低温逆境对黄瓜生长发育的影响
- 格式:doc
- 大小:29.50 KB
- 文档页数:6

低温和高温胁迫对植物生长的影响和机制植物生长对温度的适应性是快速适应其环境的关键。
当植物生长在低温或高温条件下时,植物的生长和发育将遭受一定程度的损害,严重的情况下植株可能无法生存。
因此,深入研究低温和高温胁迫对植物生长的影响和机制具有极其重要的理论和实际意义。
低温对植物生长的影响低温是指环境温度低于植物的生理和生化适宜温度范围。
植物在低温下的生理和生化过程会发生变化,最终导致它们的生长和发育受到影响。
低温对植物的生长影响因植物种类和低温程度而异,普遍影响包括:成熟期延长、代谢率减缓、植物叶片色泽变浅,在严重情况下,低温还可能导致植物死亡。
低温引起植物叶片失绿是由于低温导致合成叶绿素的过程受到抑制,同时同时植物叶片中的叶绿素降解程度增加。
此外,低温条件下植物膜的渗透率会发生变化,膜脂质会变得更加活性,组成会发生变化,近而导致对温度的响应和适应程度下降。
低温胁迫引起的代谢变化具有复杂的机制。
低温胁迫会通过调节蛋白质的翻译和转录过程,调控与代谢相关的基因,从而改变植物代谢的速率。
以水稻为例,当其生长在低温条件下时,能促进游离氨基酸的积累,这是由于低温会抑制氨基酸脱羧酶的活性,从而导致氨基酸的代谢路转向游离氨基酸的合成。
低温会增加植物的生物碱含量。
生物碱是一类富含氮或吡咯环结构化合物的次级代谢物。
除了丰富的生理功能,某些生物碱如藜芦碱还具有抗氧化作用,可以有效地防止氧化应激所导致的细胞和组织损伤。
生物碱的积累与低温胁迫的响应机制密切相关。
高温对植物生长的影响高温是指环境温度高于植物的生理和生化适宜温度范围。
高温胁迫会对植物产生严重的影响,包括光合作用的抑制、代谢率加速以及产生氧化损伤等。
生物膜的结构在高温下会发生一定的改变,使得细胞膜的通透性增强,导致ATP产生减少,卡诺循环下降,最终导致光合作用的抑制。
高温引起的氧化损伤是由于高温引起的氧化应激增加导致的。
氧化应激的产生主要是由于细胞中的ROS(反应性氧气物质)含量增加,过量ROS可导致蛋白质、核酸和脂类的氧化性损伤。

霜降对蔬菜生产的影响霜降是二十四节气中的第十八个节气,标志着寒冷的冬季即将来临。
在霜降时节,气温骤降,霜期来临,这对蔬菜生产产生了一定的影响。
本文将探讨霜降对蔬菜生产的影响,并提出对策应对这些影响。
一、土壤温度下降,影响蔬菜生长霜降时节,气温骤降,土壤温度也会相应下降。
土壤温度是蔬菜生长的重要因素之一,土壤温度过低会抑制蔬菜的生长和发育。
特别是一些热爱温暖环境的蔬菜,如番茄、辣椒等,对低温更为敏感。
霜降时节的低温会对这些蔬菜的生长产生不利影响,导致生长缓慢、产量减少。
应对策略:1. 提前做好保温措施:霜降前应在蔬菜种植区域进行地面覆盖,可使用塑料薄膜或草帘等材料进行保温,减少土壤温度的下降。
2. 选用耐寒蔬菜品种:在霜降时节选择适应低温的蔬菜品种进行种植,如冬季萝卜、冬瓜等耐寒蔬菜品种,能够更好地适应低温环境,保持一定的生长活力。
二、霜冻严重影响蔬菜品质霜降时节,霜冻现象较为常见,尤其是在寒冷地区。
霜冻会对蔬菜的品质产生严重的影响。
一方面,霜冻会引起蔬菜细胞水分结冰,导致细胞膜破裂,使蔬菜组织软化,容易腐烂;另一方面,霜冻还会破坏蔬菜中的维生素和其他营养物质,使蔬菜变得无营养、无口感。
应对策略:1. 提前采收:在霜降来临之前,尽可能提前采收蔬菜,避免受到霜冻的影响。
及时采收的蔬菜品质更好,口感更佳。
2. 增加保护措施:对于霜降来临之前无法及时采收的蔬菜,可以采取措施进行保护。
例如,搭建塑料棚或使用冻土覆盖的方法,减少霜冻对蔬菜的直接影响。
3. 合理储存:采收后的蔬菜应储存在冷藏室或其他低温环境中,以延长其保鲜期,减少品质的损失。
三、霜降促使蔬菜休眠进入生理阶段霜降时节,气温骤降,光照时间减少,这使得一些蔬菜进入休眠期,停止生长。
休眠期是蔬菜的生理阶段,蔬菜在这个阶段减缓代谢运作,生长速度大幅度下降。
对于寒冷地区的蔬菜生产来说,霜降时节的蔬菜进入休眠期是不可避免的。
应对策略:1. 调整种植计划:在霜降之前,蔬菜栽培者可以根据天气预报的情况提前进行蔬菜的收获,避免大规模进入休眠期的蔬菜产生过多的影响。

不同的温度胁迫对黄瓜抗冷性影响的初步探讨【摘要】本文研究了两个抗冷性不同的黄瓜幼苗经过6℃,15℃,常温处理3天后(常温作为对照),立即测定株高和4项生理生化指标,探讨了这些数据的成因,讨论了不同温度胁迫对黄瓜幼苗抗冷性的影响。
【关键词】温度;黄瓜;抗冷性很多热带和亚热带的作物不能忍受0-10℃低温,0℃以上低温对植物所造成的危害叫冷害[1]。
黄瓜是起源于热带雨林地区的典型的冷敏型草本植物,易发生冷害,其植株所有组织以及果实都对低温敏感,生长最适宜的温度范围狭窄,通常10-12℃以下黄瓜的生理代谢活动失调,生长缓慢或停止发育,5℃以下就难以生存[2]。
黄瓜作为北方的反季节生产的主栽蔬菜,低温冷害已成为其正常生长的主要限制因子,如何提高黄瓜的抗冷性在生产上具有重要意义[2,4],对于黄瓜的抗冷性研究,从种子发芽、苗期、开花、果实等生长时期均有报道,但研究范围狭窄,还需广大科技工作者进一步努力。
在不同的温度条件下,不同品种的黄瓜幼苗在外部形态和内部生理代谢方面都会发生不同程度的变化。
本试验主要研究两个抗冷性不同的黄瓜品种的幼苗生理生化指标的变化,这些指标主要涉及叶绿素含量,根系活力,mda含量,脯氨酸含量,通过研究这些逆境指标的变化来分析其发生冷害的机理,探讨黄瓜在不同温度下的抗冷性机制,为提高黄瓜的抗冷性研究提供参考与依据。
1、材料与方法1.1 试验材料供试品种为“天津优2号”(山东省宁阳县鲁明种子有限公司提供)简称a;“夏丰一号”(天津市宏丰蔬菜研究所生产提供)简称b。
1.2实验方法1.2.1 种子处理对所选黄瓜种子消毒,55℃清水中浸泡10min,浸泡过程中不断搅拌,然后20℃清水中浸种8h,最后25℃温箱催芽12h,过程中冲洗,待萌发。
1.2.2 育苗及幼苗处理方法采取营养土育苗方式,将萌发种子点播于塑料盆中[4],每盆播两品种,分开点播,每品种点播12株,共6盆。
幼苗长出后进行常规管理,待幼苗长至一叶一心[6],真叶展平时进行不同温度处理。

弱光和低温对大棚蔬菜的影响冬春季利用设施栽培蔬菜,当外界温度低于蔬菜正常生长的适宜温度下限时就会发生冷害或冻害。
其植物的表现有下面几个方面:1.叶片受害:在子叶期受害表现为子叶边缘失绿,出现“镶白边”,温度恢复正常后真叶生长不受影响;定植后如遇短期低温或冷风侵袭,植株部分叶片边缘受影响会呈暗绿色,然后逐渐干枯。
2. 根系受害:表现为根系生长停止,不能发生新跟,部分老根开始发黄,逐渐死亡。
即使温度很快回升,植株会表现萎蔫货生长速度减慢。
受害严重的植株难以恢复生长。
3. 生长点受害:较为严重的冻害发生时,顶芽受冻,不发新叶。
在天气转暖后可考虑补苗。
4.花果受害:蔬菜在开花期遇低温天气的影响,授粉受精率降低,稍后会形成大量的落花落果或畸形果。
对茄子的影响茄子(Solanum melongean L.)起源于亚洲东南亚热带地区,属喜温性的低温敏感型蔬菜。
近年来,随着我国节能型日光温室的普及与推广,茄子已成为设施栽培的主要果菜之一,但是在反季节生产时,覆盖物和骨架结构遮光及冬春季节的低温、雨、雪、连阴天等不良气候条件,使设施内的植物经常在低温弱光逆境中生长,从而造成作物徒长、光合能力下降和抗病虫害能力的减弱,对于像茄子以果实为产品器官的作物,低温弱光还会影响到开花、坐果及果实的发育,最终导致产量和品质的下降。
因此,要实现节能日光温室茄子等蔬菜产业的可持续发展,除了进一步改善日光温室的性能、培育日光温室专用品种,加强病虫害防治外,还需要进一步了解影响茄子光合生理特性的外界环境条件,掌握其影响机理,通过应用栽培管理措施和改善环境条件,以最大限度地发挥日光温室的生产潜力。
温室越冬茄子正处于低温弱光期,这段时间根系吸收肥力能力下降,很容易受营养元素的影响而出现异常症状(如叶片黄化、僵果、裂果、花不开放、落花落果等)。
1、叶黄化缺铁植株顶部幼叶失绿严重,呈黄白色,叶脉保持绿色,由于铁元素在植株体内移动性小,所以下部叶片发生较少,往往在顶部叶片上发生较多。



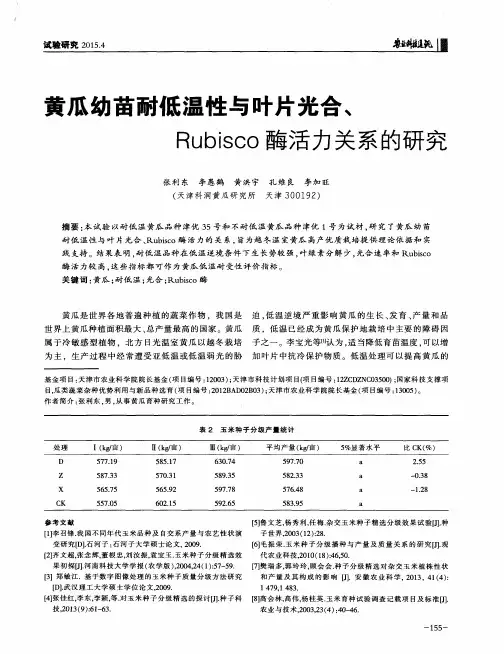

低温胁迫下黄瓜幼苗DNA甲基化与抗性的关系DNA甲基化是一种重要的表观遗传修饰方式,它通过给DNA分子附加一个甲基基团来改变基因的活性。
研究表明,DNA甲基化在植物对抗逆境胁迫的响应中扮演着至关重要的角色。
低温胁迫是黄瓜幼苗生长中的常见环境压力之一,因此,了解低温胁迫对黄瓜幼苗DNA甲基化水平的影响,对于揭示黄瓜幼苗抗逆性的调控机制具有重要的意义。
研究发现,低温胁迫会显著影响黄瓜幼苗的DNA甲基化水平。
低温胁迫引起了DNA甲基化水平的整体上升,并且促进了DNA甲基化剧烈变化区域(DMRs)的形成。
DMRs通常与基因的表达水平和功能相关,因此低温胁迫诱导的DNA甲基化变化可能对黄瓜幼苗的适应性起到重要的调控作用。
进一步的研究表明,低温胁迫诱导的DNA甲基化变化与黄瓜幼苗的抗性密切相关。
一方面,一些特定的DMRs与抗性相关基因的表达发生了变化,如一些抗冷胁迫相关基因的表达上调。
另一方面,DNA甲基化的变化也直接影响了一些关键抗性相关基因的表达调控。
这些发现表明低温胁迫通过调节黄瓜幼苗的DNA甲基化水平来调控抗性相关基因的表达,从而改善黄瓜幼苗对低温胁迫的抵抗能力。
进一步的探究发现,DNA甲基化修饰在低温胁迫触发的抗性进行中可能通过多种方式进行调控。
首先,DNA甲基化修饰可以影响DNA序列的稳定性和可读性,从而通过调节基因的转录和翻译过程来调控抗性相关基因的表达。
其次,DNA甲基化修饰可以影响组蛋白的修饰模式,从而调节染色质结构的开放程度和基因区域的可访问性。
最后,DNA甲基化修饰还可以影响非编码RNA的产生和功能,从而进一步调控基因表达的复杂网络。
综上所述,低温胁迫对黄瓜幼苗的DNA甲基化水平具有显著影响,并且这种甲基化变化与黄瓜幼苗的抗性密切相关。
低温胁迫诱导的DNA甲基化变化通过调节抗性相关基因的表达,改善了黄瓜幼苗对低温胁迫的抵抗能力。
进一步的研究有助于揭示DNA甲基化修饰在黄瓜幼苗对逆境胁迫的响应机制中的具体作用,为培育抗寒性强的黄瓜品种提供理论依据和遗传改良策略综合以上研究结果,我们可以得出结论:低温胁迫可以通过调节黄瓜幼苗的DNA甲基化水平来改善其对低温胁迫的抵抗能力。

冬季黄瓜栽培常温、低温和高温管理方法常温、低温、高温是黄瓜栽培种三种不同的管理方法,在不同管理条件下作物生长状况也不一样。
下面就来介绍一下冬季黄瓜栽培不同温度的不同管理方法。
常温管理是采用一般栽培技术要求的温度指标,白天25-28℃,最高32℃;夜间14-16℃。
低温管理白天23-25℃,最高28℃;夜间12-14℃。
高温管理白天30-35℃,最高42℃;夜间18-20℃。
一、不同管理条件下的作物生长状况1、常温管理时的生长状况常温管理时,植株生长健壮,结果稳定,结果时间长,一般以总产量来保证总产值。
在新菜区,保护地蔬菜生产量不大,季节性差价不太明显时,适宜于采用这种管理方法。
但其缺点是较难控制霜霉病的发生,应注意防治。
2、低温管理时的生长状况低温管理是利用简易温室栽培时,在低温寡照时期采用的一种适应性的管理方法。
在主要环境条件不能完全满足作物正常生长发育时,以保全植株为前提,获取一定产量的管理方法。
3、高温管理时的生长状况高温管理时,叶片明显变大,结果快而集中,但结果期缩短,植株过早衰败。
这种通过集中获取早期产量来获得高产值的方法,适宜于季节差价比较明显的地方。
高温管理也能够比较有效地控制霜霉病的发生蔓延,在条件和管理水平可以达到时,值得推广。
二、常温管理技术1、缓苗期注意保温黄瓜定植后要注意提高室内温度,促进早缓苗。
草帘应晚揭早盖,白天室内温度超过32℃时盖帘遮荫。
此期要求的适温为白天25-30℃,夜间15℃左右。
要浇足定植水,覆盖地膜;根据土壤墒情采用膜下暗灌或滴灌的方法浇好缓苗水。
2、始花期促根控秧缓苗后以促根控秧为中心,控制地上部生长,促进根系发育。
加大昼夜温差,进行变温管理,严格控制灌水量。
白天温度超过30℃时,从温室顶部通小风,随着温度的升高增大通风口;当温度降到25℃时,缩小通风口;降到20℃时,关闭通风口。
下午气温降到15-18℃时覆盖草帘,前半夜控制在15-18℃,后半夜控制在12-14℃,早晨揭开草帘前温度应降至10℃左右,不得低于8℃。

黄瓜低温危害及其补温措施(1)叶片受害叶片受害分低温受害和冻害。
叶片受到轻微冻害时,子叶期表现为叶缘失绿,有镶白边的现象,温度恢复后不会影响以后真叶的生长。
定植后受到冻害时,植株部分叶片的叶缘呈暗绿色,逐渐干枯。
这种情况在冬季或早春温室棚膜上的破洞不能及时修补,冷风由此侵入时,也容易看到这一现象。
温室中成长的植株,如果棚膜突然被大风吹毁,寒风突然侵入,会发现叶片上立即出现镀铝样的银白色,叶片也随之凋萎,而且无挽救的希望。
长期处于最低界限温度以下的黄瓜植株,叶片往往会表现出一些异常症状:①叶尖下垂,出现枫树叶。
夜温15℃以上时,叶片呈水平状展开。
在15℃以下时,叶尖下垂,周缘起皱纹。
低温下发育的叶子缺刻深,叶身长,像枫树叶状。
②虎斑叶。
低温下叶面呈现虎斑状,即主脉间叶肉褪绿变黄。
瓜条膨大受抑制。
这是由于光合作用制造的碳水化合物不能及时地向外部运转而在叶内沉积下来所造成的,严重时整个叶片会随之黄化。
如果温度回升且能维持一段时间,碳水化合物能够顺利充分地转换时,这时瓜条便可顺利膨大,叶片也能慢慢恢复转黑。
但此种机会常不易遇到。
③“水浸症”。
低温危害严重且持续时间长,温室温度大而较少通风时,叶被面会出现“水浸症”。
“水浸症”是由于夜间气温低,尤其在地温尚高时,细胞里的水分流到了细胞间隙中而引起的。
植株长势好时,水浸状可在太阳出来后消失。
但若植株衰弱或完全衰弱时,白天温度升高后水浸状也不消失,这样几经反复,细胞死亡,叶子枯死。
④龙头呈“开花型”。
生长发育和温度正常时,从侧面看龙头侧呈棉花蕾状,两片嫩叶围着顶芽。
但若夜温低、地温低(肥料不足或受病虫为害)时,龙头呈开花状,即两片应围着顶芽的嫩叶展开,顶芽伸出。
龙头呈开花型时,开花节位距顶端仅20~30cm,有时开花好象在顶端。
⑤出现缺硼或缺镁症。
夜温降到生物学零度以下时,由于植株素质变弱;或因为连年种植,过多施用化肥或有机肥少地力下降等,使根对硼的吸收力下降,引起缺硼症,其主要症状是生长点生长停止。
低温天气对农作物有何影响?
一、气温骤降引发农作物生长受阻
1. 气温骤降将导致农作物受到冻害,甚至死亡。
2. 低温环境下,农作物生长速度减缓,影响产量和品质。
二、低温会影响农作物的生理过程
1. 低温会导致植物细胞液体结冰,破坏细胞结构。
2. 长时间的低温环境可能造成植物叶片变脆、出现霜冻斑点。
三、低温天气影响农作物的生长周期
1. 低温延长了农作物的生长周期,导致生长发育不均匀。
2. 部分农作物在低温环境下会出现抽薹不良、花期错乱等现象。
四、低温天气对农作物的病虫害防治产生影响
1. 低温气候可能导致农作物病虫害的死亡率减少。
2. 低温环境下,部分病虫害的发生可能会受到影响,但某些病虫害会更容易滋生。
五、低温天气引发的土壤问题对农作物生长造成挑战
1. 低温天气会导致土壤冷害,影响植物根系正常生长。
2. 土壤中含水率降低,植物吸收水分受到限制,生长发育受阻。
霜降对农村蔬菜种植的影响霜降是中国二十四节气之一,标志着冬季气温进一步下降,阳光照射时间减少,夜间霜冻的发生频率增加。
对于农村蔬菜种植来说,霜降会带来一系列的影响,包括栽培技术调整、灾害风险管理、产量和质量的变化等。
本文将重点探讨霜降对农村蔬菜种植的影响。
一、温度下降影响生长周期和品质霜降后,气温进一步下降,这对蔬菜生长周期和品质都会带来一定的影响。
一方面,低温会延长作物生长周期,导致种植周期变长,延迟采收时间。
例如,霜降后种植的辣椒、茄子等热带作物往往需要更长时间才能成熟。
另一方面,低温还会影响蔬菜的品质。
一些蔬菜如黄瓜、茄子等对温度敏感,低温容易导致果实变软、品质下降。
因此,在霜降时期,农民需要采取相应的措施,如增加保温措施,调整栽培技术等,以保证蔬菜的正常生长和品质。
二、蔬菜产量受限霜降对蔬菜产量也会带来一定的限制。
低温和霜冻可能导致一些蔬菜植株出现受冻受灾的情况,使得植株受损甚至丧失部分或全部的产出能力。
特别是霜降后,晴天夜晚的低温极易导致蔬菜植株受冻,造成严重的损失。
为了减缓低温对产量的不利影响,农民可以采取一些应对措施。
例如,自制简易温室框架、增设保温覆盖物等,增加温室效应,提高环境温度,减少蔬菜植株被冻害的概率,从而降低产量损失。
三、病虫害防治加强霜降过后,温度的下降会使得一些病虫害的发生风险增加。
低温会导致一些常见的蔬菜病虫害的传播速度减缓,但也会增加其他病虫害的风险。
例如,霜降后长时间的湿冷天气有利于细菌的繁殖,容易引起一些病害。
因此,农村蔬菜种植者应加强病虫害的防治工作,如定期喷洒农药、加强病害监测等,以及保持合理的温湿度,减少病虫害发生的机会。
四、农耕活动调整霜降对农耕活动也会有一定的调整要求。
一方面,农民需要加强田间土壤管理,避免因低温导致的土壤冻结,破坏土壤结构和肥力。
此外,农民还需要做好蔬菜的蓄温保暖措施,如覆盖地膜、秸秆等,减少土壤温度变化,保证蔬菜正常生长。
另一方面,霜降后,农民也需要根据作物生长特点和气温变化,合理安排田间管理活动,如采摘、除草、追肥等,以提高蔬菜的产量和质量。
黄瓜生长发育需要什么样的温度?
黄瓜属喜温性蔬菜。
叶片生长的适宜温度白天为25至28摄氏度,夜间12至15摄氏度。
结瓜期适宜温度白天25至32摄氏度,夜间15摄氏度左右。
温度降到10至12摄氏度以下,植株生理活动失调,生长缓慢或停止生长。
温度低于5摄氏度以下难以适应,持续2至3天,植株会受寒害。
遇0摄氏度以下的低温时,随即冻死。
但黄瓜不同生长时期的耐寒能力差异较大,苗期的耐寒能力最强,结瓜期最弱。
但苗期经过低温锻炼的植株,结果期的抗寒能力有所增强,可忍耐短期3摄氏度以下、0摄氏度以上的低温。
因此,大棚早春栽培黄瓜时,苗期应进行抗寒炼苗。
黄瓜不耐高温,一般情况下,温度高于32摄氏度以上时,植株的光合作用开始下降。
超过35摄氏度时,生理活动会发生障碍。
40摄氏度以上,生长几乎停止,并迅速衰败。
持续45摄氏度3小时的高温,茎叶虽不发生直接伤害,但以后叶色变淡,雄花落蕾或不能开花。
50摄氏度的高温持续1小时,呼吸作用几乎停止。
温度再高,则发生日烧。
黄瓜对地温的要求更为严格,对地温的变化非常敏感。
黄瓜根伸长的最低温度为8摄氏度,最高温度为38摄氏度,生长发育最适宜的地温为25摄氏度左右。
若地温降至12度以下,根系的生理活动受阻,会引起下部叶片黄化。
在栽培中一般最低温度应维持在15摄氏度以上,最高温度不超过30摄氏度。
地温过高,易加速根系的老化。
- 1 -。
低温逆境对黄瓜生长发育的影响高福钊霍建勇(辽宁农业发展有限公司沈阳110034)黄瓜是人们喜爱的主要的蔬菜作物之一,黄瓜喜温、喜光,不适的温度影响黄瓜的正常生长发育。
在逆境条件不仅影响了黄瓜的水分和矿质代谢,还影响着光合作用和呼吸作用,造成黄瓜生长发育不良,产量、质量下降。
近年来随着保护地生产的迅猛发展,低温逆境对黄瓜生长发育的影响显得越来越突出。
1 不适低温对黄瓜生长发育及生理生化指标的影响黄瓜最适的萌发温度为30℃,黄瓜生长发育最适温度为白天25℃左右,夜间13℃左右。
黄瓜低温温度补偿点为3.3℃,低温下老叶的温度补偿点明显高于新叶(张振贤等,2003)。
昼夜温度低于24/10℃,根系活力受到抑制,温度低于16/6℃时,根系活力下降30%-60%,低温对黄瓜幼苗的生物学产量的影响非常明显,对根系的影响大于对茎叶的影响,黄瓜幼苗根系忍耐低温的下限比茎叶高(王克安等,2000)。
低温对发芽率的影响不大,仅延缓了发芽的速度。
马德华等(1999)认为耐低温能力较强的自交系耐高温能力亦较强,而耐低温能力较差的自交系耐高温能力也较差,部分黄瓜品种的幼苗对不同温度逆境抗性之间存在一定的相关性。
宋勇等(2003)试验表明黄瓜经低温(最适低温为10℃)或高温(最适高温为30℃)预处理4小时,在随后的高温(35℃)或低温(15℃)下萌发,能明显地提高其活力,说明黄瓜种子对高温和低温胁迫有交叉的适应能力。
2 不适的低温对黄瓜的细胞生理生化特性的影响低温不仅对黄瓜的形态特性造成影响,更重要的是对黄瓜的细胞生理生化特性产生影响。
马德华等研究认为黄瓜幼苗在低温下细胞的膜脂过氧化程度明显加剧,膜脂过氧化产物丙二醛(MDA)含量显著增加,过氧化物酶(POD)活性上升,过氧化氢酶(CAT)活性下降,POD活性的上升和CAT活性的下降与黄瓜幼苗耐性呈正相关。
李淑艳等(2002)的试验表明在冷害温度下处理结束时植物体内的超氧化物歧化酶(SOD)、CAT活性、POD含量均高于对照,在亚适温条件下SOD活性、脯氨酸含量高于对照,CAT活性则低于对照。
低温逆境诱导了抗氧化剂酶系的活力,植物体内抗氧化剂酶系包括SOD、POD、CAT,植物处于逆境条件下,细胞内活性氧(O2-、H2O2、OH-)的产生和清除的平衡遭到破坏,产生过多的活性氧,导致膜脂过氧化,从而造成对细胞膜的伤害。
植物体内过多的活性氧的清除,主要依赖于抗氧化剂酶系统(韩建明等,1999)。
低温逆境还使脂类物质发生变化。
杨玲(2001)用黄瓜子叶在15℃处理下,虽无可见的冷伤害症状,但类脂成分已发生显著变化,双半乳糖甘油二脂(DGDG)含量明显升高,而磷酸脂酰胆碱(PC)、磷脂酰乙醇胺(PE)大幅度下降,主要类脂半乳糖甘油二脂(MGDG)、DGDG及PC和不饱和脂肪酸尤其是亚麻酸增多。
说明黄瓜子叶在15 ℃下处理7天已经对低温产生了冷适应性。
3 低温对黄瓜光合作用的影响光合作用最适温度为28-32℃,低温影响着黄瓜的正常的光合作用。
低温条件下光合速率、呼吸速率随温度的升高而迅速增加,温度逆境胁迫使叶绿素含量明显降低而且以叶绿素a下降为主,温度逆境胁迫后气孔导度(Gs)下降,表观光合量子效率(AQE)和光合效率(CE)降低,说明叶片的光合机制遭到了明显地破坏(马德华等1999)。
同等条件下低温锻炼的幼苗比未经锻炼的幼苗净光合速率降低较少,低温锻炼后和低温处理使幼苗气孔导度和叶肉细胞间CO2浓度降低,气孔限制值上升。
未经锻炼的幼苗在净光合速率降低的同时叶肉细胞间CO2浓度增加,气孔限制值下降。
说明非气孔因素是光饱和光合效率(Pn)降低的主要原因(庞金安和马德华,2000)。
低温下光照会加剧光合功能伤害。
陈启林等(2000,2001)试验表明在5℃低温条件下光照处理,比黑暗处理对黄瓜叶片光饱和光合速率、表观光量子速率、CO2饱和光合速率和羧化速率明显下降,低温光照与黑暗对光合作用的差异不是因为气孔关闭,而是由于叶肉细胞光合活性的降低。
PSⅡ光合中心受到较严重的光抑制,加NH4Cl和加ADP+Pi的Hill反应活性与基础Hill反应活性相差不大,从叶片中提取的叶绿素9-AA荧光猝灭能力消失,类囊体膜解耦联,类囊体跨膜质子梯度显著降低,ATP合成受限。
光照明显加剧了低温对叶片光合功能的伤害。
低温下光照加剧光合功能伤害的重要机理可能是由于Rubisco活化受到抑制。
低温光照处理不仅使Rubisco酶活性降低,引起RuBP羧化限制加重,还较为严重地降低RuBP再生速率,加剧对光合作用的伤害。
低温对不同生态类型黄瓜的光合特性的影响不同。
李长缨等(1997)认为冬春型黄瓜(如长春密刺、北京刺瓜)光补偿点受温度影响大。
在亚适温弱光下,较夏秋型品种有较多的干物质积累。
冬春型品种,在弱光下通过叶绿素含量的增加来提高捕光能力,以弥补光合效率的降低。
王永健认为在15℃偏低温条件下,耐弱光的保护地品种(如欧洲8号、北京刺瓜、日本5号、长春密刺)的光补偿点明显低于不耐低温弱光的品种。
王毅等(1995)采用不同耐寒性的黄瓜品种:农大14(高耐)、长春密刺(中耐)和津研2号(低耐),在15℃/5℃(日/夜)下处理5天后,虽除黄瓜幼叶绿色变淡外,无其它异常表现,在超微结构水平,叶绿体保持正常结构,但淀粉颗粒大为减少或完全消失,在连续5℃/5℃低温处理后,幼苗叶片明显黄化、萎蔫,超微结构水平相当部分叶绿体表现结构异常,双层被膜完整性丧失,基质和基粒片层变得松散,一些片层内腔膨大,表明叶片结构受到严重损伤,细胞失去正常的功能。
4 黄瓜的抗低温鉴定指标及抗寒诱导于拴仓等(2000)认为低温条件下的胚根相对伸长率可以反映品种间的耐低温性差异,浸种后36-48小时为耐低温性鉴定的适期,浸种36小时的种芽经(2.5±0.5)℃处理24小时恢复期的胚根生长率可以区别品种间的低温耐受性。
用种子活力综合指标、超弱发光指标及胚根伸长指标构建的黄瓜芽期低温鉴定指标体系可以全面、准确地反映黄瓜的低温耐受性。
王克安(2000)认为,耐寒性不同的黄瓜品种根系对低温胁迫的适应性不同,一定的低温条件下,幼苗根系活力可作为判断幼苗受害程度和黄瓜品种间抗寒能力的参考指标。
马德华等认为黄瓜幼苗在低温下膜脂过氧化产物丙二醛(MDA)含量显著增加,是较好地鉴定黄瓜幼苗低温能力的指标之一。
低温处理可以提高黄瓜的抗逆性。
李宝光等认为,适当降低育苗温度,可以增加叶片中抗冷保护物质。
低温处理,SOD、POD的活性升高,电解渗透率和MDA 含量降低。
低温锻炼逐步进行可进一步提高黄瓜的抗冷性(先昼25-30℃,夜13-15℃育苗,再昼15-25℃夜5-6℃锻炼4-6天)。
马德华等(1998b)用强低温处理后,幼苗光合速率急剧下降,即使恢复到常温,光合能力也不能恢复到原来的水平,但耐低温品种受到的影响较小。
表明选育并选用耐低温能力强的品种是避免短期低温危害的有效途径。
高丽红等(2000)试验表明在亚适温条件下欧亚杂交种植株营养生长和生殖生长表现比欧洲品种种亚洲品种均好,地上部分健壮,根系活性高,亚洲品种对低温的适应性明显好于欧洲品种。
孙艳等(2000)试验表明适宜浓度的水杨酸SA能显著地提高黄瓜的幼苗的壮苗指数,促进壮苗的形成,SA的最佳浓度为250mg/L。
当低温胁迫时SA可显著地提高黄瓜幼苗叶片细胞膜的稳定性,抑制叶片中MDA的累积。
佘小平等(1997)试验表明:PP333能有效地降低黄瓜幼苗在低温下的相对电导率,维持较高的SOD 活性和VC、还原型谷胱甘肽(GSH)含量,因而提高了幼苗的抗冷能力。
5黄瓜在温光逆境上的遗传研究进展顾兴芳等(2002)研究表明:控制低温下黄瓜相对发芽势、相对发芽指数、相对胚根长度符合加显模型,以显性效应为主,各性状相对遗传力分别为98.1%、96.9%、98.6%,狭义遗传力为24.0%、28.6%、37.9%,控制各性状的显性基因可能为寡基因或寡基因组。
而低温下相对发芽率的遗传不符合加显模式,控制该性状的显性基因可能有2组,并存在上位作用。
与耐低温有关的黄瓜的单性结实性状一般配合力起主要作用,遗传以加性效应为主,其加性方差占总方差的96.78%(韩建明等1999)。
参考文献1.陈启林,山仑,程智慧等.低温弱光对黄瓜类囊体膜耦联度状态的影响.西北农业大学学报,2000,28(6):6-11 2.陈启林,山仑,程智慧.低温下光照对黄瓜叶片光合特性的影响.中国农业科学,2001,34(6):632-636 3.韩建明,张春奇,范丙友.黄瓜基础理论研究进展.长江蔬菜,1999,(10):1-4 4. 顾兴芳,封林林,张春震,方秀娟,张天明.黄瓜低温发芽能力遗传分析.中国蔬菜,2002(3):5-7 5. 李宝光,卢育华,徐坤等.育苗温度对黄瓜幼苗抗冷性的影响.山东农业大学学报(自然科学版).6. 李长缨,朱其杰.光强对黄瓜光合特性及亚适温下生长的影响.园艺学报,1997,24(1):97-99 7.李淑艳,于锡宏.冷胁迫对黄瓜幼苗抗寒相关生理指标的影响.北方园艺,2002(6):52-53 8.马德华,庞金安,霍振荣,李淑菊,王全.黄瓜幼苗光合与呼吸特性研究.园艺学报,1997,24(4):397-398 9. 马德华,卢育华,庞金安.低温对黄瓜幼苗膜脂达氧化的影响.园艺学报1998,25(1):61-64 10.马德华,庞金安,霍振荣等.黄瓜对不同温度逆境的抗性的研究.中国农业科学,1999,32(5):28-35 11.马德华,庞金安,李淑菊等.短期强低温处理对黄瓜幼苗光合作用的影响.河北农业大学学报,1998b,21(2):39-42 12.宋勇,肖深根,何长征.黄瓜种子萌发的逆温耐性诱导.湖南农业大学学报(自然科学版),2003,29(20):127-128 13. 孙艳,崔鸿文,胡荣.水杨酸对黄瓜幼苗壮苗的形成及抗低温胁迫能力的生理效应.西北植物学报,2000,20(4):616-620 14.庞金安,马德华,霍振荣.低温锻炼对黄瓜幼苗光合作用的影响.河南农业大学学报,2000,34(1):40-42 15.佘小平,李忠歧,杨建雄等.PP333对黄瓜幼苗素质及抗冷能力的影响.西北植物学报,1997,17(2):205-209 16.王毅,方秀娟,徐欣,刘立新.黄瓜幼苗低温锻炼对叶片细胞叶绿体桔构的影响.园艺学报,1995,22(3):299-300 17. 王克安,王冰,顾三军.低温对黄瓜生理生化、代谢功能及形态的影响.山东农业科学,1998(4):52-55 18.王克安,何启伟,陈运起等.低温对黄瓜根系活力及生物学产量影响的研究.山东农业科学,2000(4):17-19 19 .杨玲.不同低温处理对黄瓜子叶极性脂组成的影响.园艺学报,2001,28(1):36-40 20. 高丽红,张福墁,陈青君.不同类型黄瓜生长发育与环境因子关系.中国农业大学学报,2000,5(3):58-62 21. 于拴仓,崔鸿文,孟焕文.黄瓜发芽期耐低温性鉴定方法与指标的研究.西北农业大学学报,2000,28(3):1-6 22. 张振贤,艾希珍,赵世杰,杨红艳.黄瓜叶片光合作用的温度补偿点与光合启动时间.园艺学报,2003,30(2):157-162 (该文发表于《吉林蔬菜》2004.第一期)。