 24h(Fig.2b).NO3-content after24h exposure to0,1or 10l M Cd was100,94and71%,respectively,in the root, and100,84and63%,respectively,in the shoot.The activity of the PM H?-ATPase,which is known to increase in maize in response to NO3-supply(Santi et al. 1995)was measured in vesicles isolated from roots.The time course of ATP hydrolysis rate in the different treat-ments was similar to that described for NO3-uptake rate. Figure3shows an increase in the ATP hydrolysis rate in the vesicles isolated from roots of control-induced plants, with a peak after12h of treatment which matches the time of maximum NO3-uptake rate measured at0.2mM(see Fig.1).On the other hand,induced Cd-treated plants only showed a slight increase in their ATP hydrolysis rate.Cd effect on NO3-uptake kineticsNO3-uptake rates as a function of external NO3-con-centration,in the range of0.125–10mM,were measured (Figs.4,5;Table1).The kinetic parameters(V max and K m) for the high-affinity transport,which was considered to be operating below0.5mM NO3-(Hogh-Jensen et al.1997; Siddiqi et al.1990)were calculated after subtraction of the linear component of the uptake rate,estimated as the slope in the0.2–0.5mM concentration range(Table1).In afirst set of measurements(long exposure;Table1; Fig.4),root NO3-uptake rates were comparedamong Fig.1High-affinity net NO3-uptake rate in roots of maize seedlings supplied with1mM NO3-and0(control induced),1or10l M Cd in the nutrient solution.Control non-induced plants were treated in nutrient solution without NO3-uptake was measured spectrophotometrically as the depletion from a solution containing 0.2mM NO3-.Closed circles,control induced;open circles, induced?1l M Cd;closed triangles,induced?10l M Cd;open triangles,control non-induced;closed squares,non-induced?1l M Cd;open squares,non-induced?10l M Cd.Data are mean±SD, letters refer to statistically significant differences within each time point among independent experiments,underlined letters refer to overlapping data not significantly different among treatments (ANOVA,n=3,P\0.05)Plantadifferent treatments:(1)plants supplied for 12h with a complete nutrient solution containing no NO 3-(non-induced),(2)plants supplied for 12h with a complete nutrient solution containing no NO 3-and with the addition of Cd (non-induced ?1l M Cd),(3)plants fed for 12h with a complete solution containing 1mM NO 3-(induced),and (4)plants treated for 12h with both NO 3-and Cd (induced ?1l M Cd).Induced plants,compared to non-induced ones,showed increased NO 3-uptake rates both in the high-and low-affinity concentration ranges (Fig.4a,b,respectively).The kinetic parameters calculated for the high-affinity NO 3-uptake (Table 1)showed a decrease in the K m value of induced plants and an increase in the V max .On the other hand,non-induced plants did not show any difference when compared to non-induced ones treated with Cd (Fig.4a).Finally,induced plants treated with Cd showed an intermediate uptake rate in the low-affinity concentration range (Fig.4b),while in thehigh-affinity concentration range the uptake rate remained similar to the constitutive uptake rate of non-induced plants (Fig.4a).In these latter plants,Cd effect was particularly evident on the K m value,while the V max value was between that of non-induced and control induced plants (Table 1).A second set of measurements (short exposure,Fig.5)was performed using plants either non-induced or induced with 1mM NO 3-for 12h without any Cd addition,and then exposed to 0or 1l M Cd during the 10-min NO 3-uptake assay.Again,when NO 3-uptake was measured in the high-affinity concentration range,the short exposure to Cd did not cause any significant decrease in the constitutive uptake rates of non-induced plants,but strongly depressed uptake rates of induced plants (Fig.5a).In Cd-treated plants,K m value was similar independent of NO 3-induction and comparable to that of plants not induced for NO 3-uptake and not exposed to Cd during the NO 3-uptake assay (control non-induced;Table 1).V max value was significantly decreased by the short Cd treat-ment in plants induced for NO 3-uptake (control induced).Different to the prolonged exposure described above,the short exposure to Cd did not affect,either in non-induced or induced plants,the NO 3-uptake rates measured in the low-affinity concentration range,which remained as high as that of plants that were never exposed to the heavy metal (Fig.5b).Cd effect on transcript levels of genes related to NO 3-acquisition in root tissuesThe transcript amount of the genes ZmNRT2.1and ZmNRT2.2,which encode two putative high-affinity trans-porters,was analysed.mRNA level of ZmNRT2.1reachedaFig.2Activity of NR (a )and NO 3-content (b )in shoots (above )and roots (below ),measured spectrophotometrically after extraction from root tissues of maize seedlings supplied with 1mM NO 3-and 0(control induced),1or 10l M Cd in the nutrient solution.a Closed circles ,control induced;open circles ,induced ?1l M Cd;closed triangles ,induced ?10l M Cd.b White bars ,control plants at time zero;black bars ,induced;light grey bars ,induced ?1l M Cd;dark grey bars ,induced ?10l M Cd.Data are mean ±SD,letters refer to statistically significant differences within each time point among independent experiments (ANOVA,n =3,P \0.05)Fig.3Vanadate-sensitive phospho-hydrolysing activity of the PM H ?-ATPase in vesicles isolated from roots of maize seedlings supplied with 1mM NO 3-and 0(control induced),1or 10l M Cd in the nutrient solution.Phospho-hydrolysing activity was measured spectrophotometrically on root microsomal fractions.Closed circles ,control induced;open circles ,induced ?1l M Cd;closed triangles ,induced ?10l M Cd.Data are mean ±SD,letters refer to statistically significant differences within each time point among independent experiments (ANOVA,n =3,P \0.05)Plantamaximum between 8and 12h from the beginning of NO 3-supply in roots of control-induced plants (Fig.6a).On the other hand,in induced Cd-treated plants no significant change in ZmNRT2.1transcript accumulation was measured during the experimental time span (Fig.6a).Consequently,mRNA amount after 12h from starting NO 3-supply appeared to be significantly higher in control-induced plants than in plants treated with 1l M Cd.After 24h ZmNRT2.1transcripts in induced Cd-treated plants showed values thatwere not significantly different from those of control-induced plants,where feedback regulation started to occur.The time course of ZmNRT2.2mRNA accumulation showed a peak after 4h in control-induced plants with a 2.9-fold increase when compared to 0h (Fig.6b).Cd-treated plants showed a clear increase in transcript level only after 24h with values 5.6times higher than at 0h.ZmMHA3and ZmMHA4encode two putative PM H ?-ATPases.The accumulation of ZmMHA3transcripts in control-induced plants peaked after 4h of treatment and later declined (Fig.6c).In Cd-treated plants the increase in ZmMHA3mRNA was lower and delayed reaching a peak after 8h (Fig.6c).Transcript amount of ZmMHA4showed,in control-induced plants,a trend similar to that of ZmMHA3,withaFig.4Net NO 3-uptake kinetics in roots of maize seedlings after prolonged exposure to Cd (12h),measured in the high-affinity concentration range (0.125–0.5mM NO 3-,a )and in the low-affinity concentration range (0.5–10mM NO 3-,b ).Control induced plants were supplied with 1mM NO 3-for 12h before the uptake assay;control non-induced plants were treated for 12h in nutrient solution without NO 3-.For Cd-treated plants 1l M Cd was added to the nutrient solutions for 12h.Closed circles ,control induced;open circles ,induced ?1l M Cd;open triangles ,control non-induced;closed squares ,non-induced ?1l M NO 3-uptake was measured spectrophotometrically as depletion from solutions con-taining different concentrations of NO 3-.Data are mean ±SD,letters refer to statistically significant differences within each time point among independent experiments;underlined letters refer to overlap-ping data not significantly different among treatments (ANOVA,n =4,P \0.05)Fig.5Net NO 3-uptake kinetics in roots of maize seedlings after short exposure to Cd (10min),measured in the high-affinity concentration range (0.125–0.5mM NO 3-,a )and in the low-affinity concentration range (0.5–10mM NO 3-,b ).Control induced plants were supplied with 1mM NO 3-for 12h before the uptake assay;control non-induced plants were treated for 12h in nutrient solution without NO 3-.For Cd-treated plants 1l M Cd was added to the assay medium (10min).Labels,measurements and statistical analyses as described in Fig.4Planta。