基因工程酶学基础
- 格式:ppt
- 大小:1.05 MB
- 文档页数:71



第三章基因工程的酶学基础第三章基因工程的酶学基础第一节限制性核酸内切酶(RestrictionEndonuclease)一、限制性核酸内切酶的发现早在五十年代初,两个研究小组几乎同时发现了两种不同来源的l噬菌体(lK和lB)能高频感染它们各自的大肠杆菌宿主细胞(K株和B株),但当它们分别与其它宿主菌交叉混合培养时,则感染频率普遍下降数千倍。
一旦lK噬菌体在B 株中感染成功,由B株繁殖出的lK后代在第二轮接种中便能象lB一样高频感染B株,但却不再有效地感染它原来的宿主K株。
这种现象称为宿主细胞的限制和修饰作用,它广泛存在于原核细菌中。
1960s,Linn和Arber在研究细胞限制性和修饰现象时在大肠杆菌中发现了限制性内切酶,人们才搞清了细菌限制和修饰作用的分子机制。
大肠杆菌K株和B株都含有各自不同的限制-修饰系统,它们均有三个连续的基因位点控制,其中hsdR编码限制性核酸内切酶,它能识别DNA分子上的特定位点并将双链DNA切断;hsdM的编码产物是DNA甲基化酶,催化DNA分子特定位点上的碱基甲基化反应;而hsdS表达产物的功能则是协助上述两种酶识别特殊的作用位点。
lK和lB长期分别寄生在大肠杆菌的K株和B株中,宿主细胞内甲基化酶已将其染色体DNA 和噬菌体DNA特异性保护,封闭了自身所产生的限制性核酸内切酶的识别位点。
当外来DNA入侵时,便遭到宿主限制性内切酶的特异性降解,由于这种降解作用的不完全性,总有极少数入侵的DNA分子幸免于难,它们得以在宿主细胞内复制,并在复制过程中被宿主的甲基化酶修饰。
此后,入侵噬菌体的子代便能高频感染同一宿主菌,但丧失了在其原来宿主细胞中的存活力,因它们在接受了新宿主菌甲基化修饰的同时,也丧失了原宿主菌甲基化修饰的标记。
大肠杆菌C株不能产生限制性内切酶,因而其它来源的l噬菌体可以感染C株,而在C株中繁殖的l噬菌体则在K 株和B株中受到严格的限制作用,细菌正是利用限制修饰系统来区分自身DNA与外源DNA的。





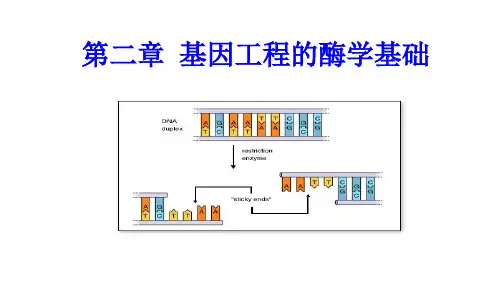
第二章基因工程的酶学基础内容一、概述二、限制性内切核酸酶三、DNA连接酶四、其他工具酶一、概述工具酶:在生物技术中常用的各种工具酶系指能用于DNA和RNA分子的切割、连接、聚合、反转录等有关的各种酶系统称为工具酶。
工具酶名称主要功能限制性内切核酸酶在DNA分子内部的特异性的restriction endonucleases 碱基序列内部进行切割DNA连接酶将两条以上的线性DNA分子或片段DNA ligase 催化形成磷酸二酯键连接成一个整体DNA聚合酶I通过向3’端逐一增加核苷酸以填补双链DNA分子上的单链DNA polymerase I 裂口,即5’→3’DNA聚合酶活性与3’→5’及5’→3’外切酶活性多核苷酸激酶催化将把一个磷酸分子加到多核苷酸链的DNA polymerase kinease 5’-OH末端上(接下表格)工具酶名称主要功能反转录酶以RNA分子为模板合成互补的cDNA链reverse transcriptaseDNA末端转移酶将同聚物尾巴加到线性双链或单链DNA分子的3’-OH DNA terminal transferase 末端或DNA的3’-末端标记dNTP碱性磷酸酶去除DNA,RNA,dNTP的5’磷酸基团BAP orCIAP核酸外切酶III 降解DNA3’-OH末端的核苷酸残基exonuclease III降解酶S1 降解单链DNA或RNA,产生带5’磷酸的单核苷酸或nuclease S1寡聚核苷酸,同时也可切割双链核酸分子的单链工具酶名称主要功能核酸酶Bal 31 降解双链DNA,RNA的5’及3’末端,nuclease Bal31Taq DNA聚合酶能在高温(72℃)下的单链DNA为模板,Taq DNA polymerase从5’→3’方向合成新生的互补链核糖核酸酶专一性降解RNARNase脱氧核糖核酸酶内切核酸酶,水解单链或双链DNA DNase二、限制性内切核酸酶1、限制性核酸内切酶的分类2、限制性核酸内切酶的命名原则3、限制性核酸内切酶的基本特征4、影响限制性内切酶的活性因素5、限制性核酸内切酶的应用1、限制性核酸内切酶的分类限制性核酸内切酶主要分成三大类:I类:能识别专一的核苷酸顺序,并在识别点附近的一些核苷酸上切割双链,但切割序列没有专一性,是随机的。