拟南芥核苷磷酸化酶基因的组织表达及其T-DNA 插入突变体鉴定
- 格式:docx
- 大小:41.19 KB
- 文档页数:7


遗传学实验报告拟南芥T-DNA插入突变体的鉴定一、实验目的:1、学习和掌握基本的植物DNA的CTAB提取法,掌握PCR、琼脂糖凝胶电泳等基本实验操作技能2、了解T-DNA插入突变体的鉴定原理,掌握其方法。
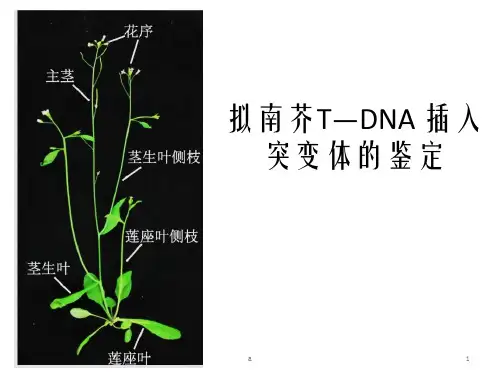
二、实验原理1、拟南芥(Arabidopsis thaliana)十字花科,植物遗传学、发育生物学和分子生物学的模式植物。
植株形态个体小,高度只有30cm左右;生长周期快,从播种到收获种子一般只需8周左右;种子多,每株可产生数千粒种子;形态特征简单,生命力强,用普通培养基就可作人工培养;遗传转化简单,转化效率高;基因组小,只有5对染色体,125MB;在2000年,拟南芥成为第一个基因组被完整测序的植物。
2、突变体突变体是遗传学研究的最重要材料。
突变体可以通过自然突变和人工诱变的方法获得。
拟南芥诱变常用方法有EMS诱变、T-DNA插入突变、激活标签。
由于T-DNA插入突变体便于对突变基因进行追踪,目前拟南芥、水稻中已经有大量的T-DNA插入突变体;SALK中心提供的拟南芥T-DNA插入突变体超过十万种。
3、T-DNA插入突变原理T-DNA,转移DNA(transferred DNA ),是根瘤农杆菌Ti质粒中的一段DNA序列,可以从农杆菌中转移并稳定整合到植物基因组。
人们将目的基因插入到经过改造的T-DNA区,借助农杆菌的感染实现外源基因向植物细胞的转移与整合,获得转基因植株。
除用于转基因以外,T-DNA插入到植物的基因中可引起基因的失活,从而产生基因敲除突变体,T-DNA大多为单拷贝插入,使其利于进行遗传分析。
4、T-DNA插入突变体PCR鉴定图 1 结果鉴定图 2 PCR引物设计三、实验材料1、材料:T-DNA插入的突变拟南芥植株;2、仪器:离心管,离心机,水浴锅,移液枪,PCR仪,电泳槽等;3、试剂:液氮,CTAB提取液,氯仿/异戊醇(24:1),无水乙醇,70%乙醇,10xTaq buffer,MgCl2,引物,琼脂糖,溴化乙锭(EB)。

拟南芥T-DNA插入突变体的鉴定09生工吴超 200900140129一、实验原理T-DNA插入法是反向遗传学研究的重要手段。
T-DNA是农杆菌的一个大质粒,长度在25kb左右。
野生型农杆菌的T-DNA上带有激素合成基因,感染植物后会导致植物细胞快速增殖形成愈伤组织,失去分化能力。
所以一般实验使用改造后的农杆菌——T-DNA中导入了卡那霉素抗性基因和抗除草剂基因。
因此在农杆菌感染植物后可用除草剂来筛选转化子。
在转化子培养到F2代出现分离后,就需要对其基因型进行鉴定。
T-DNA插入突变体鉴定方法主要有两种:三引物法和双引物法。
在本实验中使用三引物法。
三引物法的原理如图1所示,即采用三引物(LP、RP、BP)进行PCR扩增。
野生型植株目的基因的两条染色体上均未发生T-DNA插入,所以其PCR产物仅有1种,分子量约900bp(即从LP到RP);纯合突变体植株目的基因的两条染色体上均发生T-DNA插入,T-DNA本身的长度约为25kb,过长的模板会阻止目的基因特异性扩增产物的形成,所以也只能得到1种以BP与LP或RP为引物进行扩增的产物,分子量约为400-700bp;杂合突变体植株只在目的基因的一条染色体上发发生了T-DNA插入,所以PCR扩增后可同时得到两种产物。
上述3种情况的电泳结果差异明显,能有效区分不同基因型的植株。
此法优点是可同时鉴定出纯和突变体并确证T-DNA的插入情况。
图1 T-DNA插入示意图CATB,即十六烷基三甲基溴化铵,是一种离子型表面活性剂。
能溶解细胞膜和核膜蛋白,使核蛋白解聚,从而使DNA得以游离出来。
并且CATB可在高离子强度的溶液里与蛋白质和大多数多聚糖形成复合物进而形成沉淀,但不沉淀核酸。
本实验使用CATB抽提DNA。
聚合酶链式反应(Polymerase Chain Reaction,PCR)是体外核酸扩增技术。
它具有特异性高、敏感、产率高、快速、简便、重复性好、易自动化等突出优点;能在一个试管内将所要研究的目的基因或某一DNA片段于数小时内扩增至十万乃至几万倍,使肉眼能直接观察和判断。
T-DNA插入突变体的鉴定时明辉同组者:薛敏学号:201000220069摘要 Ti质粒是上有一段特殊的DNA区段,当农杆菌侵染植物细胞时,该DNA区段能自发转移进植物细胞,并插入植物染色体DNA中。
所以Ti质粒上的这一段能转移的DNA被叫做T-DNA。
将感兴趣的基因改造插入到T-DNA区段中,通过农杆菌侵染植物细胞,实现外源基因对植物的遗传转化,得到含有突变的植株。
通过本实验,我们将学习如何用PCR的方法检测所得植株是否为T-DNA的插入突变体。
1.引言T-DNA作为一种实验常用的遗传转化方法,在插入突变过程中,插入到植物染色体上的位置是随机的。
如果T-DNA插入进某个功能基因的内部,特别是插入到外显子区,将造成基因功能的丧失。
所以利用农杆菌Ti质粒转化植物细胞,是获得植物突变体的一种重要方法。
农杆菌Ti质粒转化植物细胞后,在获得的后代分离群体中,有T-DNA插入的纯合突变体,杂合突变体,和野生型。
在突变体研究中,需要的材料是纯合突变体,所以必须从分离群体中将纯合突变体鉴定出来。
本次实验中,采用液CTAB(或者TSP法)提取拟南芥植株的DNA,然后PCR将所获DNA 扩增,在之后采用琼脂糖凝胶电泳技术,分离处长度不一的DNA带,以确定样品是否为T-DNA 插入突变纯和体。
PCR(Polymerase Chain Reaction),即聚合酶链式反应是体外核算扩增技术,具有特异、敏感、产率高、快速、简便、重复性好、易自动化等突出优点;能在一个试管内将所要研究的目的基因或某一DNA片段于数小时内扩增至十万乃至百万倍,使肉眼能直接观察和判断。
(PCR基本原理如右图)3-基团,在pH8.0 BufferDNA含有PO4(本实验中为TAE)中带负电, 在电场中向正极移动。
自由电泳时,由于不同大小的DNA片段的电荷密度大致相同,各核酸分子难以分开;选用适当浓度的琼脂糖凝胶作为支持物,使之具备一定的孔径,即可发挥分子筛效应,使大小不同的核酸片段迁移率出现较大差异,达到分离的目的;同样条件对Marker电泳;起到鉴定的作用。

拟南芥T-DNA插入突变纯合体的鉴定余振洋(高山山、潘红芳)、09级生技1班、200900140156、2011/12/14摘要本实验通过CTAB法提取目的拟南芥的DNA,再用三引物法PCR扩增所需的目的基因后,用电泳检测该拟南芥是否为转基因的拟南芥,并判断其是纯合突变还是杂合突变。
关键词拟南芥;T-DNA;突变纯和体1.引言T-DNA是根癌农杆菌Ti质粒上的一段DNA序列,它能稳定地整合到植物基因组中并稳定地表达。
T—DNA在植物中一般都以低拷贝插入,多为单拷贝。
单拷贝T-DNA一旦整合到植物基因组中,就会表现出孟德尔遗传特性,在后代中长期稳定表达,且插入后不再移动,便于保存。
T—DNA插入突变在反向遗传学和功能基因组学研究中发挥着重要作用。
,T—DNA插入突变能方便地进行正向和反向遗传学研究,因而受到重视。
同时,基因组测序工作的完成使得从位点到表型的反向遗传学研究成为可能,从而使通过T—DNA插入技术构建突变体来研究功能的反向遗传学技术逐渐取代了传统的化学诱变、图位克隆等技术。
借助于农杆菌介导的遗传转化技术,T—DNA插入技术已被广泛应用于拟南芥等模式植物的突变体库构建中。
以T—DNA作为插入元件,不但能破坏插入位点基因的功能,而且能通过插入产生的功能缺失突变体的表型及生化特征的变化,为该基因的研究提供有用的线索。
由于插入的T—DNA序列是已知的,因此可以通过已知的外源基因序列,利用反向PCR、TAIL-PCR、质粒挽救等方法对突变基因进行克隆和序列分析,并对比突变的表型研究基因的功能。
还可以利用扩增出的插入位点的侧翼序列,建立侧翼序列数据库,对基因进行更全面的分析。
由此可见,T—DNA 插入标签技术已成为发现新基因、鉴定基因功能的一种重要手段。
CTAB法提取植物叶片中的DNA是我们常用的方法。
通常采用机械研磨的方法破碎植物的组织和细胞,由于植物细胞匀浆含有多种酶类(尤其是氧化酶类)对DNA的抽提产生不利的影响,在抽提缓冲液中需加入抗氧化剂或强还原剂(如巯基乙醇)以降低这些酶类的活性。


突变基因的拟南芥实验研究拟南芥(Arabidopsis thaliana)是一种模式植物,在生物学研究中发挥着重要的作用。
它的基因组序列已经被完整解读,并且其外观简单、生长周期短等特点,使得其成为基因功能研究的最佳实验材料。
突变基因是指由于DNA序列的变异,造成突变的基因。
拟南芥的突变基因贡献了大量关于植物发育与繁殖等方面的科学研究成果。
突变基因的发现突变基因的发现可以通过自然突变和诱导突变两种途径实现。
自然突变是指在自然条件下,由于DNA杂交、突变等自然因素,使得基因产生突变。
而诱导突变,则需要使用特殊的化学试剂或是电磁辐射等手段对DNA进行干预,从而获得突变基因。
诱导突变的方法目前,诱导突变的方法主要有以下几种:1. EMS法EMS是Ethyl methanesulfonate的缩写,是一种碱基化剂,能够导致DNA中的鸟嘌呤碱基突变。
通过对拟南芥幼苗进行EMS浸泡处理,可以获得大量的突变体。
2. Gamma射线法Gamma射线是一种高能辐射,能够直接影响DNA分子结构,从而导致基因突变。
使用Gamma射线进行诱导突变,可以获得不同类型的突变体,包括缺失、插入、点突变等。
3. T-DNA插入法T-DNA是一种细菌表现元(bacterial virulence factor),广泛存在于土壤中的根际细菌Agrobacterium tumefaciens中。
因为T-DNA能够与植物基因组发生同源重组,因此可以通过向植物中转化Agrobacterium,从而将T-DNA插入到植物基因组中,诱导基因突变。
突变基因的分析方法了解突变基因的表达情况,可以通过基因表达谱、荧光素酶检测、Northern blotting、Western blotting等多种方法实现。
其中基因表达谱是最常用的一种方法,能够快速、准确地检测基因的表达情况。
拟南芥突变基因的研究拟南芥作为模式植物,其突变基因的研究对于植物的发育和繁殖等方面具有重要的意义,以下是一些拟南芥突变基因的研究案例。


姓名系年级学号日期科目遗传学实验题目模式植物拟南芥T-DNA插入突变体的鉴定模式植物拟南芥T-DNA插入突变体的鉴定摘要:农杆菌Ti质粒转化植物细胞后,在获得的后代分离群体中,有T-DNA插入的纯合突变体,杂合突变体,和野生型。
在突变体研究中,需要的材料是纯合突变体。
本次实验意在对模式植物拟南芥T-DNA插入突变体进行鉴定。
实验中用到的主要实验方法有:CTAB法提取拟南芥基因组DNA、PCR扩增目的基因片段、琼脂糖凝胶电泳分离核酸。
PCR技术的基本原理类似于DNA的天然复制过程。
PCR由变性--退火--延伸三个基本反应步骤构成。
每完成一个循环需2~4分钟,2~3小时就能将待扩增目的基因扩增放大几百万倍。
琼脂糖凝胶电泳是用琼脂糖作支持介质的一种电泳方法。
它兼有“分子筛”和“电泳”的双重作用。
带电颗粒的分离不仅取决于净电荷的性质和数量,而且还取决于分子大小,这就大大提高了分辨能力。
DNA经EB染色,EB可与核酸结合,在紫外光激发下产生荧光。
现广泛应用于核酸的研究中。
引言Ti质粒是土壤农杆菌的天然质粒,该质粒上有一段特殊的DNA区段,当农杆菌侵染植物细胞时,该DNA区段能自发转移进植物细胞,并插入植物染色体DNA 中。
所以Ti质粒上的这一段能转移的DNA被叫做T-DNA。
将Ti质粒进行改造,将感兴趣的基因放进T-DNA区段中,通过农杆菌侵染植物细胞,实现外源基因对植物的遗传转化。
T-DNA插入基因内部导致基因突变:T-DNA插入到植物染色体上的位置是随机的。
如果T-DNA插入进某个功能基因的内部,特别是插入到外显子区,将造成基因功能的丧失。
所以利用农杆菌Ti质粒转化植物细胞,是获得植物突变体的一种重要方法。
农杆菌能在自然条件下感染双子叶植物和裸子植物,而对大多数单子叶植物没有感染能力。
这使农杆菌Ti 质粒转化系统的应用范围受到了一定的限制。
反向遗传学研究的首要条件是获得基因敲出突变体,建立可靠、有效、方便的T-DNA插入突变体的鉴定方法。

拟南芥T-DNA插入突变体的鉴定摘要:我们用CTAB法提取拟南芥的T-DNA插入突变体的DNA,然后用三引物法进行PCR和琼脂糖凝胶电泳来判断其为突变纯合体还是突变杂合体。
通过这次实验,我们掌握了如何来判断纯和突变和杂合突变。
关键字:拟南芥 T-DNA插入突变突变体的鉴定前言:拟南芥拟南芥是十字花科的植物,它是植物遗传学、发育生物学和分子生物学的模式植物,其具有以下这些特点:①植株形态个体小,高度只有30cm左右;②生长周期快,从播种到收获种子一般只需8周左右;③种子多,每株可产生数千粒种子;④形态特征简单,生命力强,用普通培养基就可作人工培养;⑤遗传转化简单,转化效率高;⑥基因组小,只有5对染色体,125MB;⑦在2000年,拟南芥成为第一个基因组被完整测序的植物。
突变体突变体在植物基因分离及遗传学研究的最重要材料,通过自然突变或者人工诱变同源重组、基因沉默以及插入突变等方法都可以用来构建突变体,人工诱变是指利用物理因素(X射线,Y射线,紫外线,激光等)或化学诱变(如亚硝酸,硫酸二乙酯)来处理生物,使生物发生基因突变,这种方法可提高突变率,创造人类需要的变异类型。
目前,人工诱变拟南芥常用的方法有EMS诱变、T-DNA 插入突变、激活标签等。
T-DNA插入突变Ti质粒是土壤农杆菌的天然质粒,该质粒上有一段特殊的DNA区段,当农杆菌侵染植物细胞时,该DNA区段能自发转移,插入植物染色体DNA中,Ti质粒上的这一段能转移的DNA被叫做T-DNA。
人们根据这一现象,将Ti质粒进行改造,将感兴趣的基因放进T-DNA区段中,通过农杆菌侵染植物细胞,实现外源基因对植物的遗传转化。
T-DNA插入到植物染色体上的什么位置,是随机的。
如果T-DNA插入某个功能基因的内部,特别是插入到外显子区,将造成基因功能的丧失。
所以利用农杆菌Ti质粒转化植物细胞,是获得植物突变体的一种重要方法。
T—DNA插入突变最大的用处是构建突变体库,在此基础上构建侧翼序列库;目前在拟南芥中已经建立了接近饱和的T—DNA插入突变体库,该突变体库包含超过225 000个独立的T—DNA插入株系,在预测的29 454个基因中有21 700个基因发生了插入突变[6]。
拟南芥谷氨酰胺磷酸核糖焦磷酸转酰胺酶缺陷型突变体的特征描述使用基于转基因的筛选,我们之前分离出一些蛋白质输入叶绿体有缺陷的拟南芥突变体。
定位克隆其中的一个位点,CIA1,显示出CIA1编码谷氨酰胺磷酸核糖焦磷酸转酰胺酶2(A Tase2),是负责嘌呤从头合成第一个关键步骤的三个同工酶之一。
CIA1突变体有正常的绿色子叶,但却有微小和白化或淡绿色的马赛克叶子。
添加AMP而不是细胞分裂素或NADH 到植物液体培养基,部分补充了突变体的表现型。
ATase1和ATase2都位于叶绿体上。
ATase1的过量表达充分补充了ATase2缺陷型的表表型。
也获得了一个T-DNA插入敲除A Tase1的突变体。
突变体与野生型无法区分。
一个敲除cia1/ATase1的双突变体与cia1有相同的表型,说明ATase1和ATase2之间至少有一部分基因冗余。
Cia1突变体的特征描述显示出突变体叶子有略小的细胞尺寸但只有野生型叶子细胞数量的一半。
这一表型确定嘌呤从头合成在细胞分裂中的作用。
Cia1突变体分离出的叶绿体输入蛋白质的效率比野生型叶绿体少50%。
添加ATP和GTP到分离出的突变体叶绿体不能恢复输入效率。
我们可以得出结论嘌呤从头合成不仅对细胞分裂重要,而且对叶绿体的生物合成也很重要。
嘌呤环的从头合成是植物生长和发育所必需的。
主要的产物,AMP和GMP,都是DNA和RNA的结构单位。
AMP,当转化为ATP后,是多种细胞过程的主要能量来源。
一些重要的辅酶,如NAD和FAD,也来自相同的途径。
在热带固氮豆科植物的根瘤中,如大豆和豇豆,这一途径在初级氮代谢中也起到了主导作用。
嘌呤生物合成途径中酶的活性与其他组织相比大大增强。
因此,大部分植物嘌呤生物合成的研究都使用这些豆科植物的根瘤作为材料,集中于嘌呤生物合成在氮同化中的功能。
研究嘌呤生物合成在正常植物生理或非固氮植物中的作用一直相对较少。
植物细胞中嘌呤生物合成的位置依然有争议,途经中的植物酶与在大肠杆菌中表达的相似,除了每种植物酶有一个被推测为作为细胞器定位信号的N-末端延长。
拟南芥基因突变体研究及其分子机理分析拟南芥是一种重要的模式植物,在基因突变体研究中发挥着重要的作用。
本文将从拟南芥基因突变体的定义、研究方法、重要性以及其分子机理等方面进行探讨和分析。
一、拟南芥基因突变体定义及研究方法基因突变体是指在基因序列中发生变异的个体,与野生型(WT)相比,基因突变体的表型有明显的差异。
拟南芥基因突变体是以拟南芥(Arabidopsis thaliana)为材料的基因突变研究。
它具有许多优秀的特性,如短生命周期、小型体型、遗传变异多样化和基因功能高度保守等。
目前,拟南芥基因突变体的研究方法主要分为化学诱变、遗传转化和基因编辑。
其中,化学诱变是通过化学物质引起基因突变,常用的化学物质有Ethyl methane-sulfonate (EMS)和Sodium azide (NaN3)等。
遗传转化是利用外源DNA片段引入目标基因,达到基因敲入/敲除的目的。
基因编辑则是指利用CRISPR/Cas9等基因编辑技术对目标基因进行精准的编辑,从而实现目的基因的敲入/敲除。
这些方法的优缺点各有不同,可以根据实验目的和条件选择适宜的研究方法。
二、拟南芥基因突变体的重要性拟南芥基因突变体研究有着重要的科研意义和现实意义。
首先,拟南芥是植物领域中最具代表性的模式植物之一,研究拟南芥基因突变体可以为解析生物分子机理和育种提供重要的理论依据。
其次,拟南芥基因突变体的发现对研究复杂性状、生长发育和环境响应等现象起着重要作用,同时也对人类生命健康、农业生产、环境保护等方面具有深远的影响。
三、拟南芥基因突变体分子机理分析拟南芥基因突变体分子机理分析是对基因突变体的表型变化进行解析的过程。
在基因突变体的研究中,通常采用遗传学、生物化学和分子生物学等多种技术手段进行深入研究。
遗传学方法主要包括染色体显微镜观察、连锁分析、基因定位和基因组学分析等。
在染色体显微镜观察中,通过观察细胞染色体数目、形状、大小和染色体带的特点,可以发现染色体异常和染色体突变。
拟南芥论文:拟南芥细胞分裂素糖基转移酶 O-糖基化修饰突变体过表达体基因表达模式亚细胞定位【中文摘要】细胞分裂素是一类较活跃的植物激素,具有广泛的生物学活性,不仅在调节细胞增殖和分裂中起着至关紧要的作用,并且控制着植物生长发育的各个阶段,如延缓衰老,控制芽/根平衡,营养信号转导,农作物增产等。
细胞分裂素的糖基化是细胞分裂素的重要修饰方式,细胞分裂素的糖基化主要有O-糖基化和N-糖基化两种形式。
一般来说,细胞分裂素的N-糖基化使该激素永久失活,激素的活性无法逆转;而O-糖基化使激素暂时失活,在一定条件下可通过去糖基化而恢复细胞分裂素活性。
细胞分裂素的两种糖基化修饰对细胞分裂素在植物体内的动态平衡起着重要作用。
细胞分裂素的糖基转移酶是催化其发生糖基化反应的酶。
在以前的研究中,通过体外生化实验从拟南芥中已经鉴定细胞分裂素的2个N-糖基转移酶基因和3个O-糖基转移酶基因,但这些糖基转移酶在植物体内的作用和生理学意义仍不清楚。
本论文选取一个活性较强的O-糖基转移酶基因UGT85A1,以模式植物拟南芥和烟草为实验材料,分析了该基因的突变体和过表达体在植株形态、生理等方面的特征,同时利用GUS报告系统分析了UGT85A1基因表达的组织特异性、发育阶段特异性,利用GFP报告系统分析了糖基转移酶UGT85A1在细胞内的定位。
研究结果为进一步了解细胞分裂素O-糖基化修饰的生理学意义奠定了一定的基础。
本论文的主要研究结果有以下几个方面:1.鉴定了拟南芥突变体纯合体,并与野生型拟南芥进行比较。
正常生长条件下,拟南芥突变体与野生型无明显表型差异。
在添加外源细胞分裂素的培养基上,突变体在根长、侧根数目、叶绿素含量、细胞分裂素及糖苷含量等形态和生理指标上与野生型相比也未出现显著差异。
2.构建了植物过表达载体35Spro::UGT85A1,并通过农杆菌介导的基因转化方法在烟草和拟南芥中过表达,比较了过表达体与野生型植株的形态与生理学特征。
拟南芥突变体的鉴定摘要在真核细胞的细胞核中,基因组DNA缠绕在组蛋白上,形成核小体的结构。
核心组蛋白的末端会发生一系列翻译后水平上的共价修饰,主要包括甲基化、乙酰化、磷酸化、泛素化和ADP-核糖基化等。
这些共价修饰在染色质的结构与功能及基因的转录调控方面起着重要的作用。
组蛋白的甲基化发生在赖氨酸和精氨酸位点,这些位点的甲基化参与异染色质形成,X染色体失活和基因转录的激活与沉默等许多重要的生物学功能。
早先认为组蛋白上的甲基化是不可逆的。
然而近年来在哺乳动物和酵母中相继发现了许多组蛋白去甲基化酶。
其中最大的一个基因家族是包含有JMJC结构域的组蛋白去甲基化酶家族。
在植物中还没有报道有组蛋白去甲基化酶。
我们通过基因组注释以及序列比对从拟南芥中鉴定出21个编码包含有JMJC结构域蛋白的基因。
并从SALK突变体库中筛选鉴定了相应的T-DNA插入突变体。
利用分子标记,我们筛选出了37个纯合T-DNA插入株系。
给我们以后分析这些基因在植物生长发育中所起的作用打下了坚实的基础。
关键词组蛋白密码, 甲基化,去甲基化,T-DNA插入突变体1 背景介绍1.1 “组蛋白密码”理论与表观遗传学在真核细胞的细胞核中,基因组DNA与组蛋白和非组蛋白相互结合,构成遗传信息的物质载体——染色质。
染色质由其基本单位核小体重复连接而成,每个核小体包含一个组蛋白八聚体(H2A/H2B-H3/H4-H3/H4-H2A/H2B)和围绕其上的两圈超螺旋DNA(146KB),核小体之间通过不同长度的超螺旋DNA和H1组蛋白前后相接。
组蛋白八聚体的成员作为核小体的基本组成元件,在所有真核生物中高度保守。
尽管核心组蛋白均具有一个三螺旋的结构严整的组蛋白折叠区,其两个末端却没有形成固定结构。
然而,这些末端却会发生一系列翻译后水平上的共价修饰,主要包括甲基化、乙酰化、磷酸化、泛素化和ADP-核糖基化等。
这些共价修饰在染色质的结构与功能及基因的转录调控方面起着重要的作用(1,2)。
拟南芥核苷磷酸化酶基因的组织表达及其T-DNA 插入突变体鉴定徐文晶;程玉祥【摘要】To explore information and physiological function of nucleoside phosphorylase genes,by using PCR method,the authors examined gene expression of nucleoside phosphorylase in different tissues of model plant arabidopsis, and further screened T-DNA-inserted mutants of these genes. Results:Transcriptional expression of At4g28940 and At4g28940 is high in root tissues ,and slight transcript levels are detected in other tissues ,while At4g24350 gene expression is low in various tissues .After the identification of T-DNA insertion and transcriptional levels of thegenes ,the knockout mutants of At4 g28940 and At4 g24350 have been attained in this study .%为探明植物核苷磷酸化酶家族基因信息及生理功能,采用 PCR 扩增的方法,研究了模式植物拟南芥核苷磷酸化酶家族基因在不同组织中的表达,并对其核苷磷酸化酶基因 T-DNA 插入突变体进行鉴定。
结果表明:At4g24340和 At4g28940基因在根组织中大量转录表达,其他组织均微量检测到转录表达;At4g24350在各个组织中均有少量转录表达;经 T-DNA 插入结合转录表达鉴定,At4g28940和 At 4g24350基因为敲除突变体。
【期刊名称】《贵州农业科学》【年(卷),期】2015(000)001【总页数】4页(P16-19)【关键词】拟南芥;核苷磷酸化酶;突变体【作者】徐文晶;程玉祥【作者单位】东北林业大学林木遗传育种国家重点实验室,黑龙江哈尔滨150040;东北林业大学林木遗传育种国家重点实验室,黑龙江哈尔滨 150040【正文语种】中文【中图分类】S503.52核苷磷酸化酶(Nucleoside phosphorylase,NP)是核苷酸补救合成途径的代谢酶[1]。
嘌呤核苷磷酸化酶(PNP,EC 2.4.2.1)是核苷磷酸化酶家族的一个小亚家族,在无机磷酸存在下其可逆地催化核苷或脱氧核苷糖苷键断裂,生成嘌呤碱和一磷酸核糖或脱氧核糖[2-3]。
嘌呤核苷磷酸化酶在医学上的研究报道较多,其缺乏症患者通常存在严重的免疫缺陷和神经系统功能障碍,还会引起血内尿酸不足,家族性痉挛性截瘫,脑血管病变导致的中风,多灶性白质脑病等[4-8]。
植物中也存在核苷磷酸化酶,拟南芥叶绿体多核苷磷酸化酶(cpPNPase)参与r RNA和m RNA的3′末端的成熟[9-10]。
然而,对于植物中核苷磷酸化酶生理功能人们了解甚少。
最近有研究报道[11],毛果杨基因组中存在13个NP-like基因,可能是一类营养储存蛋白。
基于笔者实验室前期研究与分析推断,毛果杨部分核苷磷酸化酶很可能和次生细胞壁代谢与修饰相关。
然而,在杨树中无法获得这些基因敲除的多重突变体用于其遗传功能鉴定。
对毛果杨13个NP-like基因在拟南芥数据库BLASTP中同源检索,仅得到At4g24340、At4g24350和At4g28940同源对应基因,且目前未见这些基因的T-DNA插入突变体的相关报道。
为此,笔者等选择易于获得家族基因多重突变体的模式植物拟南芥,分析其At4g24340、At4g24350和At4g28940基因的组织表达模式,并筛选、鉴定其T-DNA插入突变体,以期为下一步研究植物的核苷磷酸化酶的功能奠定基础。
1.1 试验材料1.1.1 拟南芥野生型拟南芥均为Columbia型,购自ABRC(Ohio State University,USA)。
1.1.2 T-DNA插入株系 At4g28940基因的T-DNA插入株系为SALK_149317 C和SALK_018193 C,At4g24350基因的T-DNA插入株系为SALK_072656 C,At4g28940基因的T-DNA插入株系为CS491284,均购自ABRC(Ohio State University,USA)。
1.1.3 主要试剂植物基因组DNA快速提取试剂盒、pBIOZOL Reagent和Silica Bead DNA Gel Extraction Kit,分别购自百泰克、BioFlux和Fermentas公司;pMD18-T载体、Taq DNA聚合酶和PrimeScript RT reagent Kit With gDNA反转录试剂盒,购于TaKaRa公司;引物由Invitrogen公司合成。
1.2 植物基因组 DNA、总 RNA的提取及cDNA的合成1) 植物基因组DNA的提取。
参照试剂盒说明书进行。
2) 植物总RNA提取及cDNA合成。
植物材料经过液氮速冻、研碎至粉末,用4℃预冷的pBIOZOL试剂悬浮粉末后移入1.5 mL离心管,具体步骤参照pBIOZOL Reagent说明书进行。
总cDNA合成采用Primescript RT reagent Kit with gDNA eraser试剂盒。
1.3 引物设计序列及PCR扩增SALK和CS株系插入T-DNA左臂端鉴定引物分别为LBb1:5’-GCGTGGACCGCTTGCTGCAACT-3’和LBb3:5’-TAGCATCTGAATTTCATAACCAATCTCGATACAC-3’。
基因引物为SALK_149317C-LP:5’-TCATACCCTAAGGTTTACTGAGT-3’和RP:5’-TATTTTGGGGTCAACTTACTCAC-3’;SALK_018193C-LP:5’-GATAAATAATTGCTTCTCCTTCA-3’和RP:5’-TGAATCACAATGTTAGAAACGTG-3’;SALK_072656C-LP:5’-GAGCTATACAACATACTGATTAG-3’和RP:TTGTCACGGGTTCGTTGATATGA;CS491284-LP:5’-GCCATGCTTATCACGTGTGTATT-3’和RP:5’-CAGACTTAAGCTATAGAACAACC-3’。
基因转录表达半定量分析的引物为At4g24340-LP∶5’-GGCTTCCATGGATCACAATCATT-3’和RP∶ 5’-AAGGATGCGTAGGATCGAGTCT-3’;At4g24350-LP: 5’-GTCTAGAGACGAGAAGCACGTG-3’和RP:5’-CAATGACCGTCACGAGACCGAT-3’;At4g28940-LP: 5’-GATTGGCGACGTCACTATTCCT-3’和RP: 5’-GCGTGCAAGTTTCAAGTAGGAC-3’。
PCR扩增为20 μL体系:13 μL水,2 μL dNTP (2 mM),2μL 10×Buffer,1.0μL Taq DNA聚合酶,上下游引物各0.5 μL和1.0 μL模板。
PCR反应程序:预变性95℃ 3 min,变性95℃ 30 s,转录表达引物及T-DNA鉴定引物退火温度分别为59℃和62℃,32次循环。
1.4 At4g24340、At4g24350和At4g24340基因的组织表达特性及T-DNA插入突变体鉴定1) 组织表达特性。
提取野生型拟南芥不同组织的总RNA,以合成的总cDNA为模板进行定量PCR扩增。
2) 插入突变体鉴定。
提取待鉴定的植株基因组DNA作为模板,分别使用LP和RP、LP和LBb1(或LBb3)、LBb1(或LBb3)和RP 3对引物进行PCR扩增,进行T-DNA插入突变体的鉴定;将上述T-DNA引物与基因引物PCR扩增的片段从琼脂糖中回收,连接到pMD18-T载体上,选择有插入片段的克隆进行测序,以确定T-DNA的插入位置。
3) 插入纯合突变体的目标基因转录分析。
提取待鉴定植株的总RNA,分别合成总cDNA,再进行半定量RT-PCR扩增。
2.1 At4g24340、At4g24350和At4g24340基因的组织表达从图1看出,At4g28940和At4g24340基因在根组织中大量转录表达,其他组织均微量检测到转录表达;At4g24350在各个组织中均有少量转录表达(图1)。
表明,At4g28940和At4g24340基因在根组织中特异地高丰度表达。
2.2 At4g24340、At4g24350和At4g28940基因T-DNA插入突变体的鉴定从图2可知,在SALK_149317C的2个植株中,基因引物均未扩增出目标带,提示被T-DNA所插入,进一步用基因引物与T-DNA引物扩增出目标带,表明T-DNA插入在该基因的区域,并且这2个植株均是T-DNA插入纯合体(图2a)。
同理,SALK_018193 C、SALK_072656 C和CS491284株系的6个植株都是T-DNA插入纯合体(图2b-d)。
从图3看出,经过测序,SALK_149317 C为反向T-DNA插入突变体,SALK_018193C为正向T-DNA插入突变体,插入位点分别在At4g24340第1内含子113 bp处和第5内含子27 bp处;SALK_072656C则为反向T-DNA插入,插入位点在At4g24350第5外显子118 bp处;CS491284为反向T-DNA插入突变体,插入位点在At4g28940的5’UTR上,距起始密码子上游54 bp处。
2.3 T-DNA插入纯合突变体的目标基因转录从图4可知,在At4g28940基因T-DNA插入纯合体SALK_149317C和SALK_018193C中,未检测到该基因的转录,而野生型则正常转录表达。
表明,SALK_149317C 和SALK_018193C是At4g28940基因的敲除突变体,分别命名为at 4g24340-1 和at4g24340-2。
同样,SALK_072656C是At4g24350基因的敲除突变体,命名为at4g24340-1。