造血干细胞的微环境
骨髓微环境中的一些转录因子和蛋白分子对 造血干细胞的移行、增殖和分化起了调控作用。
干细胞因子
与干细胞表面的 C-kit 受体结合 GATA-2对造血前体细胞的增殖与存活是关键的 加强细胞的粘附作用 GATA-1调节红系、巨核细胞等的存活、增殖 参与维持干细胞数目的恒定 和成熟
PU1的表达可促使造血干细胞子代前体细胞进 一步分化产生淋巴系, 而且间充质干细胞相对容易在体外被诱导 分化为成骨细胞和软骨细胞,所以间充质 干细胞在骨损伤,先天骨组织畸形的治疗 方面将有广泛的应用。
间充质干细胞
间充质干细胞的来源
来源于发育早期的中胚层 和外胚层。 MSC最初在骨髓中发现。
具有多向分化潜能,理想
的种子细胞。
间充质干细胞的形态
在个体出生后, 间充质干细胞附于 骨髓窦的内腔面, 形成一种包埋于窦 状网络中的三维细 胞网络。
光学显微镜
扫描电镜
间充质干细胞的表面标志
SH2、SH3、CD29、CD44、CD71、CD90、 CD106、CD120a、及CD124等
成体干细胞的概念
存在于成年个体的许多组织和器官
特点
修复 再生 自我更新
使组织和器官保持生长和衰退的 动态平衡。
成体干细胞的可塑性
成体干细胞可以突破其 “发育限制性”,跨系, 甚至跨胚层分化为其他类 型组织细胞
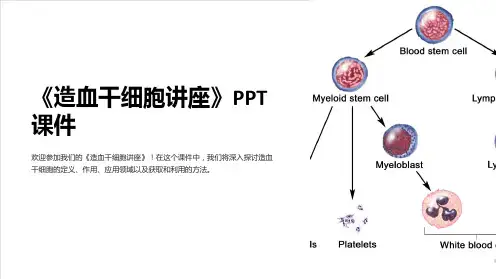
造血干细胞起源
(hemopoietic stem cell,HSC)是存 在于造血组织中的一群原始造血细 胞,它不是组织固定细胞,可存在 于造血组织及血液中。造血干细胞 在人胚胎2周时可出现于卵黄囊, 第4周开始转移至胚肝,妊娠5个月 后,骨髓开始造血,出生后骨髓成 为干细胞的主要来源。在造血组织 中,所占比例甚少,如在小鼠骨髓 中105核细胞中的有10个,在脾中 105有核细胞中只有0.2个。