最新6核糖体与核酶汇总
- 格式:doc
- 大小:27.00 KB
- 文档页数:8


1. 核糖体(riboso me)核糖体是细胞内一种核糖核蛋白颗粒(ribonucleoprotein particle), 其惟一功能是按照mRNA的指令将氨基酸合成蛋白质多肽链,所以核糖体是细胞内蛋白质合成的分子机器。
按核糖体存在的部位可分为三种类型:细胞质核糖体、线粒体核糖体、叶绿体核糖体。
按存在的生物类型可分为两种类型:真核生物核糖体和原核生物核糖体。
原核细胞的核糖体较小, 沉降系数为70S,相对分子质量为2.5x103 kDa,由50S和30S两个亚基组成; 而真核细胞的核糖体体积较大, 沉降系数是80S,相对分子质量为3.9~4.5x103 kDa, 由60S和40S两个亚基组成。
在真核细胞中, 核糖体进行蛋白质合成时,既可以游离在细胞质中, 称为游离核糖体, 也可以附着在内质网的表面, 称为膜旁核糖体或附着核糖体。
真核细胞含有较多的核糖体, 每个细胞平均有106~107个, 而原核细胞中核糖体较少每个细胞平均只有15×102~18×103个。
典型的原核生物大肠杆菌核糖体是由50S大亚基和30S小亚基组成的。
在完整的核糖体中,rRNA约占2/3, 蛋白质约为1/3。
50S大亚基含有34种不同的蛋白质和两种RNA分子,相对分子质量大的rRNA的沉降系数为23S,相对分子质量小的rRNA为5S。
30S小亚基含有21种蛋白质和一个16S的rRNA分子。
真核细胞核糖体的沉降系数为80S,大亚基为60S,小亚基为40S。
在大亚基中,有大约49种蛋白质,另外有三种rRNA∶28S rRNA、5S rRNA 和5.8S rRNA。
小亚基含有大约33种蛋白质,一种18S的rRNA。
2. 基因扩增(gene a mp li fica tion)细胞内选择性复制DNA, 产生大量的拷贝。
如两栖类卵母细胞在发育的早期,rRNA基因的数量扩增到1000多倍。
基因扩增是通过形成几千个核进行的,每个核里含有几百拷贝的编码28S、18S和5.8S的rRNA基因,最后卵母细胞中的这些rRNA基因的拷贝数几乎达到50万个,而在相同生物的其它类型细胞中,这些rRNA基因的拷贝数只有几百个。

细胞生物学目录第一章绪论第二章细胞生物的研究方法和技术第三章质膜的跨膜运输第四章细胞与环境的相互作用第五章细胞通讯第六章核糖体和核酶第七章线粒体和过氧化物酶体第八章叶绿体和光合作用第九章内质网,蛋白质分选,膜运输第十章细胞骨架,细胞运动第十一章细胞核和染色体第十二章细胞周期和细胞分裂第十三章胚胎发育和细胞分化第十四章细胞衰老和死亡第一章绪论1.原生质体:被质膜包裹在细胞内的所有的生活物质,包括细胞核和细胞质细胞质:细胞内除核以外的原生质,即细胞中细胞核以外和细胞膜以内的原生质部分原生质体:除去细胞壁的细胞2.结构域:生物大分子中具有特异结构和独立功能的区域3.装配模型:模板组装,酶效应组装,自组装4.五级装配:第一级,小分子有机物的形成第二级,小分子有机物组装成生物大分子第三级,由生物大分子进一步组装成细胞的高级结构第四级,由生物大分子组装成具有空间结构和生物功能的细胞器第五级,由各种细胞器组装成完整细胞6.支原体:目前已知的最小的细胞第二章细胞生物的研究方法和技术1.显微镜技术:光镜标本制备技术、2.光镜标本制备技术步骤:样品固定、包埋与切片、染色3.电子显微镜种类:透射电子显微镜,扫描电镜,金属投影,冷冻断裂和冷冻石刻电镜,复染技术,扫描隧道显微镜4.细胞化学技术:酶细胞化学技术,免疫细胞化学技术,放射自显影5.细胞分选技术:流式细胞术6.分离技术:离心技术,层析技术,电泳技术第三章质膜的跨膜运输1.细胞功能:外界与通透性障碍,组织和功能定位,运输作用,细胞间通讯,信号检测2.膜化学组成:膜脂,膜糖,膜蛋白3.膜脂的三个种类:磷脂,糖脂,胆固醇4.脂质体用途:用作生物膜的研究模型,作为生物大分子与药物的运载体5.膜糖功能:细胞与环境的相互作用,接触抑制,信号转导,蛋白质分选,保护作用。
6.膜蛋白类型:整合蛋白,外周蛋白,脂锚定蛋白7.膜蛋白功能:运输蛋白,酶,连接蛋白,受体(信号接受和传递)8.不对称性的研究方法:冰冻断裂复型,冰冻蚀刻9.膜流动性研究方法:质膜融合,淋巴细胞的成斑成帽效应,荧光漂白恢复技术10.膜流动性的重要性:酶活性,信号转导,物质运输,能量转换,细胞周期11.影响膜脂流动性的因素:脂肪酸链,胆固醇,卵磷脂/鞘磷脂比值12.影响膜蛋白流动的因素:整合蛋白,膜骨架,细胞外基因,相邻细胞,细胞外配体、抗体、药物大分子13.膜骨架的主要蛋白:血影蛋白,肌动蛋白和原肌球蛋白,带4.1蛋白,锚定蛋白14.转运蛋白质包括:载体蛋白,通道蛋白15.协同运输的方向:同向协同,反向协同第四章细胞与环境的相互作用1.细胞表面结构:细胞外被、膜骨架、胞质溶胶2.细胞外被功能:连接,细胞保护,屏障3.糖萼:由细胞表面的碳水化合物形成的质膜保护层,又称为多糖包被。

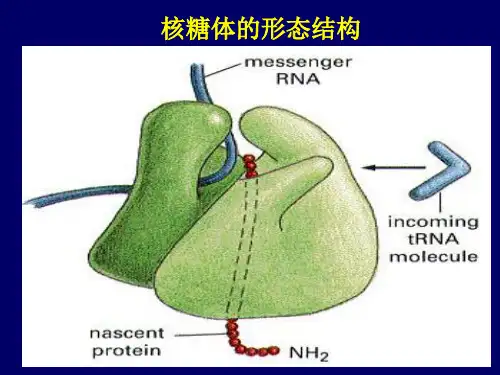
一、核糖体的形态结构⏹ 核糖体唯一的功能是按照m R N A 的指令将氨基酸合成蛋白质多肽链。
使细胞内蛋白质合成的分子机器,是细胞内数量最多的细胞器。
1、 核糖体的类型和化学组成⏹大小两个亚基都是由核糖体R N A 和核糖体蛋白组 成的。
(M g 2+的浓度)⏹ 原核生物(大肠杆菌)的核糖体:⏹ 大亚基50S :33种蛋白质;23S r R N A ,5S r R N A ⏹ 小亚基30S :21种16S rRN A (小亚基 主要由16S r R N A 决定)⏹ 真核细胞核糖体: ⏹ 大亚基60S :49种蛋白质;28S r R N A ,5 S r R N A , 5.8 S r R N A ⏹ 小亚基40S :33种蛋白质;18S r R N A 二、核糖体的生物发生⏹ 1、 核糖体r R N A 基因的转录与加工⏹ 真核生物核糖体由18S 、5.8S 、28S r R N A 和5S r R N A 基因 ⏹ 真核生物有四种r R N A 基因,⏹ 真核生物前r R N A 的修饰:两个特征1. 2以及修饰的意义。
⏹真题再现:03选择前体r R N A 甲基化的重要作用是: A .保证最后的r R N A 能够装配成正确的三级结构B .防止前体r R N A 被加工(x 对加工起引导作用) C .防止成熟r R N A 部分被降解。
二、核糖体的生物发生 ---真核生物的核糖体生物发生 ⏹ 2 5S r R N A 基因的转录与加工 ⏹ 由R N A 聚合酶3转录,使用的是内部启动子。
⏹ 学习重点⏹ 1.关于核糖体的形态结构, 主要学习掌握真核细胞和原核细胞核糖体的化学组成、细菌核糖体的结构模型。
⏹ 2. 核糖体的生物发生是本章的重点内容之一⏹ 3.核糖体的蛋白质合成作用,反义R N A 与核酶⏹ 本章考题近年来主要以小题为主。
第六章 核糖体与核酶2.1原核生物核糖体重组实验:⏹ (1)30S 亚基的蛋白质只和16SR N A 结合,50S亚基质只和23S r R N A 结合⏹ (2)不同种之间提取的30S 亚基的r R N A 和蛋白质可以装 配成有功能的30S 亚基,即不存在种间的差异⏹ (3)原核生物核糖体与真核生物核糖体的亚基彼此不 同,由二者的r R N A 和蛋白质装配成的核糖体没有活性 ⏹ (4)大肠杆菌的核糖体与玉米叶绿素核糖体亚基重组后 具有功能 ⏹(5)线粒体的核糖体亚基同原核生物核糖体亚基之间形 成的杂合核糖体没有功能 真核生物核糖体重组⏹ 边合成边装配,18S r R N A ,5.8r R N A ,28S r R N A在核仁中,边转录边装配,5S r R N A 在细胞核中转录后在运送到核仁里参与装配三、核糖体的功能—蛋白质的合成⏹ 1、 核糖体的功能位点 ⏹ ●A 位点(受位):接收氨酰t R N A 的部位 ⏹ ●P 位点(供位):肽酰t R N A 位点 ⏹ ●E 位点:中间停靠点,而且当E 位点被占据后,A 位点同氨酰t R N A 的亲和力降低,防止氨酰t R N A 的结合,直到核糖体准备就绪 ⏹ ● m R N A 结合位点2、 蛋白质合成的基本过程⏹ 2.1 肽链的起始:⏹ (1)30S 亚基与m R N A 的结合 ⏹ (2)第一个a a —t R N A 进入核糖体(P 位) ⏹ (3)完整起始复合物的装配2、 核糖体的装配⏹ 核糖体是自组装的结构,没有样板或亲体结构所组成的结构。

Chapter 6 核糖体与核酶6.1核糖体的形态结构核糖体(ribosome)是细胞内一种核糖核蛋白颗粒(ribonucleoprotein partical),是细胞内合成蛋白质的细胞器。
细胞内数量最多的细胞器。
在大肠杆菌中有几万个,占细胞干重的40%,在真核细胞中可达几十万甚至几百万个。
核糖体的主要成分是核糖体RNA(rRNA), 占60%, 蛋白质(r蛋白质), 占40%。
6.1.1 核糖体的类型按存在部位分:细胞质核糖体:游离核糖体和附着核糖体细胞器核糖体:线粒体核糖体和叶绿体核糖体。
按生物类型分两种:原核细胞的核糖体:沉降系数为70S,分子量为2.5x103KDa,由50S和30S 两个亚基组成。
真核细胞的核糖体:沉降系数是80S,分子量为3.9-4.5x103KDa,由60S和40S两个亚基组成。
6.2 核糖体的生物发生(Biogenesis)在细胞内,核糖体是自我装配的。
真核细胞和原核细胞的核糖体合成和装配过程各不相同。
核糖体的生物发生包括蛋白质和rRNA的合成、核糖体亚基的组装。
6.2.1 核糖体基因1. rRNA基因的扩增在染色体上增加rRNA基因的拷贝数:细菌的E.coli的基因组中有七套rRNA 基因;典型的真核生物细胞含有几百到几千个18S、 5.8S和28S rRNA基因的拷贝,5S rRNA基因的拷贝数多达50,000个。
2. rRNA基因的选择性扩增①两栖类卵母细胞rRNA基因扩增基因扩增是通过形成几千个核进行的,每个核里含有几百拷贝的编码18S、5.8S和28S的rRNA基因,最后卵母细胞中的这些rRNA基因的拷贝数几乎达到50万个。
②卵母细胞中rRNA基因扩增机制滚环复制(rolling circle replication)卵母细胞中rRNA基因扩增的机制,有人认为归因于从染色体上分离出来的环状DNA分子,这种环状DNA中含有rRNA基因。
由于环状DNA能够通过滚环复制的方式进行复制,因而能够产生大量的rRNA基因。


第六章.核糖体与核酶核糖体(r i b o s o me),是细胞内一种核糖核蛋白颗粒(r i b o n u c l e o p r o t e i n p a r t i c l e),其惟一功能是按照mR N A的指令将氨基酸合成蛋白质多肽链,所以核糖体是细胞内蛋白质合成的分子机器。
核糖体最早是Al b e r t C l a u d e于20世纪30年代后期发现的,其后又证明了其蛋白质合成功能。
随着分子生物学的发展,核糖体概念的涵意有了进一步的发展。
细胞内除了从事蛋白质合成的核糖体外,还有许多其它功能的核糖核蛋白体颗粒,通常是一些小分子的R N A同蛋白质组成的颗粒,它们参与R N A的加工、R N A的编辑、基因表达的调控等。
发现核糖体及核糖体功能鉴定的两个关键技术是什么?(答案)答:核糖体最早是Al b e r t C l a u d e于1930s后期用暗视野显微镜观察细胞的匀浆物时发现的,当时称为微体(M i c r o s o me s),直到1950s中期,Ge o r g e P a l a d e在电子显微镜下观察到这种颗粒的存在。
当时G e o r g e P a l a d e和他的同事研究了多种生物的细胞,发现细胞质中有类似的颗粒存在,尤其在进行蛋白质合成的细胞中特别多。
后来P h i l i p S i ek e v i t z用亚细胞组份分离技术分离了这种颗粒,并发现这些颗粒总是伴随内质网微粒体一起沉积。
化学分析揭示,这种微粒富含核苷酸,随之命名为r i b o so me,主要成分是核糖体R N A(r R N A),约占60%、蛋白质(r蛋白质)约占40%。
核糖体的蛋白质合成功能是通过放射性标记实验发现的。
将细胞与放射性标记的氨基酸短暂接触后进行匀浆,然后分级分离,发现在微粒体部分有大量新合成的放射性标记的蛋白质。
后将微粒体部分进一步分离,得到核糖体和膜微粒,这一实验结果表明核糖体与蛋白质合成有关。


第六章核糖体与核酶姓名:李淼学号:09352044 班级:生科一班日期:11.17核糖体是细胞内一种核糖蛋白颗粒,含有rRNA和r蛋白质。
核糖体可分为真核生物核糖体和原核生物核糖体,前者有细胞质核糖体、线粒体核糖体和叶绿体核糖体之分。
核糖体均有大小两个亚基组成,进行蛋白质合成时才结合在一起。
原核生物核糖体沉降系数为70S,由50S大亚基(含33种不同的蛋白质以及23S和5S rRNA)和30S小亚基(含21种蛋白质以及16S rRNA)组成;真核生物核糖体沉降系数为80S,由60S大亚基(含大约49种蛋白质以及28S、5S和5.8S rRNA)和40S 小亚基(含大约33种蛋白质以及18S rRNA)组成。
核糖体的组成成分是蛋白质和rRNA,所以编码核糖体的基因分为两类,一类是编码蛋白质的基因,另一类是rRNA基因。
细胞为了满足大量需求的rRNA,有两种方法扩大rRNA 的拷贝数。
第一是在染色体上增加rRNA基因的拷贝数,第二是通过基因扩增来实现。
真核生物的18S、5.8S和28S rRNA基因首先转录成一个45S的前rRNA,能够转录这3个前rRNA的DNA区域称为一个转录单位。
参与rRNA基因转录的酶是RNA聚合酶I,合成地点是核仁,转录间隔区被讲解掉。
原核生物的16S、23S、5S 3种rRNA基因组成一个转录单位。
5S rRNA是核糖体大亚基的一个组分,原核生物和真核生物都有,并且结构相似。
5SrRNA基因是由RNA聚合酶III 在核仁外转录的,只需要进行简单的加工或者不需要加工。
RNA聚合酶III通常是与位于转录部分内的启动子结合,而不是与转录起始位点上游的启动子结合。
核糖体的功能是进行蛋白质多肽链的合成。
核糖体的中有一个mRNA结合位点和3个tRNA结合位点:A、P、E位点。
A位点是氨酰基位点,是与新掺入的氨酰tRNA结合位点,又叫受位。
主要位于大亚基。
P位点是肽酰tRNA位点,又叫供位。
核糖体是细胞内一种核糖核蛋白颗粒,含有rRNA和r蛋白质。
核糖体可分为真核生物核糖体和原核生物核糖体,前者有细胞质核糖体、线粒体核糖体和叶绿体核糖体3中类型。
核糖体均由大小两个亚基组成,进行蛋白质合成时才结合在一起。
Mg2+的浓度对大小亚基的聚合和解离有很大的影响。
原核生物核糖体沉降系数为70S,由50S大亚基(含33种不同的蛋白质以及23S和5SrRNA)he30S小亚基(含21种蛋白质以及16SRNA)组成;真核生物核糖体沉降系数为80S,由60S大亚基(含大约49种蛋白质以及28S、5S和5.8SrRNA)和40S小亚基(含大约33种蛋白质以及18SrRNA)组成。
核糖体的生物发生包括蛋白质和rRNA的合成、核糖体亚基的组装等。
在细胞中增加编码基因的拷贝数有两种方法:一是在染色体上增加rRNA基因的拷贝数;二是通过通过形成几千个核进行基因扩增。
真核生物的18S、5.8S、28SrRNA基因组成一个转录单位,转录成一个45S前体。
前rRNA有两个独特的特点,一是含有大量的甲基化的核苷,另一个就是具有许多假尿苷。
原核生物的16S、23S、5S3种rRNA基因组成一个转录单位。
原核生物rRNA 基因也是多拷贝的,并且也要被转录成一个前体,然后再加工成成熟的rRNA。
细菌的rRNA 也有甲基化,但比真核生物rRNA甲基化程度低。
核糖体是自我装配的细胞器,当蛋白质和rRNA合成加工成熟之后就要开始装配核糖体的大小两个亚基。
在核糖体重新装配过程中,可以通过减去某一种蛋白质来验证该蛋白质在亚基装配及亚基中的作用。
真核生物核糖体亚基在核仁中装配,原核生物核糖体亚基在细胞质中装配。
核糖体的功能是在进行蛋白质多肽链的合成。
在核糖体上合成的只是蛋白质的一级结构,即多肽链。
合成是从多肽链的N端开始,到C端结束。
核糖体中有一个mRNA结合位点和3个tRNA结合位点:A位点、P位点、E位点。
原核生物mRNA通过5’端的SD序列与核糖体16SrRNA结合,真核生物则依赖于mRNA5’端甲基化帽子结构将mRNA与核糖体小亚基结合。
细胞生物学章节提要第六章核糖体与核酶细胞生物学章节提要第六章核糖体与核酶第06章总结核糖体与核酶研究方法:足迹、放射性标记(核糖体发现)、原核核糖体重组实验和合成反义RNA干扰。
核糖体(ribsome)是细胞内一种核糖蛋白颗粒(ribonucleoproteinparticle),唯一功能是翻译mrna的指令,将氨基酸合成蛋白质多肽链。
核糖体包括线粒体核糖体、叶绿体核糖体、细胞质核糖体。
真核细胞和原核细胞的核糖体组成不同,由均有两个不同大小的亚基构成。
它有rrna和核糖体蛋白质(ribosomalprotein),原核细胞由50s和30s 两个亚基构成70s的核糖体,真核细胞由60s和40s两个亚基构成80s的核糖体。
核糖体生物发生包括蛋白质和核糖核酸的合成、核糖体亚基的组装等。
在进化过程中,通过增加染色体上rRNA基因的拷贝数和基因扩增,编码rRNA的基因数量增加。
但第一个含有rRNA基因的DNA是如何形成的尚不清楚。
真核18S、5.8S和28s rRNA基因形成一个转录单元,这是转录成45s的前提。
原核生物的16S、23S和5S rRNA基因构成一个转录单元。
与成熟rRNA相比,真核生物的前rRNA含有大量甲基化核苷和大量假尿苷。
原核生物的rRNA基因也是多拷贝的(在真核生物中重复率相对较低),需要转录成前体,然后加工成rRNA。
此外,在真核生物中编码5srrna的基因位于不同的染色体上,而细菌5srrna基因与其他两个rRNA结合形成一个转录单元。
在组装部位,真核生物在核仁中,原核生物在细胞质中。
核糖体的唯一功能是蛋白质合成(proteinsynthetize)。
核糖体中与trna结合位点有a位点(asite)、p位点(psite)、e位点(esite)。
原核生物通过sd序列识别结合,真核生物通mrna5’端甲基化帽子结构识别。
蛋白质合成主要的过程为:链的起始、链的延伸、链的终止。
大约90%以上的真核生物的起始密码子是aug。
6核糖体与核酶1.核糖体(rib osome)核糖体是细胞内一种核糖核蛋白颗粒(ribonucleoprotein particle), 其惟一功能是按照mRNA的指令将氨基酸合成蛋白质多肽链,所以核糖体是细胞内蛋白质合成的分子机器。
按核糖体存在的部位可分为三种类型:细胞质核糖体、线粒体核糖体、叶绿体核糖体。
按存在的生物类型可分为两种类型:真核生物核糖体和原核生物核糖体。
原核细胞的核糖体较小, 沉降系数为70S,相对分子质量为2.5x103kDa,由50S和30S两个亚基组成; 而真核细胞的核糖体体积较大, 沉降系数是80S,相对分子质量为3.9~4.5x103 kDa, 由60S和40S两个亚基组成。
在真核细胞中, 核糖体进行蛋白质合成时,既可以游离在细胞质中, 称为游离核糖体, 也可以附着在内质网的表面, 称为膜旁核糖体或附着核糖体。
真核细胞含有较多的核糖体, 每个细胞平均有106~107个, 而原核细胞中核糖体较少每个细胞平均只有15×102~18×103个。
典型的原核生物大肠杆菌核糖体是由50S大亚基和30S小亚基组成的。
在完整的核糖体中,rRNA约占2/3, 蛋白质约为1/3。
50S大亚基含有34种不同的蛋白质和两种RNA分子,相对分子质量大的rRNA的沉降系数为23S,相对分子质量小的rRNA为5S。
30S小亚基含有21种蛋白质和一个16S的rRNA 分子。
真核细胞核糖体的沉降系数为80S,大亚基为60S,小亚基为40S。
在大亚基中,有大约49种蛋白质,另外有三种rRNA∶28S rRNA、5S rRNA和5.8S rRNA。
小亚基含有大约33种蛋白质,一种18S的rRNA。
2.基因扩增(gene amp li f icat ion)细胞内选择性复制DNA, 产生大量的拷贝。
如两栖类卵母细胞在发育的早期,rRNA基因的数量扩增到1000多倍。
基因扩增是通过形成几千个核进行的,每个核里含有几百拷贝的编码28S、18S和5.8S的rRNA基因,最后卵母细胞中的这些rRNA基因的拷贝数几乎达到50万个,而在相同生物的其它类型细胞中,这些rRNA基因的拷贝数只有几百个。
卵母细胞中有如此众多的rRNA基因拷贝,为卵细胞在受精后的发育过程中合成大量核糖体创造了条件。
至于卵母细胞中rRNA基因扩增的机制,有人认为可能是通过从染色体上分离出来的环状DNA分子,这种环状DNA中含有rRNA基因,但是第一个含有rRNA基因的环状DNA是如何形成的尚不清楚。
由于环状DNA能够通过滚环复制(rolling circle replication)的方式进行复制,因而能够产生大量的rRNA 基因。
3. 5S rRNA基因(5S rRNAgene)5S rRNA是核糖体大亚基的一个组份,原核生物和真核生物都有5S rRNA,而且结构相似。
真核生物的5S rRNA基因与其它三种rRNA基因不在同一条染色体上,它是由核仁以外的染色体基因转录的,然后运输到核仁内参与核糖体的装配。
5S rRNA基因的数量比45S rRNA转录单位多,人的5S rRNA基因有500个拷贝,并且在染色体上串连排列。
非洲爪蟾的5S rRNA基因的一个重复单位含有一个5S rRNA基因、一个不转录的假基因(101bp的5S rRNA基因的片段),每个重复单位间被不转录的间隔序列隔开,间隔序列的长度变化不定,最长达400bp;5S rRNA基因转录的速度很快,其结果产生过量的5S rRNA,有些最后要被降解掉。
5S rRNA基因是由RNA聚合酶Ⅲ转录的,原初转录物的5'端与成熟的5S rRNA的5'端完全相同。
在某些生物中,3'端通常含有多余的核苷酸,在加工时要被切除。
所以,5S rRNA只需要进行简单的加工,或者根本不需要进行加工。
4. A位点(A s ite)即氨酰基位点,是与新掺入的氨酰tRNA(aminoacyl-tRNA )结合的位点, 又叫受位(entry site),主要位于大亚基,是接受氨酰tRNA的部位。
5. P位点(P s ite)即肽酰tRNA位点(peptidyl-tRNA site), 又叫供位(donor site), 或肽酰基位点, 主要位于大亚基, 是肽基tRNA移交肽链后肽酰tRN A所占据的位置, 即与延伸中的肽酰tRNA结合位点。
6. E 位点(exit site, E s it e)E位点是脱氨酰tRNA(deaminoacyl-tRNA)离开A位点到完全从核糖体释放出来的一个中间停靠点,只是作暂时的停留。
当E位点被占据之后,A位点同氨酰tRNA的亲和力降低,防止了氨酰tRNA的结合,直到核糖体准备就绪,E位点腾空,才会接受下一个氨酰tRNA。
7. SD序列(SD sequence)在原核生物中, 核糖体中与mRNA结合位点位于16S rRNA 的3'端,mRNA 中与核糖体16S rRNA结合的序列称为SD序列(SD sequence),它是1974年由J.Shine 和 L.Dalgarno发现的,故此而命名。
SD序列是mRNA中5'端富含嘌呤的短核苷酸序列,一般位于mRNA的起始密码AUG的上游5~10个碱基处,并且同16S rRNA 3'端的序列互补。
8.多聚核糖体(polyr ibosomes)在蛋白质合成过程中,同一条mRNA分子能够同多个核糖体结合,同时合成若干条蛋白质多肽链,结合在同一条mRNA上的核糖体就称为多聚核糖体(polysome 或polyribosomes)。
在mRNA的起始密码子部位,核糖体亚基装配成完整的起始复合物,然后向mRNA的3'端移动,直到到达终止密码子处。
当第一个核糖体离开起始密码子后,空出的起始密码子的位置足够与另一个核糖体结合时,第二个核糖体的小亚基就会结合上来,并装配成完整的起始复合物,开始蛋白质的合成。
同样,第三个核糖体、第四个核糖体、……依次结合到mRNA上形成多聚核糖体。
根据电子显微照片推算,多聚核糖体中,每个核糖体间相隔约80个核苷酸。
9.嘌呤霉素(puromyc in)嘌呤霉素是一种蛋白质合成抑制剂,它具有与tRNA分子末端类似的结构, 能够同氨基酸结合,代替氨酰化的tRNA同核糖体的A位点结合,并掺入到生长的肽链中。
虽然嘌呤霉素能够同A位点结合,但是不能参与随后的任何反应, 因而导致蛋白质合成的终止并释放出C-末端含有嘌呤霉素的不成熟的多肽。
10. N-端规则(N-end ru le)每一种蛋白质都有寿命特征, 称为半衰期(half-life)。
蛋白质的半衰期与多肽链N-端特异的氨基酸有关,它们对蛋白质的寿命有控制作用。
如末端是精氨酸或赖氨酸的多肽,寿命就很短,而末端是缬氨酸或甲硫氨酸的多肽,寿命就很长。
蛋白质N-末端与半衰期的关系,称为N端规则。
11.蛋白酶体(pro teasomes)蛋白酶体既存在于细胞核中,又存在于胞质溶胶中, 是溶酶体外的蛋白水解体系, 由10~20个不同的亚基组成中空的圆桶形的结构,显示多种肽酶的活性,能够从碱性、酸性和中性氨基酸的羧基侧水解多种与遍在蛋白连接的蛋白质底物。
蛋白酶体对蛋白质的降解是与环境隔离的。
主要降解两种类型的蛋白质:一类是错误折叠的蛋白质,另一类就是需要进行数量调控的蛋白质。
蛋白酶体对蛋白质的降解通过泛素(ubiquitin)介导,所以又称为泛素降解途径。
泛素是由76个氨基酸残基组成的小肽,它的作用主要是识别要被降解的蛋白质,然后将这种蛋白质送入蛋白酶体的圆桶中进行降解。
蛋白酶体对蛋白质的降解作用分为两个过程:一是对被降解的蛋白质进行标记,由泛素完成;二是蛋白酶解作用,由蛋白酶体催化。
蛋白酶体存在于所有真核细胞中,其活性受γ干扰素的调节。
12.核酶(r ibozyme)核酶一词用于描述具有催化活性的RNA, 即化学本质是核糖核酸(RNA), 却具有酶的催化功能。
核酶的作用底物可以是不同的分子, 有些作用底物就是同一RNA分子中的某些部位。
核酶的功能很多,有的能够切割RNA, 有的能够切割DNA, 有些还具有RNA 连接酶、磷酸酶等活性。
与蛋白质酶相比,核酶的催化效率较低,是一种较为原始的催化酶。
13.小分子RNA(smal l RNA)存在于真核生物细胞核和细胞质中,它们的长度为100到300个碱基(酵母中最长的约1000个碱基)。
多的每个细胞中可含有105~106个这种RNA分子,少的则不可直接检测到, 它们由RNA聚合酶Ⅱ或RNA聚合酶Ⅲ所合成, 其中某些象mRNA一样可被加帽。
主要有两种类型的小分子RNA:一类是snRNA(small nuclear RNA),存在于细胞核中;另一类是scRNA(small cytoplasmic RNA),存在于细胞质中。
小分子RNA通常与蛋白质组成复合物, 在细胞的生命活动中起重要的作用, 某些snRNPs和剪接作用密切相关,它们分别与供体和受体剪接位点以及分支顺序相互补。
scRNA参与蛋白质的合成和运输, 如SRP颗粒就是一种由一个7SRNA和蛋白质组成的核糖核蛋白体颗粒,主要功能是识别信号肽, 并将核糖体引导到内质网。
14.反义RNA(ant isense RNA)反义RNA是指与mRNA互补的RNA分子, 也包括与其它RNA互补的RNA 分子。
由于核糖体不能翻译双链的RNA,所以反义RNA与mRNA特异性的互补结合, 即抑制了该mRNA的翻译。
通过反义RNA控制mRNA的翻译是原核生物基因表达调控的一种方式,最早是在E.coli 的产肠杆菌素的Col E1质粒中发现的,许多实验证明在真核生物中也存在反义RNA。
近几年来通过人工合成反义RNA的基因, 并将其导入细胞内转录成反义RNA, 即能抑制某特定基因的表达,阻断该基因的功能, 有助于了解该基因对细胞生长和分化的作用。
细胞中反义RNA的来源有两种途径∶第一是反向转录的产物,在多数情况下, 反义RNA是特定靶基因互补链反向转录产物, 即产生mRNA和反义RNA的DNA是同一区段的互补链。
第二种来源是不同基因产物,如OMPF基因是大肠杆菌的膜蛋白基因,与透性有关,其反义基因MICFZE则为另一基因。
15.内含子(in tron)内含子是基因内的间隔序列,不出现在成熟的RNA分子中,在转录后通过加工被切除。
大多数真核生物的基因都有内含子。
16.外显子(exon)外显子是最后出现在成熟RNA中的基因序列, 又称表达序列。
17.自我剪接(self-spl ici ng)具有自我催化能力,将自身的某些部位切除的现象称为自我剪接。
在酵母和真菌的线粒体mRNA和tRNA前体加工、叶绿体的tRNA 和rRNA前体加工、某些细菌病毒的mRNA前体加工中都发现了自我剪接现象。