分子人类学的人种分类
- 格式:doc
- 大小:28.50 KB
- 文档页数:10

三大人种的划分标准
人种的划分是一个复杂和敏感的话题,几种主要的人种分类方法如下:
一、人类遗传学分类法:
这种分类方法主要根据地理和人类进化的角度进行划分。
在这个分类中,人类被分为三个主要的群体:非洲人种、亚洲人种和欧洲人种。
这种分类方法主要侧重于遗传差异和人类进化,但它并不能完全准确地揭示人类的多样性和复杂性。
二、人种学分类法:
该分类方法主要侧重于人的生理特征,如皮肤颜色、眼睛形状、头发类型等。
根据这种分类方法,一般将人类分为三个主要人种:蒙古人种(亚洲人种)、高加索人种(白人种)和尼格罗人种(非洲人种)。
这种分类方法也受到了批评,因为人的生理特征并不是只由一个基因或一组基因决定的,且人群之间的边界并不明确。
三、文化人类学分类法:
这种分类方法主要基于文化和社会特征,包括语言、宗教、风俗习惯等。
根据这种分类方法,人类被分为多个文化群体,如西方文化、东方文化、非洲文化等。
但需要注意的是,文化是一个动态变化的概念,并且一个地区内的文化群体可能存在着巨大的多样性和细微差异。

世界人种分类——胡顿分类法
人类学家胡顿(E·A·Hooton)提出人种分类法(1946)
1、蒙古人种(Mongoloid race)
亦称亚美人种或黄色人种,分亚洲和美洲两大支系,亚洲支系包括北、东、南三个分支以及各种过渡类型和混和类型;美洲支系包括因纽特人和印第安人;约占世界人口41%。
2、赤道人种(Equatorial race)
亦称尼格罗-澳大利亚人种,俗称黑色人种。
分尼格罗和澳大利亚两个支系以及各种过渡类型和中间类型,包括热带非洲、大洋州、南亚和东南亚许多民族。
18世纪后,美洲亦有分布,约占世界人口16%。
3、欧罗巴人种(Europeoid race)
亦称欧亚人种或高加索(Caucasoid)人种,俗称白色人种。
分南北两个支系,南支包括印度、地中海和巴尔干高加索分支类型;北支包括大西洋、波罗的海和白海波罗的海分支类型以及各种过渡类型和中间类型。
过去主要分布在欧洲、北非、西亚和南亚,包括印欧和闪含两大语系的各个民族,16世纪后,美洲、大洋州、南非亦有分布,约占世界人口43%。

人种分类算法
人种分类算法是指根据人类体质特征的不同,将全球人类划分为不同的人种。
以下是几种常见的人种分类算法:
1.三分法:将全球人类划分为黄种人、白种人和黑种人三大类。
2.四分法:将全球人类划分为黄种人、白种人、黑种人和棕种
人四大类。
3.五分法:将全球人类划分为黄种人、白种人、黑种人、棕种
人和红种人五大类。
4.分子人类学:根据人类Y染色体单倍型类群进行划分,将全
球人类划分为K、N、R、C、O等类型。
5.遗传学和进化生物学:根据人类的遗传基因和进化历程,将
全球人类划分为非洲系、欧洲系、亚洲系和美洲系等类型。
需要注意的是,人种分类算法并不是绝对的,不同的分类算法有其优缺点,且人种的划分也受到历史、文化和社会因素的影响。
因此,在研究和应用中应该根据具体情境选择合适的分类算法,并考虑多种因素的综合影响。

人种分类引言人种分类是人类学的一个重要研究领域,旨在理解和解释人类群体之间的差异和相似性。
人种分类的概念在历史上经历了多次变迁和争议,但它仍然是人类学、遗传学和社会科学的重要话题之一。
本文将探讨人种分类的定义、历史背景以及相关的科学研究,并讨论该领域的争议和影响。
一、人种分类的定义与历史发展1.1 人种分类的定义人种分类是指根据人类的遗传、生理、形态和文化特征将人类划分为不同群体的过程。
人种分类的目的是识别和比较人类群体之间的差异,以更好地了解人类进化、遗传和文化传承。
1.2 人种分类的历史背景人种分类的历史可以追溯到古代希腊和古代罗马时期,当时人们通过外貌特征和地理分布将人类分为不同的群体。
随着人类学和生物学的发展,人种分类的方法也不断演变,包括基于遗传、血型、颅骨形态、DNA、语言和文化等方面的分类方法。
二、人种分类的科学研究2.1 基于遗传的人种分类基于遗传的人种分类是最常用的分类方法之一。
研究人类基因组的发现使得科学家们能够比较不同人类群体之间的遗传差异。
这些研究通常使用DNA分析技术,例如单倍型分析和基因测序,以确定人类群体之间的遗传关系。
2.2 基于形态和生理的人种分类除了基因组研究,人种分类还可以基于形态和生理特征,如皮肤颜色、骨骼结构和血型等。
这些特征在不同人类群体之间存在一定程度的差异,因此可以辅助人种分类的研究。
2.3 基于文化的人种分类人种分类不仅仅局限于生物学特征,还包括了文化方面的研究。
人类不同群体的社会行为、语言和宗教信仰等因素也可以用来对人种进行分类。
然而,由于文化是多样化和动态的,这种分类方法也存在一定的争议。
三、人种分类的争议与影响3.1 争议:人种分类的模糊性人种分类的定义在不同文化和学科之间存在差异,因此人种分类本身就具有一定程度的模糊性。
一些科学家认为人种分类只是一种概念上的工具,无法准确反映人类的复杂遗传和文化多样性。
3.2 争议:人种分类的种族主义倾向尽管人种分类的研究被广泛用于科学研究和人类学领域,但它也被批评为可能存在种族主义倾向。

世界人种的大致分类和黄种人的迁徙世界人种的大致分类和黄种人的迁徙在生物学上,现在世界上的人种只是一个种,属于哺乳动物纲灵长目人科(Hominidae)人属(Homo) 智人种(Homosapiens)。
智人种根据共同遗传特征可以分为三个不同的智人种亚种。
1.蒙古利亚人种(Mongoloid):又称黄种人、亚美人种;该人种起源于中亚和东亚的干旱草原和半荒漠地区,其中包括居住于中亚东亚北亚的大陆人种、太平洋人种、北极圈的爱斯基摩人种美洲的印第安人种;该人种在三大人种中人数最多,现分布于中国、朝鲜、日本、西伯利亚、中南半岛、南北美洲;该人种的特征是皮肤黄色、头发黑而直、头型近似方形,面部宽平,鼻低、颧骨突出,眼狭细。
2.尼格罗人种(Negroid):又称黑色人种、赤道人种;起源于非洲,其中包括西非的森林尼格罗人种、中非班图人种、南非布须曼人种、澳大利亚人种等;该人种现分布于撒哈拉沙漠以南的非洲大陆、西印度群岛、美国、澳大利亚等地;其特征是皮肤黑棕色、毛发细短、卷曲、体毛特少、眼睛为棕色、头型长、鼻低而大、唇厚。
3.欧罗巴人种(Europeoid):又名白色人种或高加索人种(Cavcasoid);它起源于欧、亚、非相连接地区,包括北欧的波罗的海人种、东北欧的北海波罗的海人种、南欧的印度地中海人种、西亚的巴尔干高加索人种、中欧人种;该人种现分布于全部欧洲、亚洲的西伯利亚、西南亚、北非、印度、澳大利亚、南北美洲。
从语言来说,白种人分印欧、高加索、闪米特含米特、乌拉尔共4个语系。
其中,斯拉夫、日尔曼、拉丁、凯尔特、希腊阿尔巴尼亚、波罗的、吐火罗、雅利安、安纳托利亚、巴斯克属于印欧语系。
(爱尔兰威尔士苏格兰高卢属于凯尔特,立陶宛拉脱维亚亚美尼亚属于波罗的,波斯普什图库尔德语俾路支塔吉克奥塞梯属于西雅利安,巴基斯坦印度斯坦孟加拉吉普赛南尼泊尔僧伽罗马尔代夫属于东雅利安,赫梯吕底亚吕齐亚属于安纳托利亚)卡巴尔达、车臣印古什、达吉斯坦、格鲁吉亚属于高加索语系。

【原】世界四大人种是如何形成的?2019-12-14作者:沽酒居士西元2019、09、15;东元4716人类的四大人种,大约是在六到三万年前形成的。
一、人种进化二百五十万年前,最早的人类能人,即早期猿人,诞生于非洲东部,一百六十万年前,能人灭绝。
一万八十万年前,在非洲诞生了直立人,即晚期猿人,由于气候的变化,直立人中的一部分走出非洲,前往亚洲和欧洲,元谋猿人,北京猿人等等即是直立人。
二十万年前,亚洲和欧洲的直立人也灭绝了。
十几万前,在非洲又诞生了早期智人,他们由留在非洲的直立人进化而来,也是由于气候变化,他们的一部分也走出了非洲,但是,三万年前,来到亚洲和非洲的早期智人也灭绝了。
因此,云南元谋猿人属于170万年前的晚期能人到早期直立人的代表,70万年前到20万年前的北京猿人属于中期到晚期直立人。
这两支早已灭绝,只有3.4万年前到2.7万年前开始的晚期智人北京山顶洞人(面貌已经非常接近现代智人),勉强可以称为中国人的祖先。
所谓的三大人种,是19世纪的提法,并不准确,因为除三大人种欧罗巴人种(白种人)、尼格罗人种(黑种人)、蒙古人种(黄种人)之外还有棕种人,是西元20世纪末提出来四大人种的划分。
二、四大人种第一种四大人种的划分:欧罗巴人种(白种人)、尼格罗人种(黑种人)、蒙古人种(黄种人)、混血人种。
在学界将大洋洲的原始居民单独分类棕种人之前,第四大人种一直指的是南美洲欧洲大航海时代造成的混血人种。
第二种四大人种的划分:大约六万到五万年前,留在非洲的早期智人又进化为晚期智人,同样由于气候变化,晚期智人的一部分也走出了非洲,走向欧洲和亚洲。
关于人类的进化,气候和环境的变化是重要原因,这些变化导致人类一次次走出非洲。
另外,气候和环境的变化使人类在非洲不断进化,但是在其他大洲,这些变化则使得那里的人类被一批批灭绝,不断由非洲而来的一批批新品种所取代,这是因为只有非洲大陆的热带森林和草原,才能使得气候变化不至于灭绝那里的人类,相反能起到促进那里人类进化的作用,而其他大洲则不具备非洲那样得天独厚的条件。

分子人类学的人种分类(祖先人)东非现代智人A 系,(非洲棕种人)科伊桑人B 系,(非洲小黑人)俾格米人C 系,(亚洲棕种人)蒙古—澳洲人种,和D 系一起被称为老亚洲人D 系,(亚洲矮黑人)日本—西藏人种,安达曼人,西藏氐族,阿依努人E 系,(非洲黑人)尼格罗人,非洲最主要的黑人种群F 系,(泛白种人)广义欧亚人种J 系,(暗白种人)闪米特人,中东—地中海,阿拉伯人,犹太人I 系,(土白种人)先欧洲土著,分布南欧,北欧H 系,(棕黄种人)南印度,达罗毗涂人,古哈巴拉文明创建者K 系,(黄白种人)早期白种人NO 系,(黄种人)NO 系为新亚洲人,CD 系称老亚洲人N 系,(北亚黄种人)乌拉语支,芬兰人,极地人O 系,(东亚黄种人)01,沿海支:百越,马来,中南半岛02,东亚支:瓯骆族,骆越族,日本,朝鲜,泰国03,华北支:华夏族,炎黄东夷苗瑶P 系,(白种人)高加索人种Q 系,(黄白过渡种人)印第安人R 系,(显白种人)高加索人种R1a 印欧人:高加索东支,雅利安人,波斯,北印度人,东欧人)R1b 西欧人:高加索西支,凯尔特人,斯拉夫人,日耳曼人)这是第二种流传的分类法A 政1治1正确版一。
黑种人(尼格罗人种)1。
古黑人:A2。
老黑人:B3。
新黑人:E (E3b除外),部分D* , DE,少数R1b1二。
白种人(高加索人种)1。
古白人:G, H , I, J部分F,个别F2。
老白人:E3b,E3c3。
新白人:R, P, L,部分K三。
棕种人(澳大利亚人种)1 。
老棕人:C2,C4,C52。
新棕人:M ,部分K四。
黄种人(蒙古人种)1。
老黄人:C1, C3, D (部分D*除外)2。
新黄人:O, N , NO, Q,个别K,个别FB 言论1 自1 由1 版一。
黑种人(非洲—日本人种)1。
小黑人:B2。
矮黑人;D3。
大黑人:E,DE二。
白种人(欧洲人种)1。
暗白人:G, H , I, J, L, M,部分F,部分K 2。
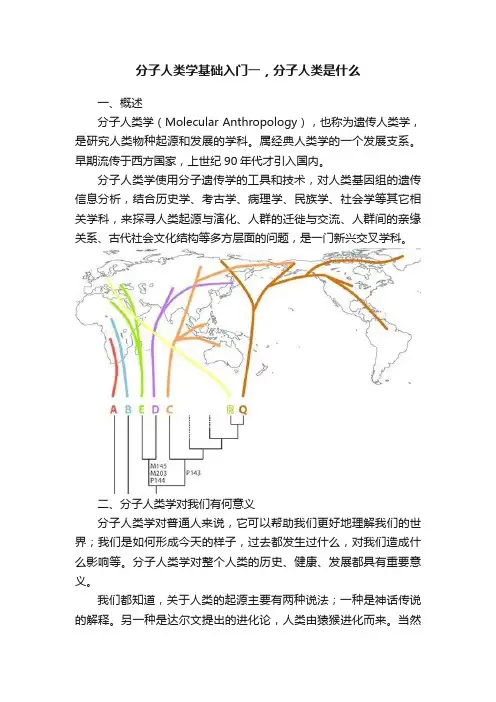
分子人类学基础入门一,分子人类是什么一、概述分子人类学(Molecular Anthropology),也称为遗传人类学,是研究人类物种起源和发展的学科。
属经典人类学的一个发展支系。
早期流传于西方国家,上世纪90年代才引入国内。
分子人类学使用分子遗传学的工具和技术,对人类基因组的遗传信息分析,结合历史学、考古学、病理学、民族学、社会学等其它相关学科,来探寻人类起源与演化、人群的迁徙与交流、人群间的亲缘关系、古代社会文化结构等多方层面的问题,是一门新兴交叉学科。
二、分子人类学对我们有何意义分子人类学对普通人来说,它可以帮助我们更好地理解我们的世界;我们是如何形成今天的样子,过去都发生过什么,对我们造成什么影响等。
分子人类学对整个人类的历史、健康、发展都具有重要意义。
我们都知道,关于人类的起源主要有两种说法;一种是神话传说的解释。
另一种是达尔文提出的进化论,人类由猿猴进化而来。
当然还有其它有趣的说法,比如人类是外星人所创造的。
因为猿猴与人类自身差异极大,普通人难以相信猿猴是如何一步一步形成今天的人类,所以大多数人宁愿相信后者。
分子人类学的研究,用DNA的证据证明了进化论在宏观层面的正确性,同时也填补了很多人类基因史未知的深刻事件。
如人类和其它所有物种一样“进化附带杂交”,在生存发展的同时也一直不断地和邻居群体或近邻物种发生杂交,在现代人类未走出非洲之前这些现象就已经存在了,走出非洲以后更是如此。
近些年的突破性研究发现,约7万年前现代人类走出非洲后,并没对原来在欧亚的早期智人(如尼安德特人、丹尼索瓦人等)进行完全取代,而是与其有过“联婚”杂交,有过长达4万多年的共存期。
这些杂交事件也发生的不只一次,而是多次。
并且今天所有亚欧人身体里都含有这些早期智人1%~3%的DNA片段。
由于现代智人和早期智人的生殖隔离,最终使得这些早期智人全部灭绝,能够生存至今的只有我们现代智人。
在过去分子人类学还没有兴起的年代,研究历史、探寻民族起源只能靠有限的史料和考古发现来大概推测,这是非常具有局限性的。

人种分类标准
人种分类是一个复杂的话题,不同的学科和文化背景可能有不同的分类标准。
以下是几种常见的人种分类标准:
1. 生物人类学分类:传统的生物人类学分类通常根据肤色、面部形态、头发类型和身体特征等生理特征将人种分为三大类:尼格罗人种、蒙古人种和高加索人种。
2. 色调分类:这种分类标准主要根据肤色进行区分,将人种分为黑人、棕色人种、黄色人种和白人。
3. 国际人种分类:国际人种分类(International Classification of Races)是国际统计界统一的人种分类标准,根据人口统计学特征将人群分为几类,如白种人、黑种人、黄种人、棕种人等。
4. 遗传学分类:基于遗传学特征进行分类,根据人群之间的遗传差异将人种分为不同的亚种。
需要注意的是,人种分类在科学界引发了很多争议,因为它在某种程度上暗示了人种的分离和等级差异,会造成歧视和偏见。
现在越来越多的学者和研究人员认为,人种是一个连续的谱系,没有明确的边界,应该强调个体的多样性而不是归类。

四类人群指的是哪四类
1、东亚人种(黄种人)
黄色人种主要分布于乌拉尔地区、东亚、北亚、东北亚、西伯利亚地区、南亚北部、东南亚及少数在美洲、大洋洲。
黄种人约占世界人口的37%。
2、高加索人种(白种人)
白色人种,又称欧罗巴人种、高加索人种,指具有特定体貌特征的一个人种,是体质人类学的概念。
白种人是世界上人数最多、分布最广的人种,占世界人口的百分之五十四左右。
3、非洲人种(黑种人)
非洲的黑皮肤居民分为很多不同的类型。
最常见的是苏丹尼格罗人和班图尼格罗人。
除此之外,还有黑白混血的埃塞俄比亚人和黑黄混血的马达加斯加人。
以及世界上最古老的种族——尼格利罗人和科伊桑人。
4、大洋洲人种(棕种人)
棕色人种是按照遗传和体质为人类分类所划分出的一个人种,分子人类学研究证实了他们的遗传基因的独特性。
一般来说,棕色人种包括澳大利亚土着人、美拉尼西亚人、巴布亚人和维达人(原始澳大利亚人)等支系。

德国人是欧罗巴人种,即白人。
德国人,有一亿至一亿五千万人口,分享共同的祖籍、文化和历史,通常以德语为母语。
根据分子人类学,德国人是由日耳曼人、斯拉夫人、部分犹太人和阿拉伯人融合而成的民族。
另有一部分德意志人,或中译德国人这个术语在使用中可能有多种含义。
在国籍上,德国人指的是拥有德国国籍的人在族群上,德国人可以包括以下几个特征:和德意志文化有关联以德语为第一语言德意志裔人:祖先来自德国或是曾经属于德国的地区者。
德国人是德国著名作家艾米尔路德维希创作的一部震撼人心的史诗。
它刻画了德国从公元800年到20世纪40年代的盛衰史。
这不是一部通常的编年史,而是通过对德国历史风云人物有血有肉的描绘,使读者似乎身历其境,重温千百年来的德国人历史。
德国人皮肤颜色较浅,多白皮肤,金发,棕发,红发等浅发,多蓝色或绿色眼睛。
波鸿鲁尔大学的心理学家告诉德国媒体:多数德国人觉得自己很漂亮。
中国人种基因图谱随着分子人类学数据的不断积累,父系Y染色体与母系mtDNA研究的一系列进展,使得中国人群的多样性结构逐渐明晰。
现有的Y染色体数据揭示,现代人出非洲后由东南亚经多次迁徙进入东亚。
在旧石器时代,现代人最初定居东亚或东南亚之后,紧接着不断北迁,这奠定了中国人遗传结构的基础。
通过了解人类基因的遗传成分,绘制中国人种基因图谱。
中国人种基因图谱:一对为性染色体,XY组合的为男性,XX组合的为女性。
Y染色体只在父亲与儿子传代,呈严格的父系遗传,研究Y染色体,可以发现人群在父系关系上的迁徙和发展。
母系mtDNA表现为母系遗传。
通过检测现代人mtDNA,能弄清各民族、各地人的母系血缘关系。
通过研究Y-DNA与mtDNA的重合型,可以揭开中国人的祖先来源之谜。
单倍群人类父系基因Y-DNA人类母系基因mtDNAY-DNA与mtDNA的重合型中国人起源华夏族的起源与形成单倍群在分子进化的研究中,单倍群或单倍型类群是一组类似的单倍型,它们有一个共同的单核苷酸多态性祖先。
因为单倍群由相似的单倍型组成, 所以可以从单倍型来预测单倍群.单核苷酸多态性试验被用来确认单倍型。
单倍群以字母来标记,并且以数字和一些字母来做补充,,例如O3a4。
Y染色体和线粒体单倍群有不同的单倍群标记方法。
单倍群用来标记数千年前的祖先来源。
在人类遗传学中, 最普遍被研究的单倍群是『人类Y染色体脱氧核糖核酸单倍群(Y-DNA单倍群)』和『人类线粒体脱氧核糖核酸单倍群(mtDNA单倍群)』,这两个都可以被用来定义遗传群体。
Y染色体脱氧核糖核酸单倍群仅仅被从父系线遗传,同时mtDNA仅仅被从母系线遗传。
人类父系基因Y-DNA在人类基因学里,人类Y染色体DNA单倍型类群通过Y染色体遗传变异特性进行人类学研究的一门科学,主要用于研究人类的“非洲起源论”及以后的种群分布的遗传学证据。
人类有23对46条染色体,其中22对44条为常染色体,另外一对为性染色体,XY组合的为男性,XX组合的为女性。
人种划分标准一、体质特征1.身高:不同人种的平均身高存在差异,如非洲人普遍较高,而亚洲人普遍较矮。
2.体重:同样,不同人种的平均体重也存在差异。
一般来说,非洲人的体重相对较重,而亚洲人的体重相对较轻。
3.肤色:人种的肤色大致可以分为白色、黄色、黑色、棕色等主要类型。
不同人种的肤色在色泽、深浅、分布上存在一定的差异。
4.眼睛:不同人种的眼睛形状、大小、颜色也存在一定的差异,如欧洲人的眼睛普遍较大,而亚洲人的眼睛则相对较小。
5.鼻子:不同人种的鼻子形状、大小也存在一定差异,如欧洲人的鼻子普遍较直挺,而亚洲人的鼻子则相对较扁平。
二、遗传特性1.基因组:不同人种的基因组存在一定的差异,这是由于人类进化的历史和地理分布所决定的。
2.单基因遗传特性:例如,人类的红绿色盲基因主要分布在某些特定的人群中,如欧洲人中男性红绿色盲的患病率较高。
3.多基因遗传特性:多基因遗传特性是指由多个基因共同决定的某种生理特征,如身高、体重等。
不同人种的多基因遗传特性也存在一定的差异。
三、文化特征1.语言:不同人种的语言体系存在很大的差异,这是由于历史、地理、社会等多种因素所决定的。
2.宗教信仰:不同人种的宗教信仰也存在很大的差异,如基督教主要分布在欧洲和美洲,伊斯兰教主要分布在亚洲和非洲等地区。
3.习俗和传统:不同人种的习俗和传统也存在很大的差异,如亚洲的儒家文化、印度的印度教文化、非洲的部族文化等。
四、地理分布1.人种地理分布具有一定的规律性,例如白种人主要分布在欧洲、北美洲、澳大利亚等地区,黑种人主要分布在非洲、美洲等地区,黄种人主要分布在亚洲等地区。
2.人种的地理分布也反映了人类进化的历史和人口迁移的历史,对于研究人类演化有着重要的意义。
五、社会结构1.社会结构是人种划分的一个重要方面,不同人种的社会结构存在很大的差异。
例如,欧洲和美国的白种人社会结构相对较为复杂,存在着明显的阶级和社会阶层分化,而非洲和亚洲的一些地区则存在着较为明显的部族和民族分化。
人种的划分人种的划分不是根据其语言、风俗和国籍的不同来划分,而是根据人类自然体质特征的不同,以及具有某种共同的遗传自然特征(属生物学范畴)为根据而划分的。
作为自然特征的主要指标是身长、肤色、发色、发型、眼色、头型、面型、鼻型等等。
据此,世界人种主要分为:白色人种、黄色人种、黑色人种以及其他人种等。
白色人种也称欧罗巴人种,是世界上人口最多的人种,约占全球人口总数的54%。
白种人多生活在较为寒冷的地区。
这个地区太阳斜射,光线和紫外线都较弱,所以白种人皮肤内黑色素含量低,为白红色到深棕色;毛发多而软,表现为波状金发或棕发;体毛稠密可防寒;眼睛发蓝,鼻子高而窄,鼻孔通道较长,可使冷空气较缓慢地进入肺部。
白种人的分布以欧洲为中心,还包括美洲、澳大利亚、东方的印度以及西亚的许多国家。
白种人又分为雅利安语族(日耳曼、拉丁、斯拉夫各族以及东方的印度族)、含族(分布于非洲北部以及东北部)和闪族(分布于西亚、阿拉伯及非洲东北部)三部分。
黄色人种也称蒙古利亚人种,是世界上第二大人种,约占全球人口总数的37%,其中一半是中国人。
黄色人种的皮肤是淡黄到棕色;体毛少,毛发直而黑;眼睛棕色,脸宽平,宽额;鼻子为中等高度。
黄种人的分布以亚洲为中心,扩散到周围各岛,并远至北美的印第安人。
黄种人又可分为蒙古人种(北蒙古人种以蒙古人为代表,中蒙古人种以中国人、朝鲜人为代表,南蒙古人种以缅甸人、泰国人为代表)、印度尼西亚人种、印第安人种等等。
欧洲的马札尔人、芬兰人也属黄色人种。
黑色人种也称尼格罗种人,是世界上第三大人种,约占世界人口总数的8.5%。
黑种人多生活在热带赤道地区。
这个地区太阳直射时间长,气温高,紫外线强,所以黑种人皮肤内黑色素含量高,以吸收太阳光中的紫外线保护皮肤,为黑色到黑棕色;毛发细而短,表现为卷曲形或波形黑发,头发空隙中充满空气,形成一个隔热层,起着一个保护头部的作用;体毛特少;眼睛棕色,鼻子低而宽,鼻孔通道短便于散热;头型长;唇厚。
地球人种的科学划分通常我们都简单地把人种分为白种人、黄种人、黑种人,不过这只是一种通俗的说法,并不准确。
因为肤色受外界影响很大,恰恰不是最重要的人种特征。
人类学的科学分类方法是按照语言划分,大概可分为:1、印欧语系民族(广义的雅利安人);2、闪米特语系民族(包括犹太人,阿拉伯人,古巴比伦人,古亚述人,腓尼基人等);3、含米特语系民族(包括古埃及人,北非柏柏尔人);4、汉藏语系民族;5、阿尔泰语系民族(包括蒙古,突厥,通古斯[女真]三大语族)等几大类。
其中大家最熟悉的当属雅利安民族,这一经纳粹德国反复强调的名词,并不是什么纯种民族,且听慢慢分解。
今天的欧洲除了保加利亚、匈牙利、芬兰、俄罗斯有属于阿尔泰语系游牧民族的后裔和混血外,其他几乎都是雅利安民族的后裔和混血。
雅利安人是活跃在东欧到中亚的草原地带的民族,公元前,伴随着他们的迁徙,他们征服了欧亚大陆几乎所有的土著民族,并把他们的血液和文化混入了对方的血管。
这实在是历史上的一个奇迹,况且他们当时还未跨上马背。
雅利安人主要是驯牛,他们随着牛车以慢速游动,直到与阿尔泰语系的蒙古利亚人相遇混血后才学会了驯马,但当他们到希腊时,已经威风凛凛地跨在游牧马背上了。
当然这是以后的事。
欧洲原先遍布着和埃及人非常相似的伊比利亚种族。
最先西进欧洲腹地的雅利安人是克尔特人,他们身材较高,皮肤白皙,习性剽悍,文化较高,很快把伊比利亚人征服并同化了。
他们不断地向大西洋方向推进,能够不被同化的土著民族极少,只有近代的葡萄牙人大部分还属于伊比利亚血统。
克尔特人就成了现在法国的先民。
雅利安人因为有游牧民族的游动性,即使再美好的土地,也挽留不住他们好动的脚步。
北欧的克尔特人一部分向西扩展,再渡海到了斯堪第那维亚半岛,成了挪威、瑞典、丹麦、冰岛人的先民。
在喀尔巴阡山和黑海以北,雅利安人和当地土著民族混合的结果,又产生了波兰、捷克、塞尔维亚人和部分俄罗斯人。
另一部分则进入意大利和希腊,公元1500年前左右,雅利安人进入希腊半岛,摧毁了爱琴文化,和当地土著民族混合后,创造了更高的文化。
从分子人类学角度分析汉族和华夏文明的形成目前分子人类学的进展配合考古学,人类比较学,以及古籍记载的大量证据,越来越清晰地勾画出汉族形成的过程。
以下做出个人的推测。
按照早期分子人类学的说法,最早到达中国大陆的是C系和D系人群,C系人群主要是沿海岸走的一群人。
后来在地球盛冰期结束后,在中国南方诞生的类马来人种O系分成几大支系几乎同时向北方大陆挺进,其扩散历史当在一万年以上。
随父系O系北上的母系主要对应的是mt-NR下的mt-F和mt-B.而这个时候在中亚诞生的黄种人Q系,向西南占据了中东两河流域和非洲的尼罗河流域,当其他人种支系还在石器时代时,他们最早进入了文明社会,发展出苏美尔文明和古埃及文明。
苏美尔文明(可以称做殷人系)发展出楔形文字和青铜器。
苏美尔人(殷人)迁移路线是从两河流域(对应苏美尔文明)于公元前2300年迁移到印度河流域(对应古印度哈拉巴文明),于公元前1700年再从印度河流域沿喜马拉雅山南麓进入中国中国境内的长流上游流域,在四川盆地建立三星堆文明,三星堆殷人可能又分成北线和沿长江而下东线两条线路进入中原,最后于公元前1300年于安阳建立殷商(即盘庚迁殷),公元前1046,周人攻破殷人首都朝歌后,殷人后裔大部从山东乘船迁移去了美洲,并在公元前1000年左右在中美洲发展出奥尔梅克文明,成为印第安人文明之母。
中外一些学者提出殷人东渡美洲论解释奥尔梅克文明突然出现以及奥尔梅克艺术凤格和中国殷商时代艺术惊人相像。
在文字方面,从两河流域的楔形文字,到哈拉巴文字符号,再到三星堆巴蜀符号,再到安阳殷墟甲骨文,是一个完整的迁移演化链条。
还有学者认为巴蜀符号与玛雅文同出一源,土家族为巴人后裔,保存有完整的伏羲推八卦、女娲造人、大洪水的传说。
印第安人最古老的文明因子同样是太昊少昊文化——太极、八卦、扶桑坛台、崇拜龙凤太阳,从这个角度讲应当有一支三星堆人是沿长江而下在武汉留下盘龙城青铜文化后又北上进入中原河南。
分子人类学的人种分类
(祖先人)东非现代智人
A系,(非洲棕种人)科伊桑人
B系,(非洲小黑人)俾格米人
C系,(亚洲棕种人)蒙古—澳洲人种,和D系一起被称为老亚洲人
D系,(亚洲矮黑人)日本—西藏人种,安达曼人,西藏氐族,阿依努人
E系,(非洲黑人)尼格罗人,非洲最主要的黑人种群
F系,(泛白种人)广义欧亚人种
J系,(暗白种人)闪米特人,中东—地中海,阿拉伯人,犹太人
I系,(土白种人)先欧洲土著,分布南欧,北欧
H系,(棕黄种人)南印度,达罗毗涂人,古哈巴拉文明创建者
K系,(黄白种人)早期白种人
NO系,(黄种人)NO系为新亚洲人,CD系称老亚洲人
N系,(北亚黄种人)乌拉语支,芬兰人,极地人
O系,(东亚黄种人)
O1,沿海支:百越,马来,中南半岛
O2,东亚支:瓯骆族,骆越族,日本,朝鲜,泰国
O3,华北支:华夏族,炎黄东夷苗瑶
P系,(白种人)高加索人种
Q系,(黄白过渡种人)印第安人
R系,(显白种人)高加索人种
R1a印欧人:高加索东支,雅利安人,波斯,北印度人,东欧人)
R1b西欧人:高加索西支,凯尔特人,斯拉夫人,日耳曼人)这是第二种流传的分类法
A政1治1正确版
一。
黑种人(尼格罗人种)
1。
古黑人:A
2。
老黑人:B
3。
新黑人:E(E3b除外),部分D*,DE,少数R1b1 二。
白种人(高加索人种)
1。
古白人:G,H,I,J,部分F,个别F
2。
老白人:E3b,E3c
3。
新白人:R,P,L,部分K
三。
棕种人(澳大利亚人种)
1。
老棕人:C2,C4,C5
2。
新棕人:M,部分K
四。
黄种人(蒙古人种)
1。
老黄人:C1,C3,D(部分D*除外)
2。
新黄人:O,N,NO,Q,个别K,个别F
B言论1自1由1版
一。
黑种人(非洲—日本人种)
1。
小黑人:B
2。
矮黑人;D
3。
大黑人:E,DE
二。
白种人(欧洲人种)
1。
暗白人:G,H,I,J,L,M,部分F,部分K 2。
纯白人:R,P,部分K,个别F
三。
棕种人(蒙古人种)
1。
黑棕人:A
2。
纯棕人:C
四。
黄种人(华夏人种)
1。
纯黄人:O,N,NO,个别K,个别F
2。
白黄人:Q,个别K,个别F 还有第三种分类法,以染色体单倍群标号为分类
Y染色体各大单倍群及中国常见单倍群速查
A和B——Y染色体最古老的分支,只分布在非洲。
C-M130:较早期到达东亚的人群,高频于阿尔泰语系的蒙古、满、哈萨克等族及澳大利亚土著等,汉人中通常5%–10%。
D-M174:较早期到达东亚的人群,在西藏、日本等地将近一半,在汉族和南方少数民族也有较低比例分布。
D1-M15:藏族及周边民族较高频、汉族及南方部分少数民族有较低比例分布。
D2-M55:仅分布于日本,占日本40%以上,绳文人的主要成分。
D3-P99:青藏高原东部(康区)、白马人及纳西族等高频。
E:非洲高频,南欧及中东有一定分布,中国极少。
F*-M89(G至T的祖群):中国零星分布,个别少数民族高频。
G:土耳其、高加索、哈萨克斯坦西部高频,中国零星分布。
H:印度次大陆,中国极少。
I:主要分布在欧洲,北欧和地中海巴尔干高频,中国极少。
J:阿拉伯、犹太人等高频,中国零星分布。
K*-M9(L至T的祖群):中国零星分布,个别少数民族高频。
L:西亚至南亚低频分布。
M:新几内亚土著和美拉尼西亚。
N-M231:较晚期到达东亚的人群。
阿尔泰语系、芬兰人等中高频分布,在中国广泛分布,汉人中通常10%以下,部分少数民族中较高频。
N1c-Tat:乌拉尔语系的标志性单倍群,中国少量分布。
O-M175:较晚期到达东亚的人群,广泛高频分布于东亚,占汉族75%以上。
O1a-M119:中国东南沿海、壮侗族群、台湾原住民分布较集中,东南亚岛屿也有广泛分布。
O2-M268:汉族中5%以上。
O2a-M95:华南、南方少数民族、中南半岛及印度Munda人群分布较多。
O2b-M176:最主要集中于朝鲜半岛、朝鲜族和日本弥生人,越南和汉族也有少量分布。
O3-M122:中国最常见的单倍群,遍及整个东亚和东南亚,占汉族50%–60%左右。
O3a3b-M7:苗瑶族群特征类型,通常占汉族5%以下。
O3a3c-M134:汉族30%左右,广泛分布于东亚、东南亚。
O3a3c1-M117:汉族和藏缅语族的特征类型,汉族15%以上。
O3a4-002611:汉族常见类型,来源尚未知,汉族10%以上。
P*-M45(Q和R的祖群):很少见。
Q-M242:印第安人的绝大部分,北亚一些群体高频,汉族2%左右。
R-M207:印欧语系的主要群体,高频分布于欧洲至中亚、南亚,汉族2%左右,中国部分少数民族较高。
S:新几内亚土著和美拉尼西亚。
T:印度、中东、地中海、东非等地较低频分布。
基本得出各人的分类规律,首先是按照外观体貌特征,肤色来分类,如黑种人,棕种人,白种人,黄种人。
第二,按照肤色人种来分类,再按照发展顺序时期的类型,如古,老,新非洲人。
古,老,新棕种人,古,老,新白种人,老黄种人,新黄种人。
第三,按照Y-DNA染色体单倍群标号来分,也是人种先后突变出新的支系来分。
如A、B、C、D、E、F、G。
估计就是这样分类该很完整了,。