甜蛋白基因MBLII 对莴苣的遗传转化
- 格式:pdf
- 大小:200.36 KB
- 文档页数:5

甜樱桃类钙调蛋白基因家族的生物信息学分析
吴军平;付全娟;郭远鹏;侯森;孙玉刚;魏国芹
【期刊名称】《落叶果树》
【年(卷),期】2024(56)2
【摘要】类钙调蛋白(CML)是植物生长发育过程中应对刺激的一种重要蛋白,基于甜樱桃全基因组,利用生物信息学的方法对甜樱桃CML基因家族成员进行鉴定,并对理化性质、染色体定位、基因结构、启动子作用元件等进行分析。
结果表
明,PavCML家族有49个基因,不均匀的分布于1~8号染色体上,亚细胞定位在细胞质、叶绿体、细胞核等部位,启动子顺式作用元件分析发现PavCML家族基因响应多种元件。
与转录组数据结合进行荧光定量PCR分析,结果显示CML在裂果部位的相对表达量高,表明该基因可能参与裂果过程,为CML家族基因的功能鉴定提供理论依据。
【总页数】8页(P32-39)
【作者】吴军平;付全娟;郭远鹏;侯森;孙玉刚;魏国芹
【作者单位】山东农业大学;山东省果树研究所;鄄城县国有第一林场
【正文语种】中文
【中图分类】S662.5
【相关文献】
1.甜荞热激转录因子基因家族的鉴定及生物信息学分析
2.陆地棉类谷氧还蛋(GhGRL)基因家族生物信息学分析
3.甜樱桃PaGAST基因的电子克隆及生物信息
学分析4.枣类甜蛋白基因家族的鉴定与生物信息学分析5.番木瓜赖氨酸基序类受体激酶LysM-RLK基因家族鉴定及生物信息学分析
因版权原因,仅展示原文概要,查看原文内容请购买。

《CmBTB4与CmBTB32基因在甜瓜发育中功能的初步分析》篇一一、引言甜瓜作为重要的经济作物,其发育过程中的基因调控机制一直是研究的热点。
近年来,随着分子生物学技术的快速发展,越来越多的基因被克隆并进行了功能分析。
其中,CmBTB4与CmBTB32基因在甜瓜发育中起着重要的作用。
本文将就这两个基因在甜瓜发育中的功能进行初步分析。
二、研究背景与目的BtB基因家族是一类广泛存在于植物中的转录调控因子,其在植物生长发育和应激反应中具有重要作用。
CmBTB4与CmBTB32作为甜瓜中的BtB基因家族成员,其功能尚未明确。
本研究旨在初步分析这两个基因在甜瓜发育中的功能,为进一步了解甜瓜生长发育的分子机制提供理论依据。
三、研究方法1. 基因克隆:通过PCR、RACE等技术手段,克隆出CmBTB4与CmBTB32基因的cDNA序列。
2. 生物信息学分析:利用生物信息学软件,对两个基因的序列进行结构分析、保守结构域预测等。
3. 表达模式分析:通过实时荧光定量PCR技术,检测两个基因在甜瓜不同组织、不同发育阶段的表达情况。
4. 转基因实验:构建过表达和敲低表达载体,通过遗传转化技术将载体导入甜瓜中,观察转基因甜瓜的表型变化。
四、结果与分析1. 基因结构与序列分析通过对CmBTB4与CmBTB32基因的cDNA序列进行克隆和测序,我们发现两个基因均具有典型的BtB基因家族特征,包括保守的BTB结构域和Zn-finger结构域。
进一步的分析表明,两个基因在染色体上的位置和表达模式可能存在差异。
2. 表达模式分析实时荧光定量PCR结果显示,CmBTB4与CmBTB32基因在甜瓜的不同组织、不同发育阶段中均有表达,且表达模式存在差异。
其中,CmBTB4基因在果实发育早期高表达,而CmBTB32基因则在果实成熟期高表达。
这表明两个基因在甜瓜发育过程中可能具有不同的功能。
3. 转基因实验分析通过构建过表达和敲低表达载体并导入甜瓜中,我们观察到转基因甜瓜的表型发生了明显变化。


作物学报ACTA AGRONOMICA SINICA 2008, 34(3): 349−354 /zwxb/ ISSN 0496-3490; CODEN TSHPA9E-mail: xbzw@DOI: 10.3724/SP.J.1006.2008.00349小麦类甜蛋白基因的转化及转基因植株的抗病性分析邢莉萍王华忠蒋正宁倪金龙曹爱忠于玲陈佩度*(南京农业大学作物遗传与种质创新国家重点实验室, 江苏南京 210095)摘要:目前已克隆出许多类甜蛋白基因并成功转化马铃薯、水稻和小麦等植物, 均可引起植株抗病性的提高。
在以往的研究中我们从小麦中克隆了一个类甜蛋白基因(Ta-Tlp), 该基因在高抗白粉病的小麦6VS/6AL易位系中能够高水平表达, 推测它与白粉病抗性密切相关。
为进一步研究该基因的功能, 本研究构建了包含在ubi强启动子控制下的Ta-Tlp基因的表达载体pAHC-TlP, 用基因枪将其导入小麦品种扬麦158的幼胚愈伤组织, 在含除草剂的选择培养基上经两轮筛选和分化, 获得再生抗性植株。
对T0、T1和T2代植株进行PCR、Southern杂交与RT-PCR分析, 结果表明Ta-Tlp基因已整合进受体基因组并且能够表达。
对T0代及其衍生的T1代(来自6个T0代阳性株系的183个单株)和T2代(来自6个T0代阳性株系的241个单株)转化植株进行白粉病抗性鉴定, Ta-Tlp基因超量表达的转基因植株, 能延缓白粉病发病进程, 其白粉病抗性有一定提高。
对T2代阳性转化植株进行赤霉病抗性鉴定表明, 其抗性与受体对照相比无显著差异。
关键词:小麦类甜蛋白基因; 基因枪; 遗传转化; 白粉病; 抗性鉴定Transformation of Wheat Thaumatin-Like Protein Gene and Diseases Resistance Analysis of the Transgenic PlantsXING Li-Ping, WANG Hua-Zhong, JIANG Zheng-Ning, NI Jin-Long, CAO Ai-Zhong, YU Ling, and CHEN Pei-Du*(National Key Laboratory of Crop Genetics and Germplasm Enhancement, Nanjing Agricultural University, Nanjing 210095, Jiangsu, China)Abstract: The thaumatin-like protein in plants, an important member of pathogenesis-related proteins (PRs), involves in plant resistant reaction to many fungi. A series of thaumatin-like protein genes have been isolated and transformed to potato (Solanum tuberosum L.), rice (Oryza sativa L.), and wheat (Triticum aestivum L.) resulting in resistance improvement to plant diseases. We have cloned a thaumantin-like protein gene, Ta-Tlp, from wheat in our earlier studies. The Ta-Tlp gene expresses in high level in wheat 6VS/6AL translocation line with high resistance to powdery mildew (Erysiphe graminis f. sp. tritici Em. Marchal.), imply-ing its close relation to the resistance of the disease. To further understand the gene’s function, we constructed Ta-Tlp into a ex-pression vector driven by the strong ubi promoter in the present study. The constructed vector pAHC-TlP was transformed into immature embryo-derived calli of a common wheat cultivar Yangmai 158 through particle bombardment. After two rounds of herbicide bialaphos selection and regeneration, herbicide-resistance plants were obtained. The Ta-Tlp was confirmed having been integrated into the wheat genome and expressed in T1 and T2 generations by PCR, Southern blot, and RT-PCR. The transgenic plants of T0, T1, and T2 generations were inoculated by E. graminis f. sp. tritici and F. graminearum for resistance identification.All plants of T0, T1, (183 individuals derived from 6 positive T0 lines), and T2 (241 individuals derived from 6 positive T0 lines) generations appeared some resistance to wheat powdery mildew by delaying disease development, but no distinct resistance to scab (F. graminearum).Keywords: Wheat thaumatin-like protein gene (Ta-Tlp); Particle bombardment; Genetic transformation; Powdery mildew; Resistance identification基金项目:国家高技术研究发展计划(863计划)项目(2004AAA222140, 2006AA10A104);江苏省自然科学基金项目(BK2007163);教育部长江学者和创新团队发展计划项目(10418)作者简介:邢莉萍(1981–), 女, 江苏人, 在读博士, 专业方向:植物基因工程。

莴苣线粒体基因组密码子使用偏好性分析张扬;孙曙光;李晴;魏珍【期刊名称】《河南农业科学》【年(卷),期】2022(51)10【摘要】为了分析栽培莴苣Lactuca sativa‘Salinas’和野生莴苣Lactuca saligna的线粒体基因组密码子使用特征差异,对2种莴苣的线粒体基因组编码序列进行密码子使用偏好性研究,比较其密码子偏好性形成的影响因素和演化过程。
结果表明:栽培莴苣和野生莴苣线粒体编码基因的总GC含量分别为43.43%和42.54%,富含A/T碱基。
2种莴苣的有效密码子数(ENC)都在36.61~61.00,密码子偏好性较弱。
中性绘图分析表明,2种莴苣线粒体编码基因的第1、2位和第3位碱基GC含量较为保守,选择是影响2种莴苣线粒体基因密码子使用偏好性的主要因素。
PR2-plot分析表明,2种莴苣线粒体基因第3位碱基更偏好使用嘧啶碱基。
ENC-plot分析结果显示,2种莴苣的线粒体基因密码子偏好性主要受到选择压力影响。
对2种莴苣的相对同义密码子使用度(RSCU)分析表明,栽培莴苣和野生莴苣分别有28个和29个高频密码子,除亮氨酸外,2种莴苣的高频密码子均以A/U结尾。
2种莴苣线粒体编码基因的密码子使用偏好性总体上较接近,但最优密码子存在较大差异。
这种差异可能是因为2种莴苣的低表达基因库差异造成的,也可能与选择压力及人工育种对栽培莴苣线粒体基因组进化的影响有关。
【总页数】11页(P114-124)【作者】张扬;孙曙光;李晴;魏珍【作者单位】河南省农业科学院;河南省作物分子育种研究院;郑州大学生命科学学院【正文语种】中文【中图分类】S636.2【相关文献】1.稻瘟病病菌线粒体基因密码子使用偏好性分析2.普通野生稻线粒体蛋白质编码基因密码子使用偏好性的分析3.普通野生稻线粒体蛋白质编码基因密码子使用偏好性的分析4.栽培大豆和野生大豆线粒体基因组密码子使用偏性的比较分析5.澳洲坚果光壳种叶绿体基因组的密码子使用偏好性及其影响因素分析因版权原因,仅展示原文概要,查看原文内容请购买。
《甜瓜CmPYL4、CmPYR1基因在果实成熟过程中功能的初步分析》篇一摘要:本研究初步分析了甜瓜(Cucumis melo)中的CmPYL4和CmPYR1基因在果实成熟过程中的功能。
通过生物信息学分析、基因表达模式研究以及转基因实验,揭示了这两个基因在甜瓜果实成熟过程中的潜在作用。
本文为进一步了解甜瓜果实成熟机制及遗传改良提供了理论依据。
一、引言甜瓜作为一种重要的经济作物,其果实的成熟过程直接关系到果实的品质和经济效益。
近年来,植物激素信号转导途径在果实成熟过程中的作用受到了广泛关注。
PYL(Pyrabactin-like)和PYR(Pyrabactin resistance)基因作为植物激素信号的关键调控因子,对果实成熟具有重要的影响。
因此,本研究选取甜瓜的CmPYL4和CmPYR1基因,探讨其在果实成熟过程中的功能。
二、材料与方法1. 材料实验材料为不同成熟阶段的甜瓜果实,以及相应的转基因植物材料。
2. 方法(1)生物信息学分析:利用生物信息学软件对CmPYL4和CmPYR1基因进行序列分析、表达模式预测等。
(2)基因表达模式研究:采用实时荧光定量PCR技术,分析不同成熟阶段甜瓜果实中这两个基因的表达情况。
(3)转基因实验:构建转基因植物,研究CmPYL4和CmPYR1基因过表达或敲除对果实成熟的影响。
三、结果与分析1. 生物信息学分析结果CmPYL4和CmPYR1基因具有典型的PYL/PYR基因特征,其序列保守性高,可能与植物激素信号转导密切相关。
表达模式预测显示,这两个基因在果实成熟过程中表达量发生变化。
2. 基因表达模式研究结果实时荧光定量PCR结果显示,CmPYL4和CmPYR1基因在甜瓜果实成熟过程中表达量呈现明显变化,与果实的成熟度呈正相关。
这表明这两个基因可能参与调控甜瓜果实的成熟过程。
3. 转基因实验结果(1)过表达实验:过表达CmPYL4和CmPYR1基因的转基因甜瓜果实表现出提前成熟的趋势,果实的糖分、风味等品质得到改善。
第43卷第4期2021年10月Vol.43,No.4Oct.,2021中国糖料Sugar Crops of China doi :10.13570/ki.scc.2021.04.002甜菜BTB 蛋白家族的挖掘与生物信息学分析刘宇1,2,3,4,5,杨巧1,2,3,4,5,王晓丹1,2,3,4,5,赵春雷1,2,3,4,5,王希1,2,3,4,5,李彦丽1,2,3,4,5,贾海伦1,2,3,4,5,丁广洲1,2,3,4,5,陈丽1,2,3,4,5(1.黑龙江大学现代农业与生态环境学院,哈尔滨150080;2.黑龙江省甜菜工程技术研究中心,哈尔滨150080;3.国家糖料改良中心,哈尔滨150080;4.黑龙江省普通高等学校甜菜遗传育种重点实验室,哈尔滨150080;5.国家甜菜种质中期库,哈尔滨150080)摘要:BTB (Broad-complex,tramtrack,and bric-à-brac )蛋白在植物生长发育、抗逆性、蛋白质泛素化和降解、细胞骨架组成、离子通道及细胞周期调控等方面发挥着重要作用。
本研究从甜菜基因组大数据中获得了49个包含BTB 结构域的序列,并利用生物信息学方法对甜菜BTB 蛋白家族进行了系统发育、理化性质、基因结构、保守结构域、染色体定位等分析。
结果显示:甜菜BTB 家族编码的氨基酸大部分都属于酸性氨基酸,氨基酸数目为139~1120个,翻译的蛋白大部分为亲水性蛋白,可分为9个亚家族(Ankyrin 、Armadillo 、MATH 、NPH3、BACK 、TAZ 、TPR 、BTB-only 、Other ),在染色体上呈不均匀分布,主要分布在第5~7号染色体上;亚细胞定位的结果发现36个BTB 蛋白都位于细胞核,其余的位于细胞膜和叶绿体上;49个BTB 蛋白的磷酸化位点数目为3~25个,不同蛋白磷酸化位点数目差异很大,49个BTB 蛋白的等电点为4.70~9.51,分子量为15848.66~124344.33Da ;甜菜BTB 家族不同亚族的基因结构存在较大差异,不同亚族的内含子数量和长度都不同;预测的二级结构和三级结构结果表明,不同亚家族的二级结构以BTB 结构域为主,但不同亚族还包含了不同的结构域;三级结构中不同结构的数量虽存在着差异,但都以α螺旋和β折叠为主。
《甜瓜CmPYL4、CmPYR1基因在果实成熟过程中功能的初步分析》篇一一、引言甜瓜作为重要的经济作物之一,其果实成熟过程中的品质和风味变化一直是科研人员关注的焦点。
近年来,随着分子生物学技术的不断发展,基因功能的研究逐渐成为甜瓜果实成熟机制研究的重要手段。
本文旨在初步分析甜瓜CmPYL4和CmPYR1基因在果实成熟过程中的功能,以期为甜瓜品质改良提供理论依据。
二、材料与方法1. 材料本实验选取了不同成熟阶段的甜瓜果实作为实验材料,包括未成熟、成熟和过熟三个阶段的果实。
2. 方法(1)基因克隆与序列分析:通过PCR技术克隆出CmPYL4和CmPYR1基因的cDNA序列,并进行序列分析。
(2)表达模式分析:利用实时荧光定量PCR(RT-PCR)技术,分析CmPYL4和CmPYR1基因在不同成熟阶段甜瓜果实中的表达情况。
(3)转基因技术:构建CmPYL4和CmPYR1基因的过表达和敲除载体,通过遗传转化技术获得转基因甜瓜植株。
(4)表型分析:对转基因甜瓜植株的果实成熟过程进行观察,并记录相关表型数据。
三、结果与分析1. 基因序列分析通过序列分析发现,CmPYL4和CmPYR1基因的cDNA序列具有较高的保守性,与已知的植物PYL和PYR基因家族成员具有较高的相似性。
这表明这两个基因可能具有相似的生物学功能。
2. 表达模式分析RT-PCR结果显示,CmPYL4和CmPYR1基因在甜瓜果实成熟过程中表达量发生变化。
在果实未成熟阶段,两个基因的表达量较低;随着果实的成熟,表达量逐渐升高;在过熟阶段,表达量又有所下降。
这表明这两个基因可能参与甜瓜果实的成熟过程。
3. 转基因技术分析通过构建过表达和敲除载体,我们获得了转基因甜瓜植株。
对转基因植株的果实成熟过程进行观察发现,过表达CmPYL4和CmPYR1基因的植株果实成熟速度较快,果肉色泽和风味品质得到改善;而敲除这两个基因的植株则表现出相反的表型,即果实成熟速度减慢,果肉色泽和风味品质变差。
重组甜蛋白Brazzein和MBLⅡ基因植物表达载体构建及人参果再生体系的建立重组甜蛋白Brazzein和MBLⅡ基因植物表达载体构建及人参果再生体系的建立人参果(Panax ginseng C.A.Meyer)是一种重要的中药材和保健食品原料,其具有许多生物活性成分,如人参皂苷、多糖等,对人体健康具有积极的影响。
然而,由于人参果生长周期长,繁殖速度慢,产量有限,导致其资源紧缺。
因此,开展人参果的遗传改良和基因工程研究对于提高其产量和质量具有重要意义。
甜蛋白Brazzein和MBLⅡ (MonellinⅡ)是一类天然的甜味蛋白,其甜味相当于蔗糖的3000-8000倍,且不会引起血糖升高,是一种理想的天然甜味剂。
由于其良好的甜味特性,Brazzein和MBLⅡ被广泛应用于食品和饮料工业。
将Brazzein和MBLⅡ基因导入人参果中,可以赋予人参果甜味,提高其消费者认可度和市场竞争力。
在本研究中,我们通过遗传工程方法构建了重组甜蛋白Brazzein和MBLⅡ基因植物表达载体。
首先,我们从甜瓜(Cucumis melo)和蜜瓜(Cucumis melo L.)中分离得到了Brazzein和MBLⅡ基因序列。
然后,将这两个基因序列克隆并与植物表达载体pCAMBIA1301进行连接,构建了Brazzein和MBLⅡ基因的植物表达载体pCAMBIA1301-Brazzein和pCAMBIA1301-MBLⅡ。
接下来,我们建立了人参果的再生体系。
选取健壮的人参果子实为材料,经过表皮消毒后,将其以4℃贮存。
接种时,将人参果子实表皮去除,取出种子,进行无菌处理。
然后,将种子接种于含有植物生长调节剂的培养基中,培养基的配方为MS+6-BA 2.0 mg/L+NAA 0.5 mg/L+sucrose 30 g/L+agar 7g/L。
将接种的种子置于暗室中,进行初级培养。
在发芽时,将初级培养基转移到含有低浓度抗生素的培养基中,以选择带有Brazzein和MBLⅡ基因的转基因植物。
园 艺 学 报 2001,28(3):246~250Acta Horticulturae Sinica收稿日期:2000-11-01;修回日期:2001-03-27基金项目:北京市重点实验室基金资助项目(951890200)甜蛋白基因MBL II 对莴苣的遗传转化刘敬梅1,2 陈大明2 陈 杭2(1浙江大学园艺系,杭州310029;2北京市农林科学院蔬菜研究中心,北京100089)摘 要:以4日龄莴苣(Lactua sativa L.)无菌苗子叶为外植体,通过根癌农杆菌介导,成功地进行了马槟榔甜蛋白基因MBL II 对莴苣的遗传转化。
抗生素浓度和子叶外植体与农杆菌的共培养时间是影响转化的重要因素。
附加卡那霉素(Kan)50mg/L 的诱芽培养基MS I(MS+NAA 0.1mg/L+6 BA 0.1mg/L +羧卞青霉素500mg/L)最适于侵染后子叶外植体的诱芽培养。
在外植体与农杆菌共培养0~7d 的范围内,以共培养3d 最佳(生芽率58.3%,白化率29%)。
1~2cm 再生芽移入诱根培养基MS II (MS+NAA 0.05mg/L +Kan 50mg/L+羧苄青霉素300mg/L)中,诱根率可达100%。
获得的抗性植株经组织化学及PCR 特异扩增鉴定和统计,7.6%阳性。
Southern blot 结果证明MB L II 基因已整合到莴苣基因组中。
关键词:甜蛋白;莴苣;遗传转化中图分类号:S 636.2;Q 785 文献标识码:A 文章编号:0513 353X (2001)03 0246 05关于莴苣的组织培养和遗传转化已有成功的报道 1~3 。
Curtis 等将硝酸还原酶基因导入4个莴苣主要栽培种,降低了莴苣叶片的硝酸盐含量,改善了莴苣的品质,提高了食用价值和经济价值。
莴苣的口味直接影响其受喜爱的程度。
Samuel Sun 等 4 从中国云南省马槟榔(Capperis masaikai )的种子中克隆了甜蛋白基因MB L II,该蛋白的甜度是相同质量蔗糖的400倍,而且甜味持久。
作者采用根癌农杆菌介导的遗传转化方法,将MB L II 基因导入莴苣中,为利用基因工程改良莴苣风味品质进行了探索。
1 材料与方法1.1 植物材料将莴苣品种!北山三号∀种子用5%NaClO 灭菌15min 后,无菌水冲洗4~5次,置于1/2MS 培养基上,黑暗下28#萌发。
4d 后切取子叶,进行遗传转化。
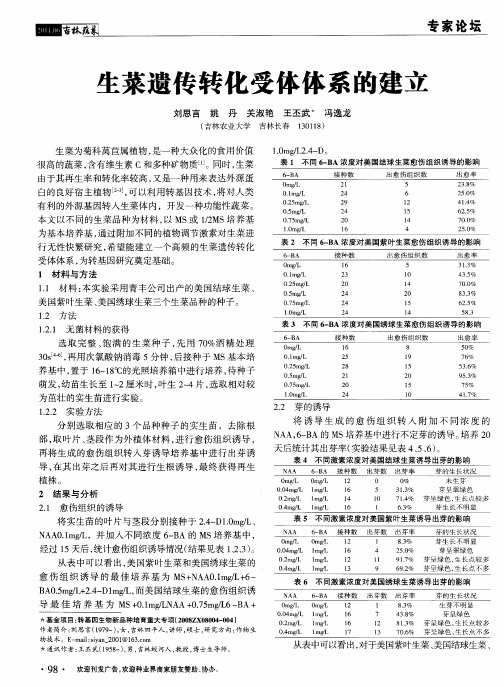
1.2 质粒和菌株图1 质粒pL X10(1.6kb 嵌合基因MBL II 位于质粒pBI121上)Fig.1 pLX10(1.6kb chimeric MBL II gene in pBI 121)根癌农杆菌菌株LB A4404,含双元载体,其中质粒pLX10含有MBL II 、NP T II 和GUS 基因(见图1)。
该基因由美国夏威夷大学植物分子生理学系教授Samuel Sun 提供。
1.3 共培养及抗性芽筛选莴苣子叶外植体去尖端和基端,浸入28#液体YEB 培养基中培养2d 的根癌农杆菌菌液中,1h 后取出,用无菌滤纸吸干菌液,28#培养基上暗培养0、1、3、5、7d 后,转移到含卡那霉素(Kan)0(对照)、25、50、100、200、250mg/L 的筛选培养基MSI 上筛选转化芽。
20d 左右,在外植体切口边缘的白色愈伤组织上长出1~2cm 芽。
将芽切下放入MS II 培养基(MS+NAA 0.05mg/L+Kan 50mg/L+羧苄青霉素300mg/L)中诱根。
生根后将小植株先移入含Hogland 和Snyde 营养液的蛭石中炼苗10~15d,再移入土壤,置于温室自然光照下生长。
1.4 GU S 检测GU S 活性采用Jefferson 等 5 的方法检测。
选取Kan 抗性芽叶片,浸入GUS 染色液(100mmol/L 磷酸缓冲液,0.1%Triton X 100, 1.0mmol/L EDTA,0.5mmol/L K 3Fe(CN)6,0.05mmol/L K 4(CN)6,1m mol/L X Gluc)中,37#保温3~5h,吸弃染色液,加入70%酒精脱色,待绿色褪尽,整张叶片或叶脉处呈现深兰色者为阳性。
1.5 PCR 扩增1.5.1 引物设计 根据MB L II 序列设计一对特异引物P 1、P 2,序列如下:P 1:5∀ACC TTGGC TCTC TTCGTTC T 3∀;P 2:5∀GC TCCGATGTTGGGTATGTT 3∀。
1.5.2 DNA 快速提取及PCR 从GU S 染色阳性植株上取半片小幼叶,放入1.5mL 离心管中,加入100 L 0.5mol/L Na OH 溶液,匀浆,立即13000g 离心1min,取5 L 上清液至一个含45 L 100m mol/L Tris HCl (pH 8.0)的新离心管中,混匀,取2 L 用于20 L 体系PCR 反应。
PCR 反应在PERKI N ELMER Gene Amp PCR Syste m 9600上进行,反应条件为94#30s,55#30s,72#1min,进行30个循环。
PCR 扩增产物用1.5%琼脂糖凝胶电泳检测。
1.6 Southern blot 分析参照陈大明等 6 报道的方法,提取转基因莴苣植株总DNA,取20 g DNA 用限制性内切酶EcoRI 酶切,0.7%琼脂糖凝胶电泳12~16h,DNA 充分分离后,转移到尼龙膜上 7。
采用Boehriger 公司地高辛标记试剂盒,操作步骤按厂商说明书进行。
将甜蛋白基因400bp PC R 产物进行标记,并用含7%SDS 和50%去离子甲酰胺杂交液进行预杂交和杂交。
杂交膜经2∃SSC 、0.1%SDS 室温洗涤两次,0.1∃SSC 、0.1%SDS 68#洗涤两次后再经抗原抗体显色反应,然后进行X 光片曝光处理。
2 结果与讨论2.1 Kan 抗性植株的获得及影响因子2.1.1 抗Kan 植株的再生 莴苣对照子叶外植体在无Kan 的MS I 培养基中再生率可达100%,当Kan 浓度达到50mg/L 时难以再生出正常芽。
经根癌农杆菌侵染的部分外植体在含Kan 培养基中首先膨大,然后在切口处形成白色愈伤组织,10d 左右在愈伤组织中间2473期 刘敬梅等:甜蛋白基因MBL II 对莴苣的遗传转化可见绿色小芽,但比对照出芽稍迟,而且每块外植体上的芽数也少于对照(对照一般7~ 8个芽/外植体,经侵染处理的则为4~5个芽/外植体)。
与农杆菌共培养后的外植体在Kan25~250mg/L的浓度梯度中,芽再生率随Kan浓度升高而降低,达到250mg/L后,芽再生困难(图2)。
待抗性芽长至1~2cm后,移入筛选生根培养基,均能100%生根。
再生植株在蛭石中炼苗10~15d后,移入土壤成活率达到100%。
2.1.2 共培养时间对Kan抗性芽分化的影响 莴苣子叶外植体经根癌农杆菌侵染后在无Kan培养基中共培养是成功转化的重要因素。
不经过共培养阶段的子叶外植体,在筛选培养基上能够在伤口处产生愈伤组织,但很难再生出芽。
这可能是由于共培养时间过短,T DNA转移不充分造成的。
而共培养7d或更长,则根癌农杆菌过度繁殖,外植体受伤严重,芽再生率迅速降低。
图3结果表明,外植体在共培养0~5d范围内,虽然随共培养时间的增加,抗性芽分化频率提高,但再生芽中假阳性芽也大幅度升高。
这些芽在筛选生根培养基上逐渐白化死去,为无效芽。
这与Sueo 等 3的报道相一致。
因此莴苣的转化以共培养3d最佳(生芽率为58.3%,白化率29%)。
试验中还发现,暗培养对转化率有很大影响。
光照下共培养的再生率仅为2.1%,而在黑暗下进行共培养的外植体的出芽率达到58.3%。
2.2 转基因莴苣的鉴定2.2.1 GU S基因在莴苣叶片中的表达 GU S检测结果表明,不同抗性芽叶片GUS 基因表达部位和表达程度各不相同。
一般情况下,GU S基因在叶片的叶脉处表达较强,叶脉呈蓝色;有些叶片GU S基因表达特别强,则全部呈深蓝色;有的叶片GUS 基因表达很弱,只有零星蓝色斑点(图4)。
统计GU S染色结果,Kan抗性芽中7.6%为GU S阳性芽。
大量非转化芽的存在,可能是由于在遗传转化过程中,子叶外植体与农杆菌共培养的同时,形成了许图2 K an浓度对莴苣抗性芽分化的影响Fig.2 E ffects of K an concentration on shootdifferentiation frequen cy图3 不同共培养时间对芽再生的影响Fig.3 Effect of duration of co cu ltureon shoot regeneration图4 抗K an再生芽叶片的GUS染色分析1.整张叶片蓝色;2,3.叶脉蓝色为主; 4.零星蓝斑。
Fig.4 GUS histochemical ass ay of leaves ofkanamycin resis tant s hoots.1.Whole leaf in blue;2, 3.M ainl y vein i n bl ue;4.Sporadic bl ue spots.248 园 艺 学 报 28卷多非转化芽原基。
这些芽原基在移入筛选培养基后,能够利用子叶中储藏的养分,发育成假阳性再生芽。
2.2.2 莴苣总DNA 的PCR 特异扩增检测 利用甜蛋白基因序列内部的一对特异引物,对GU S 阳性植株进行PCR 特异扩增,发现GUS 染色阳性植株均可扩增出特异的400bp 条带(图5)。
2.2.3 转基因莴苣植株的Southern blot 从图6结果可以看出,转基因莴苣总DNA 与MB L II 400bp 探针均有一条杂交带,而且两植株的杂交带大小不同,表明MBL II 基因已单拷贝整合到莴苣基因组DNA 中,而且整合的位置不同。
甜蛋白基因在莴苣中的表达水平,仍需做进一步的蛋白检测。
图5 转基因莴苣植株的PCR 特异扩增检测1.DL2000分子量标准;2.pLX10质粒(含MBL II基因); 3.未经转化植株;4~8.转基因植株Fig.5 PCR screening of transformed lettu ce1.D L2000D NA marker2.pLX10plas mid with M BL II gene3.Non trans formed plant 4-8.Trans formed pl ant 图6 转基因莴苣植株的Southern blot 分析1.阳性对照(含M BL II 基因的质粒pLX100.8kb 酶切片段); 2.未转基因植株;3,4.转基因植株Fig.6 Southern blot analys is of trans formed lettuce 1.Pos itve control (0.8kb fragment wi th MBL II gene);2.Non trans formed plant;3, 4.Transformed plants.参考文献:1 Richard M ,Ellen M ,Sus an S,et al.Transformation of lettuce (Lactuca sativ )a mediated by Agrobac te rium tumy fac ie ns .Plant Cell Reports,1987,6:439~4422 Curtis L S ,Power J B,De Loat A M M ,et al.Expres sion of a chi meric nitrate reductase gene in trans genic lettuce nitratee in leaves.Plant Cell Reports,1999,18:889~8963 Sueo E,Hirotaka I,Masahi ro O,et al.Induced e xpressi on of a chi meric gene cons truct in trans genic lettuce plants using tobacco pathogenesis related protien gene promoter regi on.Plant Cell Reports,1990,9:6~94 Sa muel S,Liwen X,Zhong H,et al.Recombinant s weet protien Mabinli n.PCT Patent 1997,WO 97/009455 Jeffers on R A,Kavanagh T A,Bevan M W.GUS fusion: glucuronidase as a sensi tive and vers atile gene fusion marker in hi gher plants.EMBO.J.,1987,6:3901~39076 陈大明,张上隆,金勇丰.一种木本果树基因组DN A 提取方法研究.浙江大学学报,1997,23(6):621~6247 萨姆布鲁克,弗里奇,曼斯阿蒂斯.分子克隆实验指南.金冬雁等译.北京:科学出版社,1992.475~4832493期 刘敬梅等:甜蛋白基因MBL II 对莴苣的遗传转化250 园 艺 学 报 28卷Genetic Transformation and Plant Regeneration of Lettuce with Sweet Protein Gene MBL IILiu Jingmei1,2,Chen Da ming2,and Chen Hang2(1Department o f H orticu lture,Zhejian g Un iversit y,Hangzhou310029; 2Bei jing Vegetable Research Cente r,Bei j ing 100089)Abstract:The sweet protein gene MB L II was introduced into lettuce by co culturing e xcised cotyledon explants with Agrobacterium tume faciens.The Agrobacterium strain LBA4404contained the binary vector pLX10,which carried sweet protein gene MBL II,selec tive gene NPT II and reporter gene GU S.The best transformation efficiency was obtained when explants were co cultured with Agrobacterium for3days and cultured on the selective medium containing50mg/L kanamycine (shoots regeneration rate58.3%,albino rate29%).Roots were induced in all adventitious shoots on MSII(MS+NAA0.05m g/L+Kan50mg/L+carbencillin300mg/L).The transgenic nature of regenerants were demonstrated by plant growth on medium containing kanamycine,GU S histochemi cal assay and PCR amplification.7.6%of all Kanamycine resistant shoots was GU S positive. Southern blot results confir med that the MB L II gene had been incorporated into the genome of lettuce.Key words:Sweet protein;Lettuce;Transformation广 告邮售美国芦荟、食用仙人掌苗芦荟,可盆栽观赏。