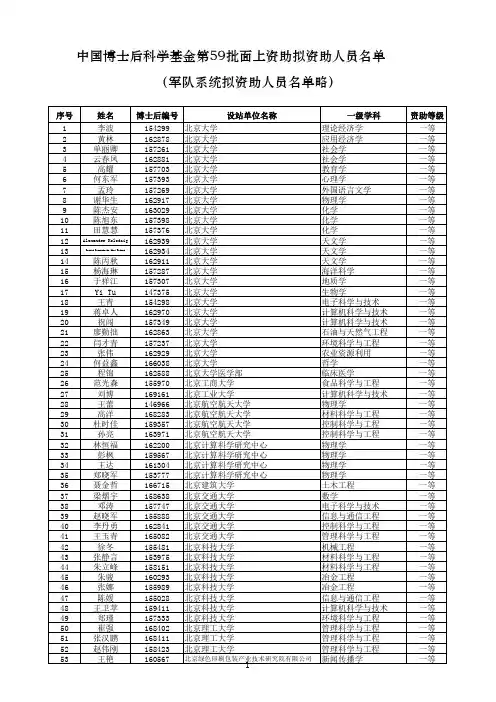
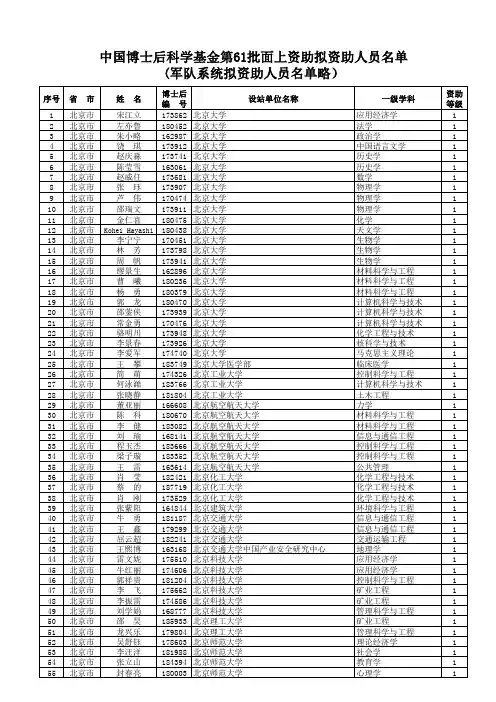
中国博士后科学基金第56批面上资助获资助人员名单
- 格式:pdf
- 大小:1.84 MB
- 文档页数:46


中国博士后科学基金第四十九批面上资助一等资助(五万元)生物类名单:(注:军队系统获一等资助的博士后人员10人,因涉密未列出。
)序号姓名单位学科3 王旭初北京大学生物学7 周妍妍北京大学医疗卫生医疗集团鸡西矿业总医院中医学56 夏永刚黑龙江中医药大学中药学57 文李湖南农业大学作物学65 舒迎花华南农业大学生物学66 黄超华中科技大学药学70 杨艳萍吉林大学临床医学81 朱泽章南京大学医学院附属鼓楼医院临床医学104 巩晶山东大学生物学105 王雪鹏山东农业大学兽医学109 王靖方上海生物信息技术研究中心生物学110 汪澈沈阳农业大学生物学112 夏霖四川大学临床医学114 秦秀德天津中医药大学中医学119 郑红云武汉大学临床医学131 杨向兵西北农林科技大学生物学135 黄正行浙江大学生物医学工程138 方琦浙江大学植物保护145 程民中国科学技术大学基础医学148 王鹏中国农业大学畜牧学149 马磊中国农业科学院畜牧学150 李欣中国农业科学院植物保护156 张金艳中国中医科学院中西医结合157 郝岗平中国中医科学院中药学158 高国庆中国中医科学院中医学160 黎大勇中国科学院动物所生物学169 沈婵娟中国科学院上海生命科学研究院生物学170 李青中国科学院上海生命科学研究院生物学172 陈培华中国科学院生物物理研究所生物学173 赵学金中国科学院微生物研究所兽医学174 冷艳中国科学院武汉病毒研究所生物学二等资助(3万元)209 李娟北京大学生物学210 刘国华北京大学生物学211 刘晓颖北京大学生物学212 徐菁北京大学生物学213 于凌飞北京大学生物学239 白光一北京大学医学部临床医学240 胡晓青北京大学医学部临床医学241 王雪艳北京大学医学部临床医学242 昝珂北京大学医学部药学260 王梦航北京航空航天大学生物医学工程269 刘宏北京化工大学食品科学与工程301 张丹丹北京师范大学生物医学工程309 王冬北京市农林科学院食品科学与工程315 刘铁北京协和医学院临床医学316 肖静北京协和医学院药学317 罗焜北京协和医学院中药学360 侯一民电子科技大学生物医学工程385 彭进松东北林业大学化学393 李金龙东北农业大学兽医学394 孙东波东北农业大学兽医学412 卢绪章东南大学临床医学413 邓大伟东南大学生物医学工程414 武灵芝东南大学生物医学工程420 郑武福建农林大学生物学429 陈俊复旦大学化学430 凌星复旦大学化学431 夏奡复旦大学化学435 杨冬琴复旦大学临床医学439 郭辉复旦大学生物学440 刘晓磊复旦大学生物学441 莫文娟复旦大学生物学442 姚小贞复旦大学生物学443 张雯婷复旦大学生物学444 朱永青复旦大学生物学461 李新明甘肃省农业科学院食品科学与工程465 李茹广西大学生物学466 申佩弘广西大学生物学468 夏星广州中医药大学中药学471 黄伟国家海洋局第二海洋研究所海洋科学472 曾淦宁国家海洋局第二海洋研究所海洋科学473 胡利民国家海洋局第一海洋研究所海洋科学475 周波国家人类基因组南方研究中心生物学524 王长林哈尔滨工业大学生物学525 侯爱菊哈尔滨工业大学食品科学与工程556 崔璨哈尔滨医科大学临床医学557 甘润韬哈尔滨医科大学临床医学558 蒋晶哈尔滨医科大学临床医学559 康欣梅哈尔滨医科大学临床医学560 孔瑞哈尔滨医科大学临床医学561 李杰哈尔滨医科大学临床医学562 马辉哈尔滨医科大学临床医学563 王静萱哈尔滨医科大学临床医学564 王威哈尔滨医科大学临床医学565 原劲杨哈尔滨医科大学临床医学566 宋武琦哈尔滨医科大学生物学567 于丹红哈尔滨医科大学生物学571 武永刚河北大学化学573 王辉河北师范大学生物学574 王瑞菊河北师范大学生物学586 田智勇河南大学化学587 朱金花河南大学化学600 赵景祥黑龙江大学化学606 吕典秋黑龙江省农业科学院生物学607 王知斌黑龙江中医药大学中药学609 但红湖北中医药大学中药学610 郑淑琴湖南大学化学614 黄国华湖南农业大学生物学615 易有金湖南农业大学食品科学与工程656 王尉华南理工大学临床医学659 王晓华华南理工大学食品科学与工程664 傅嘉欣华南农业大学生物学669 Talgat Inerbaev 华南师范大学化学670 高爱梅华南师范大学化学673 RadhakrishnanK.V 华南师范大学生物学674 胡博华南师范大学生物学694 乔现锋华中科技大学化学695 王建颖华中科技大学化学702 刘静华中科技大学临床医学704 徐承启华中科技大学生物学714 王阶平华中农业大学生物学715 周雁华中农业大学生物学716 方瑞华中农业大学兽医学717 戢凤琴华中农业大学兽医学718 王春梅华中农业大学兽医学720 毛永江华中农业大学畜牧学721 牛丽莉华中农业大学畜牧学741 黄宜兵吉林大学化学742 李春光吉林大学化学743 刘鸿吉林大学化学744 漆奇吉林大学化学745 辛毅吉林大学化学746 张国华吉林大学化学747 郑传涛吉林大学化学759 王国庆吉林大学临床医学762 王会岩吉林大学生物学777 邓穗平暨南大学生物学778 田海妍暨南大学药学779 于洋暨南大学药学793 陈慧卿江苏大学生物学794 任乾江苏大学生物学795 徐庆刚江苏大学生物学796 麦维军江苏大学水产学797 朱克明江苏大学作物学803 刘丹江西省人民医院药学820 王玉珍内蒙古农业大学食品科学与工程822 吴信南昌大学食品科学与工程823 何昕华南方医科大学生物学835 陈永飞南京大学化学836 方瑞琴南京大学化学837 钱立武南京大学化学838 任雪峰南京大学化学839 陶传洲南京大学化学840 张东恩南京大学化学841 张载超南京大学化学847 何伟奇南京大学生物学848 薛佳宇南京大学生物学849 夏友谊南京大学生物医学工程853 肖竹平南京大学药学887 黄志锋南京理工大学药学895 习丙文南京农业大学水产学896 梁明祥南京农业大学作物学897 牟会荣南京农业大学作物学905 丁海霞南京医科大学临床医学906 赵鹏南京医科大学临床医学907 郑东辉南京医科大学临床医学914 DávidRédei南开大学生物学919 徐广贤宁夏医学院附属医院临床医学922 姜彦青岛大学临床医学946 李立宏清华大学化学947 张佳玮清华大学化学948 吴筱清华大学化学工程与技术949 俞鸣铗清华大学化学工程与技术981 单晓昳清华大学生物学982 梁斌清华大学生物学983 王沙清华大学生物学984 严晓华清华大学生物学985 张丹清华大学生物医学工程1002 李西文清华大学药学1029 季伟山东大学光学工程1030 张俊豪山东大学化学1031 邢伟山东大学化学工程与技术1047 赵秀鹤山东大学临床医学1049 李瑶瑶山东大学生物学1050 秦启龙山东大学生物学1051 苏擘山东大学生物学1052 魏天迪山东大学生物学1053 朱春原山东大学生物学1054 刘澄玉山东大学生物医学工程1063 孙斌山东大学药学1064 王延风山东大学药学1078 张锦丽山东农业大学食品科学与工程1087 王健陕西师范大学生物学1107 孔浩上海交通大学化学1108 王胜上海交通大学化学1122 李金文上海交通大学作物学1123 李佳上海交通大学临床医学1124 石国军上海交通大学临床医学1125 张敏光上海交通大学临床医学1126 袁圆阳上海交通大学生物学1127 郑秀娟上海交通大学生物医学工程1128 王伟卫上海交通大学药学1137 王克鹏深圳北京大学香港科技大学医学中心临床医学1138 陈浩深圳北京大学香港科技大学医学中心生物学1139 刁亚锐深圳北京大学香港科技大学医学中心生物学1157 王永杰首都师范大学生物学1159 范一帆首都医科大学临床医学1160 刘亚静首都医科大学临床医学1161 许杰首都医科大学临床医学1165 尚书勇四川大学化学工程与技术1169 陈芹四川大学临床医学1170 韩琳四川大学临床医学1171 陆慧敏四川大学临床医学1172 石华山四川大学临床医学1173 田丽四川大学临床医学1174 薛建新四川大学临床医学1175 邹俊四川大学临床医学1177 孙华钦四川大学生物学1178 常菁四川大学生物医学工程1179 刘博四川大学生物医学工程1182 刘捷四川大学药学1183 欧阳亮四川大学药学1188 孙万平苏州大学临床医学1192 李峰苏州纳米技术与纳米仿生研究所化学1195 韩丽娜太原理工大学化学工程与技术1211 刘清岱天津科技大学食品科学与工程1214 李树颖天津医科大学临床医学1215 刘亮天津医科大学临床医学1216 王晓芳天津医科大学临床医学1217 李西川天津医科大学生物学1218 滕杰天津医科大学中药学1246 陈贝贝武汉大学化学1247 崔然武汉大学化学1248 刘鹏武汉大学化学1249 肖丽武汉大学化学1262 黄伟武汉大学水产学1294 陈欣西安交通大学临床医学1295 王伟西安交通大学临床医学1304 任莹辉西北大学化学1305 尹兵西北大学化学1322 于修烛西北农林科技大学食品科学与工程1338 骆轩厦门大学海洋科学1339 王荣方厦门大学化学1340 颜伟忠厦门大学化学1351 韩炜新疆大学生物学1353 赵玲新疆农业大学生物学1359 李娟扬州大学化学1377 马文明浙江大学海洋科学1378 谢湖均浙江大学化学1379 祝莹浙江大学化学1395 王家传浙江大学临床医学1397 董海燕浙江大学生物学1398 张佳蓉浙江大学生物学1399 蔡华浙江大学生物医学工程1400 李加友浙江大学食品科学与工程1410 伍义行浙江大学药学1411 杨晓春浙江大学药学1423 刘宏波浙江大学作物学1424 曾凡荣浙江大学作物学1428 古华茂浙江工商大学食品科学与工程1430 高栋浙江海正集团有限公司化学工程与技术1431 夏道宗浙江中医药大学中药学1462 伊珍珍中国海洋大学生物学1579 董斌中国石油大学(华东)化学1581 关腾中国药科大学中药学1582 欧阳嶷中国医科大学临床医学1583 张光伟中国医科大学生物医学工程1589 马卫国中国中医科学院中西医结合1590 牛旭艳中国中医科学院中西医结合1591 任丽中国中医科学院中西医结合1592 孙明谦中国中医科学院中西医结合1593 谢鹏扬中国中医科学院中西医结合1594 许少华中国中医科学院中西医结合1595 郑光中国中医科学院中西医结合1596 陈颖中国中医科学院中药学1597 林青中国中医科学院中药学1598 刘越中国中医科学院中药学1599 郜洁中国中医科学院中医学1600 黄玉燕中国中医科学院中医学1601 李卫东中国中医科学院中医学1602 刘力力中国中医科学院中医学1603 桑希生中国中医科学院中医学1606 杨玮婷中国科学院长春应用化学研究所化学1607 杨艳中国科学院长春应用化学研究所化学1608 伊斐艳中国科学院长春应用化学研究所化学1609 赵先恩中国科学院长春应用化学研究所化学1611 龚波中国科学院成都生物研究所临床医学1612 黄璐琳中国科学院成都生物研究所生物学1613 罗艳中国科学院成都生物研究所生物学1614 张林中国科学院成都生物研究所生物学1616 房大维中国科学院大连化学物理研究所化学1617 申万岭中国科学院大连化学物理研究所化学1618 王周君中国科学院大连化学物理研究所化学1639 白雪中国科学院动物研究所生物学1640 王震波中国科学院动物研究所生物学1641 魏延昌中国科学院动物研究所生物学1642 张同作中国科学院动物研究所生物学1661 王鹏中国科学院海洋研究所海洋科学1662 周克中国科学院海洋研究所海洋科学1667 刘伟庆中国科学院合肥物质科学研究院化学1672 李靖中国科学院华南植物园生物学1673 苗青中国科学院华南植物园生物学1675 肖武名中国科学院华南植物园作物学1676 李晓娜中国科学院化学研究所化学1677 刘洪亮中国科学院化学研究所化学1678 刘巧玲中国科学院化学研究所化学1679 夏铁中国科学院化学研究所化学1680 杨惠中国科学院化学研究所化学1681 赵传壮中国科学院化学研究所化学1686 刘荻萩中国科学院昆明动物研究所生物学1687 贾祥凤中国科学院兰州化学物理研究所化学1689 程皓中国科学院南海海洋研究所海洋科学1696 欧阳新华中国科学院宁波材料技术与工程研究所化学1702 黄宗安中国科学院上海生命科学研究院生物学1703 吕亮东中国科学院上海生命科学研究院生物学1704 童一民中国科学院上海生命科学研究院生物学1705 徐锋中国科学院上海生命科学研究院生物学1706 曾涛中国科学院上海生命科学研究院生物学1707 郑广勇中国科学院上海生命科学研究院生物学1710 梅崇余中国科学院上海有机化学研究所化学1711 孙鹏中国科学院上海有机化学研究所药学1716 吝涛中国科学院生态环境研究中心生物学1717 孔瑞瑞中国科学院生物物理研究所生物学1718 姚明慧中国科学院生物物理研究所生物学1719 叶步青中国科学院生物物理研究所生物学1720 张家龙中国科学院生物物理研究所生物学1721 徐小兰中国科学院生物物理研究所药学1723 陈彩霞中国科学院微生物研究所药学1724 蔡昆中国科学院武汉病毒研究所生物学1729 王宁中国科学院心理研究所生物学1733 王雷中国科学院新疆生态与地理研究所生物学1741 符书兰中国科学院遗传与发育生物学研究所生物学1742 王慧玲中国科学院植物研究所生物学1750 陈述中南大学化学1751 梁文杰中南大学化学工程与技术1758 杨云波中南大学临床医学1773 陈嘉媚中山大学化学1774 李国璧中山大学化学1784 冯亚玲中山大学临床医学1785 李丹丹中山大学临床医学1786 李乃洋中山大学临床医学1787 刘颜颜中山大学临床医学1788 王红亮中山大学临床医学1789 陈禹中山大学生物学1790 赖芬菊中山大学生物学1791 杨灿朝中山大学生物学1792 苑占江中山大学生物学1796 何蓉蓉中山大学药学1797 李嘉丽中山大学药学1798 王小广中山大学药学。



第53卷 第11期 2023年11月中国海洋大学学报P E R I O D I C A L O F O C E A N U N I V E R S I T Y O F C H I N A53(11):087~098N o v .,2023黄河口及莱州湾海域磷的时空分布及浮游生物对低磷胁迫的响应❋段晓萌1,2,秦华伟3,马浩阳1,许泽昊1,2,吕浩然1,2,梁生康1,2❋❋(1.中国海洋大学化学化工学院,山东青岛266100;2.中国海洋大学海洋化学理论与工程技术教育部重点实验室,山东青岛266100;3.山东省海洋资源与环境研究院,山东烟台250299)摘 要: 本文利用2019 2021年在黄河口及莱州湾海域进行的4次陆海同步调查结果,分析了环莱州湾主要入海河口和排污口陆源输入磷的季节变化㊁黄河口及莱州湾海域内不同形态磷及不同碱性磷酸酶活性(A P A )的时空分布特征,探讨了海域内磷受限状况及浮游植物和浮游细菌对低磷胁迫的响应㊂结果表明,磷的陆源输入中黄河贡献最大,小清河次之;总磷(T P )入海通量呈现出夏季>春季>秋季>冬季的季节变化,春㊁夏㊁秋季磷输入以颗粒态磷(P P )为主,冬季以溶解态磷(D P )为主;受农业施肥及河道内浮游生物活动的影响,春季陆源D P 以溶解有机磷(D O P )为主,其他季节以活性磷酸盐(P O 4-P )为主㊂研究海域内T P 浓度及构成不仅受陆源输入的影响,还受浮游生物消亡㊁海上养殖活动和沉积物释放等作用共同调控,T P 浓度呈现出春季>夏季ʈ秋季>冬季的季节变化㊁近岸高远岸低的分布特征,高值区主要位于黄河和小清河河口区域;T P 构成上,春季以D P 为主,夏㊁秋季D P 与P P 相当,冬季以P P 为主;春㊁秋季D P 以D O P 为主,而夏㊁冬季D P 以P O 4-P 为主㊂海域内浮游植物普遍受到P O 4-P 的绝对限制和相对限制,夏季强降雨导致大量磷排放入海,研究海域磷限制情况得到缓解㊂海域内A P A 处于较高水平,秋㊁冬季A P A t o t a l 分别主要由A P A p h y ㊁A P A f r e e 贡献;当水体中D I N ʒP O 4-P 极高㊁P O 4-P 严重缺乏时,A P A p h y ㊁A P A b a c 较高,浮游植物及细菌主要依赖细胞周质及细胞膜上的A P 来水解D O P 以维持新陈代谢㊂P O 4-P 通过诱导-抑制机制对A P 进行调控,A P A 随P O 4-P 的增加而降低,当P O 4-P 低于0.05μm o l ㊃L -1时,浮游植物迅速分泌出大量的碱性磷酸酶来应对低磷胁迫;当高于阈值时,A P A 维持在较低水平㊂关键词: 黄河口;莱州湾;磷;时空分布;碱性磷酸酶活性;低磷胁迫中图法分类号: P 734 文献标志码: A 文章编号: 1672-5174(2023)11-087-12D O I : 10.16441/j.c n k i .h d x b .20220268引用格式: 段晓萌,秦华伟,马浩阳,等.黄河口及莱州湾海域磷的时空分布及浮游生物对低磷胁迫的响应[J ].中国海洋大学学报(自然科学版),2023,53(11):87-98.D u a n X i a o m e n g ,Q i n H u a w e i ,M a H a o y a n g ,e t a l .T e m p o r o s p a t i a l d i s t r i b u t i o n o f p h o s p h o r u s a n d r e s p o n s e o f p h y t o pl a n k -t o n t o l o w p h o s p h o r u s s t r e s s i n t h e Y e l l o w R i v e r e s t u a r y a n d L a i z h o u B a y [J ].P e r i o d i c a l o f O c e a n U n i v e r s i t y of C h i n a ,2023,53(11):87-98.❋ 基金项目:国家重点研究发展计划项目(2018Y F C 1407601);中央高校基本科研业务费专项资金(201962011,202042008)资助S u p p o r t e d b y t h e N a t i o n a l K e y R e s e a r c h a n d D e v e l o p m e n t P r o je c t of C h i n a (2018Y F C 1407602);t h e F u n d a m e n t a l R e s e a r c h F u n d s f o r t h e C e n t r a l U n i v e r s i t i e s (201962011,202042008)收稿日期:2022-05-10;修订日期:2022-06-06作者简介:段晓萌(1996 ),女,硕士生㊂E -m a i l :d x m 8338@s t u .o u c .e d u .c n❋❋ 通信作者:E -m a i l :l i a n gs k @o u c .e d u .c n 磷作为重要的生源要素,在海洋浮游植物和浮游细菌新陈代谢的过程中具有关键作用[1-3]㊂根据其存在形态,海水中的磷通常被分为溶解态磷(D i s s o l v e d i n o r ga n i c p h o s p h o r u s ,D P )和颗粒态磷(P a r t i c u l a t e p h o s ph o r u s ,P P )㊂D P 又以溶解无机磷(D i s s o l v e d i n o r g a n i c p h o s p h o r -u s ,D I P )和溶解有机磷(D i s s o l v e d o r g a n i c p h o s ph o r u s ,D O P )两种形式存在㊂D I P 的主要存在形式为正磷酸盐(P O 4-P ),P O 4-P 可以被大多数微生物直接吸收[4]㊂通常认为,当海水中P O 4-P <0.10μm o l ㊃L -1时,浮游生物无法从环境中获取足够的P O 4-P 来维持生长,即受到P O 4-P 的绝对限制;当溶解无机氮(D i s s o l v e d i n o r g a n i c n i t r o g e n ,D I N )与P O 4-P 比值超过16ʒ1时,认为浮游生物生长会受到P O 4-P 的相对限制㊂目前,许多河口区及近海海域的浮游生物生长受到磷的限制[5-11]㊂研究发现大部分D O P 需要通过相关酶类的转化为D I P 后才能被浮游生物利用[6,12],其中,含量占D O P 总量80%~85%的磷酸酯[13-14]可以在碱性磷酸酶(A l k a l i n e p h o s ph a t a s e ,A P )的水解作用下,释放出P O 4-P 被浮游植物及细菌利用㊂A P 作为一种典型的胞外酶,通常存在于水体㊁细胞周质内或细胞膜上[5],碱性磷酸酶活性(A l k a l i n e p h o s p h a t a s e a c t i v i t y,A P A )常被用来衡量海区内浮游生物受磷胁迫的状态Copyright ©博看网. All Rights Reserved.中国海洋大学学报2023年以及D O P的生物可利用性㊂根据过滤孔径和浮游生物粒径将A P A分为游离态(A P A f r e e,<0.2μm)及结合态,结合态被区分为浮游植物结合态(A P A p h y,> 3μm)和细菌结合态(A P A b a c,0.2~3μm)[15-18]㊂A P A f r e e来源复杂,游离于海水环境中,可在较长时间内保持活性[19-20],而位于细胞膜或细胞周质内的A P A p h y 和A P A b a c与细胞结合紧密[21],三者可以在磷限制条件下促进浮游植物和细菌对D O P的利用,对P的生物地球化学循环均有较大贡献㊂莱州湾作为渤海三大海湾之一,处于黄河三角洲高效生态经济区的核心区域和山东半岛城市群经济圈的腹地,其生态环境对山东省沿海社会经济的高质量发展具有重要支撑作用㊂而黄河口作为连接黄河与莱州湾㊁渤海湾的通道,其营养盐水平和透明度等要素显著影响莱州湾海域的生产力及浮游植物群落[22-23]㊂自1970年代末以来,随着工业化和城市化等人类活动高速增长,氮磷排放量大幅增加,导致莱州湾生态环境质量趋于恶化㊂1980年代初以来,已有众多学者对莱州湾氮㊁磷营养物质浓度及时空分布开展较为系统的研究,并基于P O4-P 浓度及D I NʒP O4-P比值对湾内营养盐限制状况进行了判定[24-26]㊂由于入湾总氮增幅远远高于总磷增幅[27],导致1990年代中期湾内的D I P大部分时段均处于较低水平,磷限制严重[11,28-31]㊂随着营养盐浓度和结构变化,湾内浮游植物群落结构也发生了显著的变化,突出表现在甲藻等磷需求低的物种及嗜氮性藻种明显增加,并在局部海域占据优势地位[22-23,31-32],浮游植物小型化趋势明显,如聚球藻等微微型浮游植物在海湾内被广泛检出[33-35],结果进一步导致了渔业资源衰退㊁生态系统服务功能下降等后果,已对环莱州湾地区高质量发展形成制约㊂目前,对莱州湾磷的赋存形态及时空分布特征缺乏系统了解,而且对海域磷限制背景下D O P的生物可利用性缺乏客观认知,导致对该海域磷的循环尤其对浮游生态系统磷的供给补充机制仍缺乏深入认知㊂因此,本研究通过2019 2021年在莱州湾开展的4次陆海同步调查,分析了黄河口及莱州湾海域不同形态磷的时空分布,以及陆源输入磷的季节变化特征;研究黄河口及莱州湾海域内不同碱性磷酸酶活性的季节变化,解析莱州湾浮游生物对低磷胁迫的响应㊂研究结果可为深化莱州湾磷的生物地球化学循环认知提供帮助㊂1材料与方法1.1调查区域及时间课题组分别于2019年5月10 12日(春季)㊁2019年8月18 21日(夏季)㊁2021年11月2 14日(秋季)和2021年3月22 27日(冬季)进行了4次陆海同步调查㊂其中,2019年8月航次时间处于强台风 利奇马 登陆山东[36]之后;2021年8月下旬至10月黄河中下游经历了历史上罕见的秋汛洪水[37],2021年11月航次处于秋汛之后㊂陆海同步调查站位如图1所示,海域大面站位覆盖了整个莱州湾及黄河口邻近海域;海域整体位于渤海南部,具有显著的大陆性气候持征,潮汐类型属不正规混合半潮,海域内水体交换能力较弱[38-41]㊂陆上监测站位分别布设于黄河㊁广利河㊁小清河等10条河流入海口及莱州湾污水处理厂(莱污处)和龙口市第二污水处理厂(龙二污)2个直排海口,径流量和污染物入海通量之和占到环莱州湾陆源排放总量的95%以上[42]㊂调查时间上,陆上流量监测及样品采集与海域大面站调查准同步进行,海域水文㊁化学和生物生态要素同步测定㊂图1黄河口及莱州湾海域调查站位图F i g.1S a m p l i n g s t a t i o n s o n l a n d-s e a s y n c h r o n o u s i n v e s t i g a t i o nf o r Y e l l o w R i v e r E s t u a r y(Y R E)a n d L a i z h o u B a y(L Z B)1.2样品采集及测定海水样品采集及测定参照‘海洋监测规范“(G B 17378 2007)和‘海洋调查规范“(G B/T12763 2007),水样经过孔径为0.45μm醋酸纤维滤膜过滤后,装入经酸洗的聚乙烯瓶中,与膜样一起冷冻保存(-20ħ),水样用于测定溶解态营养盐(主要包括N H4-N㊁N O3-N㊁N O2-N㊁P O4-P㊁S i O3-S i㊁D N㊁D P),膜样用于测定叶绿素a(C h l a)㊁悬浮颗粒物(S P M)及颗粒态营养盐㊂玻璃滤器预先经过2%~10%的盐酸浸泡24h,然后用超纯水清洗3~6次㊂环莱州湾河流和直排海口的样品按照‘地表水和污水监测技术规范标准(H J-T92002)“采集,并同步监测流量,水样盛于2L高密度聚乙烯塑料桶中冷藏保存(4ħ),6h内运回实验室中进行过滤和分析㊂主要入海口流量采用直读转子流速仪(Z S X-5,中国)测量㊂海水温度和盐度用多参数仪(M a n t a3.0, E u r e k a,美国)测定㊂营养盐通过营养盐自动分析仪(Q u A A t r o A p p l i c a t i o n s,德国)测定;N H4-N㊁N O3-N㊁N O2-N㊁P O4-P㊁S i O3-S i的检出限分别为0.03㊁0.02㊁0.01㊁0.01和0.14μm o l㊃L-1㊂溶解无机氮(D I N)为88Copyright©博看网. All Rights Reserved.11期段晓萌,等:黄河口及莱州湾海域磷的时空分布及浮游生物对低磷胁迫的响应N O3-N㊁N O2-N和N H4-N之和㊂C h l a用90%丙酮萃取后通过荧光法(F-4500,日本岛津)测定,检测限为0.01μg㊃L-1㊂S P M采用差减法测定㊂D N和D P采用碱性过硫酸钾氧化法测定[43]㊂D O N和D O P分别为D N和D I N㊁D P和P O4-P的差值㊂P N㊁P P通过将膜样经碱性过硫酸钾氧化法后测定[44]㊂T N㊁T P分别为D N和P N㊁D P和P P之和㊂A P A样品的采集及测定参照文献[15,45-46]㊂海水样品采集后分别用0.22和3μm孔径的针头式微孔滤膜过滤[47],分级过滤的海水和原位未过滤海水中分别加入荧光底物(4-甲基伞形酮磷酸酯,M U F-P),在原位海水温度下避光反应2h后,带回实验室用荧光光度计(F4700,日本岛津)测定样品的荧光强度(E x=365n m, E m=445n m),每个样品采集3个平行样[48]㊂A P A t o t a l 为总的A P A,以未过滤水样中A P A含量表示;A P A p h y 为浮游植物结合态的A P A,通过未过滤原位水样与3μm孔径膜上的A P A差值计算;A P A b a c表示浮游细菌结合态的A P A,以3μm孔径膜与0.22μm孔径膜上A P A的差值计算;A P A f r e e表示游离态的A P A,以经0.22μm孔径膜过滤的滤液中A P A含量表示㊂1.3数据处理及分析数据分析使用S P S S25.0软件进行;平面分布图使用S u r f e r15.0绘制;柱状图㊁散点图等使用O r i g i n2021绘制㊂2结果与讨论2.1黄河口及莱州湾海域陆源输入磷的季节变化所调查的入海河流和排污中,黄河流量最大[42],对陆源T P入海通量贡献也最大,各季节黄河口T P通量平均占比高达88.7%;小清河沿途受纳大量工业废水和固体废弃物,水质污染严重[49-50],其入海通量仅次于黄河,各季节平均占比为9.7%;广利河㊁虞河㊁白浪河㊁潍河入海通量占比分别为0.6%㊁0.3%㊁0.3%㊁0.2%,对湾内磷贡献相对较少;其余河流及直排污口磷的输入量极低,占比之和不足0.1%㊂陆源输入磷的赋存形态和入海通量均具有显著的季节变化特征㊂受径流量影响,夏季T P入海通量最高,合计达到6.52ˑ106m o l㊃d-1(见图2(b))㊂受高悬浮颗粒物含量影响,河流中P P为T P的主要存在形态[51-52],在T P中占比达到89%,其次为P O4-P,占比为7%,D O P仅占4%(见图2(d))㊂春季T P的入海通量为0.93ˑ106m o l㊃d-1(见图2(b)),磷的形态仍以P P为主,占比为66%,其次为D O P,占比为29%,远高于P O4-P,这可能是由于春季农业施用的有机肥较多,导致各入海口D P均以D O P为主;也可能是由于春季温度适宜,河道内浮游植物生长后聚集,大量的P O4-P在生物作用下被转化为D O P(见图2(c))㊂秋季径流量高于春季,而T P入海通量低于春季,为0.39ˑ106m o l㊃d-1 (见图2(b)),这主要是因为调查前环莱州湾经历了历时长且雨量大的降水过程[37],大量的磷已被冲刷入海;秋季仅针对黄河口进行了T P构成的调查,P赋存形态与夏季相似,以P P为主,占T P的83%,P O4-P㊁D O P 分别占15㊁2%(见图2(e))㊂冬季径流量较小,T P入海通量最低,仅为0.17ˑ106m o l㊃d-1(见图2(f));水体中磷赋存形态明显受到悬沙量的影响[53],明显区别于其他三个季节,P P占比降至17.9%,T P以D P为主,占比达81.7%,D P中以P O4-P为主(见图2(f))㊂图2黄河口及环莱州湾主要入海口(包括黄河㊁广利河㊁小清河㊁弥河㊁白浪河㊁虞河㊁潍河㊁胶莱河㊁莱州市污水处理厂㊁界河和龙口市污水处理厂的入海口)的流量(a)㊁T P入海通量(b)及不同形态P在T P中占比的季节变化((c)春季;(d)夏季;(e)秋季;(f)冬季㊂)F i g.2S e a s o n a l v a r i a t i o n o f r u n o f f(a),T P f l u x(b)a n d t h e p r o p o r t i o n s o f d i f f e r e n t f o r m s i n T P a t m a i n e n t r a n c e s o ft h e Y e l l o w R i v e r a n d L a i z h o u B a y((c)S p r i n g;(d)S u m m e r;(e)A u t u m n;(f)W i n t e r.)T h o s e e n t r c m r e s a r e l o c t e d i n Y e l l o w R i v e r,G u a n g l i R i v e r,X i a o q i n g R i v e r,M i R i v e r,B a i l i a n g R i v e r,Y U R i v e r,W e i R i v e r,J I a o l a i R i v e r,t h e s e w a g e p l a n t L a i z h o u R i v e r,J i e R i v e r,a n d t h e s e w a g e p l a n t i n L o n g k o u98Copyright©博看网. All Rights Reserved.中 国 海 洋 大 学 学 报2023年2.2黄河口及莱州湾海域中不同形态磷的时空分布及影响因素春季,莱州湾水体中T P 呈现近岸高㊁远岸低 的分布趋势,平均值为(1.35ʃ0.26)μm o ㊃L -1(见表1),最高值和最低值分别出现在潍河口和湾口东部海域(见图3q)㊂T P 中,P P 呈斑块状分布(见图3m ),整体浓度较低,平均值为(0.25ʃ0.09)μm o l ㊃L -1,在T P 中的平均占比为24%(见图4(b))湾内,这是由于春季径流量较小,河流运输的P P 主要积累于河口处[53]㊂D P 与T P 的分布趋势基本一致(见图3i ),平均值为(1.01ʃ1.05)μm o l ㊃L -1,在T P 中的平均占比为77%(见图4(b ))㊂D P 中,P O 4-P 呈现湾西南部高于其他区域的分布特征(见图3a ),小清河口附近有一明显高值区,与刘义豪等[26]的研究结果一致㊂D O P 也基本呈现近岸高远岸低分布特征(见图3e ),平均值为(0.91ʃ0.75)μm o l ㊃L -1,在T P 中的平均占比高达70%,是D P 的主要成分㊂P O 4-P ㊁D O P 均与盐度呈显著负相关(r =-0.846,P <0.01;r =-0.765,P <0.01),这表明D P 组成和分布明显受到陆源输入的显著影响[23,28,54-55]㊂浮游生物的生命活动也是D O P 的重要来源,春季温度适宜浮游植物生长,C h l a 却处于较低水平(见表1),这表明调查可能处于浮游植物消亡期,在此期间水体中大量P O 4-P 被大量消耗㊁D O P 被释放[56],图3 黄河口及莱州湾海水中不同形态磷的时空分布(a d ㊁e h ㊁i l ㊁m p ㊁q t :春㊁夏㊁秋㊁冬季P O 4-P ㊁D O P ㊁D P ㊁P P ㊁T P 的平面分布㊂)F i g .3 T e m p o r o s p a t i a l d i s t r i b u t i o n o f d i f f e r e n t p h o s p h o r u s i n s u r f a c e w a t e r s o f t h e Y e l l o w R i v e r a n d L a i z h o u B a yi n d i f f e r e n t s e a s o n s (a d ,e h ,i l ,m p :H o r i z o n t a l d i s t r i b u t i o n s o f P O 4-P ,D O P ,D P ,P P a n d T P i n i n s p r i n g ,s u m m e r ,a u t u m n a n d w i n t e r .)09Copyright ©博看网. All Rights Reserved.11期段晓萌,等:黄河口及莱州湾海域磷的时空分布及浮游生物对低磷胁迫的响应导致D O P 为D P 主要成分,P O 4-P 绝对受限面积高达81.5%,除湾西南部沿岸外大部分区域处于磷绝对限制㊂夏季T P 平均值为(0.97ʃ1.42)μm o l ㊃L -1(见表1),P P ㊁D P 平均值分别为(0.46ʃ0.60)μm o l ㊃L -1㊁(0.51ʃ0.93)μm o l ㊃L -1(见表1),两者水平相当,且分布趋势均与T P 的一致㊂湾东部养殖区附近D P 和P P 浓度也较高(见图3r),这表明海水养殖也是影响湾内磷浓度及分布的重要因素,海水养殖不仅会直接增加海域内的磷负荷[57-58],也会通过下行调控作用影响浮游植物对磷的积累和消耗㊂D P 以P O 4-P 为主,P O 4-P 的平均值为(0.39ʃ0.96)μm o ㊃L -1(见表1),P O 4-P 在小清河口有一梯度较大的高浓度水舌,这主要是由于 利奇马 台风导致沿岸形成以风浪为主的海浪[36],海浪的作用下沿岸沉积物再悬浮作用增强,暴雨行洪导致小清河内大量的污染物和底泥被冲刷入海,使得大量P O 4-P 被释放于水体中[59],P O 4-P 绝对受限面积显著减少㊁D I NʒP O 4-P 明显降低,湾内的磷限制得到缓解;D O P 仅为(0.12ʃ0.06)μm o l ㊃L -1(见表1),远低于P O 4-P ,无明显的空间分布特征(见图3f )㊂秋季T P 平均值为(0.64ʃ0.42)μm o l ㊃L -1(见表1),在小清河口附近存在高值区(见图3s )㊂T P 构成与夏季相似,D P 与P P 水平相当,平均值分别为(0.31ʃ0.31)㊁(0.33ʃ0.17)μm o l ㊃L -1(见表1),均在小清河口存在高浓度水舌(见图3k ㊁o )㊂D O P 平均值为(0.26ʃ0.20)μm o l ㊃L -1(见表1),平面分布特征与D P 一致(见图3g );P O 4-P 平均值仅为(0.09ʃ0.16)μm o l ㊃L -1(见表1),远低于D O P ,湾内P O 4-P 仅在小清河口附近浓度较高(见图3c ),浮游植物在高浓度营养盐的刺激下迅速生长后将P O 4-P 转化为P P 后又以D O P 的形式释放于水体中,进而导致海域内D P 以D O P 为主㊂此外,秋汛期间,黄河水体流速快㊁泥沙含量高[59-60],大量的无机氮被汇入湾内,导致P O 4-P 迅速被吸收利用,湾内D I N ʒP O 4-P 高达R e d f i e l d 比值的69倍,P O 4-P 绝对限制面积高达87.45%,这表明调水期间输入大量高氮㊁低磷的淡水,加重了莱州湾海域的磷限制和营养盐失衡的程度[61]㊂冬季T P 的平均值为(0.49ʃ0.34)μm o l ㊃L -1(见表1),在黄河口附近的浓度较高(见图3t );T P 以P P为主(见图4(b )),P P 平均值为(0.33ʃ0.26)μm o l ㊃L -1(见表1),在黄河口处存在高值(见图3p );P O 4-P ㊁D O P ㊁D P 均处于较低水平,平均浓度分别为(0.09ʃ0.10)㊁(0.07ʃ0.05)㊁(0.16ʃ0.11)μm o l ㊃L -1(见表2;图3d ㊁h ㊁l)㊂磷的形态㊁浓度及分布受浮游植物生长调控,P P 与C h l a 间呈显著正相关(r =-0.915,P <0.01),表明浮游植物吸收P O 4-P 转化为P P ㊂由于受陆源输入㊁浮游生物生消过程㊁沉积物悬浮释放等多重因素的影响,调查海域内磷形态和分布具有显著差异性㊂降雨等自然因素㊁农业施肥及调流调沙等人类活动的共同作用,环莱州湾各条河流的磷负荷及赋存形态有明显的季节差异[53,62]㊂丰水期河流的入莱州湾通量在全年中占绝对优势[63],磷在河口区的吸附解吸及生物的吸收转化等过程导致陆源输入T P的季节变化与海域内的并不一致㊂海域内T P 基本呈现出春季>夏ʈ秋季>冬季的特征(见图4(a))㊂2.3黄河口及莱州湾海域内浮游生物的低磷胁迫响应A P A 是衡量海水生态系统对D O P 利用的重要指标,磷限制条件下,浮游植物及浮游细菌通过合成㊁分泌出大量的A P 利用D O P 来应对低磷胁迫[64-66]㊂调查海域水体中均检测到了A P A ,这表明调查期间莱州湾海域浮游植物及细菌的生长㊁繁殖在很大程度上受到了磷的胁迫作用㊂图4 黄河口及莱州湾海水中T P 浓度(a )及构成(b)的季节变化F i g .4 S e a s o n a l v a r i a t i o n o f c o n c e n t r a t i o n (a )a n d c o m po s i t i o n (b )o f t o t a l p h o s p h o r u s i n s u r f a c e w a t e r s o f t h e Y e l l o w R i v e r a n d L a i z h o u B a y19Copyright ©博看网. All Rights Reserved.中 国 海 洋 大 学 学 报2023年表1 不同季节黄河口及莱州湾水文参数及各项营养盐要素T a b l e 1 H y d r o l o g i b a l pa r a m e t e r s a n d n u t r i e n t e l e m e n t s i n d i f f e r e n t s e a s o n s i n Y R E a n d L Z B 参数P a r a m e t e r统计值S t a t i s t i c 春季S p r i n g夏季S u m m e r秋季A u t u m n冬季W i n t e rT /ħ平均值①16.80ʃ2.3126.18ʃ1.1612.88ʃ3.636.74ʃ1.79范围②12.23~22.9323.56~27.885.45~17.013.84~12.10S平均值①27.43ʃ4.9126.12ʃ7.1722.72ʃ3.6727.64ʃ4.55范围②0.01~31.340.94~32.6911.15~29.484.24~31.25C h l a /(μg ㊃L -1)平均值①1.20ʃ0.986.82ʃ6.075.56ʃ6.196.30ʃ5.92范围②N .D .~4.130.95~22.100.18~27.490.37~24.94S P M /(m g ㊃L -1)平均值①41.60ʃ47.43 61.83ʃ177.8113.78ʃ14.6324.05ʃ10.30范围②15.94~254.068.50~10620.28~67.8011.55~52.13D I N /(μm o l ㊃L -1)平均值①38.88ʃ23.9145.06ʃ51.0943.50ʃ41.0328.16ʃ68.97范围②12.31~129.431.02~208.8111.48~140.212.45~440.94D O N /(μm o l ㊃L -1)平均值①123.08ʃ131.4445.23ʃ118.9025.22ʃ26.649.68ʃ4.26范围②1.03~747.931.03~747.934.58~142.622.93~27.04D N /(μm o l ㊃L -1)平均值①152.67ʃ148.6078.38ʃ128.4267.29ʃ46.8838.51ʃ72.42范围②40.07~877.3613.69~877.3619.85~282.839.00~467.99P N /(μm o l ㊃L -1)平均值①3.59ʃ1.5057.93ʃ52.196.44ʃ3.033.86ʃ1.75范围②0.73~8.140.73~237.391.79~14.402.07~12.40T N /(μm o l ㊃L -1)平均值①159.05ʃ152.70137.91ʃ136.7073.74ʃ47.4342.37ʃ73.89范围②42.01~877.3620.70~877.3623.32~290.6912.10~480.39P O 4-P /(μm o l ㊃L -1)平均值①0.19ʃ0.400.39ʃ0.960.09ʃ0.160.09ʃ0.10范围②0.02~1.810.02~4.950.01~0.860.02~0.51D O P /(μm o l ㊃L -1)平均值①0.91ʃ0.750.12ʃ0.060.26ʃ0.200.07ʃ0.05范围②0.45~3.950.02~0.22 0.07~1.20 N .D .~0.26D P /(μm o l ㊃L -1)平均值①1.01ʃ1.050.51ʃ0.93 0.31ʃ0.310.16ʃ0.11范围②0.15~5.760.05~4.950.11~2.060.06~0.62P P /(μm o l ㊃L -1)平均值①0.25ʃ0.090.46ʃ0.600.33ʃ0.170.33ʃ0.26范围②0.06~0.460.07~3.420.08~0.840.15~1.73T P /(μm o l ㊃L -1)平均值①1.35ʃ0.260.97ʃ1.420.64ʃ0.420.49ʃ0.34范围②0.62~1.450.18~6.300.30~2.820.23~2.35D I N ʒP O 4-P 平均值①711.47ʃ423.11452.88ʃ500.211083.64ʃ1043.69363.69ʃ308.48范围②71.51~1547.0517.61~2632.45116.35~5348.0216.56~1248.97P N ʒP P平均值①14.34ʃ6.66284.24ʃ277.1320.12ʃ4.1512.37ʃ1.80范围②N .D .~45.77 3.05~1056.5810.33~29.037.16~15.09A P A p h y/(n m o l ㊃L -1㊃h -1)平均值① 715.10ʃ1592.604.63ʃ9.43范围② 3.75~7568.500.05~58.67A P A b a c/(n m o l ㊃L -1㊃h -1)平均值① 55.04ʃ286.231.50ʃ1.73范围② N .D .~1786.25N .D .~7.00 A P A f r e e/(n m o l ㊃L -1㊃h -1)平均值① 25.72ʃ61.199.16ʃ4.10 范围② 0.25~327.000.02~16.33A P A t o t a l /(n m o l㊃L -1㊃h -1)平均值① 795.86ʃ1749.8715.28ʃ11.74范围②5.25~8091.500.07~75.50注:表中 N .D . 表示未检出, 表示未采集㊂ N .D . i n t h e t a b l e m e a n s n o t c h e c k e d o u t ,a n d m e a n s n o t c o l l e c t e d .①M e a n V a l u e ;②R a n ge .29Copyright ©博看网. All Rights Reserved.11期段晓萌,等:黄河口及莱州湾海域磷的时空分布及浮游生物对低磷胁迫的响应秋季,A P A t o t a l 基本呈现出湾南部近岸高㊁远岸低的特征(见图5d ),A P A p h y 与AP A f r e e 的平面分布特征与A P A t o t a l 相似(见图5a ㊁c ),A P A b a c 则在湾东部养殖区附近存在一高值(见图5b )㊂A P A t o t a l 平均含量(795.86ʃ1749.87)n m o l㊃L -1㊃h -1(见表1),其中,A P A p h y 贡献最大,在AP A t o t a l 中的平均占比为85%,其次为A P A b a c ,平均占比为12%,A P A f r e e 占比最小,仅为3%㊂这表明,秋季莱州湾在较长时间内处于严重受限状态[67],浮游植物及细菌合成㊁分泌出的A P 会大量聚集在细胞周围,快速利用D O P [16]㊂冬季A P A t o t a l 分布与秋季类似,在白浪河㊁弥河口有一高值,但黄河口附近较低(见图5h );A P A p h y 在弥河口㊁白浪河口存在一小范围高值区(见图5e );A P A f r e e则在黄河口出现低值区(见图5g );A P A b a c 整体处于较低水平(见图5f )㊂A P A t o t a l 均值为(15.28ʃ11.74)n m o l ㊃L -1㊃h -1(见表1),仅为秋季的1/50(见图6(a )),与长江口[68]㊁黄海[47]等海域A P A 处于同等水平,这可能是由于冬季D I N ʒP O 4-P 比值低㊁磷受限程度轻,A P 被大量表达;另外秋季温度远高于冬季,有利于A P 的表达,进一步导致秋季A P A 远高于冬季㊂A P A t o t a l 中,A P A f r e e 在A P A t o t a l 中占比最高,均值为62%,A P A p h y次之,占比均值为27%(见图6(b )),这表明,磷受限程度相对较轻时,浮游植物及细菌对细胞周质内和细胞膜上A P 的依赖程度会明显降低㊂此外,浮游生物种类是影响A P 表达的直接因素[69-71],秋㊁冬季节A P A t o t a l及构成的差异也可能是由浮游生物种类不同所致㊂(a d :秋季,e h :冬季㊂a d :A u t u m n ,e h :W i n t e r .)图5 黄河口及莱州湾海域中不同类型A P A 的季节变化F i g .5 S e a s o n a l v a r i a t i o n o f d i f f e r e n t s i z e f r a c t i o n s o f a l k a l i n e p h o s p h a t a s e a c t i v i t y (A P A )i n s u r f a c e w a t e r o f t h e Y e l l o w R i v e r a n d L a i z h o uB a y图6 黄河口及莱州湾海域中A P A t o t a l (a )及其构成(b )的变化F i g .6 S e a s o n a l v a r i a t i o n o f A P A t o t a l (a )a n d i t s c o m p o s i t i o n (b )o f t h e Y e l l o w R i v e r a n d L a i z h o u B a y 研究表明,海洋浮游生态系统中A P 的分泌显著受水体中P O 4-P 含量的影响,P O 4-P 主要通过 诱导 抑制 机制调节A P 的活性,即P O 4-P 缺乏状态下浮游生物会诱导产生大量碱性磷酸酶,在很大程度上接触并水解D O P ;当P O 4-P 浓度较高时酶的活性受到抑制[72-75]㊂对于黄河口及莱州湾海域(见图7),当P O 4-P 39Copyright ©博看网. All Rights Reserved.中 国 海 洋 大 学 学 报2023年低于阈值0.05μm o l ㊃L -1时,不同类型的A P A 均显著增加,表明低磷环境下浮游植物和细菌会产生大量A P 来利用D O P 以维持自身的新陈代谢;当P O 4-P 高于阈值时,浮游生物对磷的需求基本得到满足,A P A 维持在较低水平,A P 的分泌受到抑制㊂本文的P O 4-P 阈值与大西洋和太平洋等海域的(0.030μm o l㊃L -1)较为接近[73];然而,马来西亚半岛沿海水域和河口区P O 4-P 的阈值则高达0.9μm o l ㊃L -1[74],表明不同海域调控A P A 的P O 4-P 有较大差异㊂这可能是由于不同海域浮游生态系统以及温度㊁营养盐等环境因子的差异所致,具体原因还需要进一步深入研究㊂黄河口及莱州湾海域中P O 4-P 阈值明显低于P O 4-P 绝对受限阈值(0.1μm o l ㊃L -1),表明A P A 作为衡量磷限制的指标更为灵敏[11]㊂图7 黄河口及莱州湾海域中碱性磷酸酶活性与P O 4-P 之间的关系F i g .7 T h e r e l a t i o n s h i p b e t w e e n d i f f e r e n t A P A P O 4-P i n s u r f a c e w a t e r o f t h e Y e l l o w R i v e r a n d L a i z h o u B a y3 结论(1)黄河口对T P 总通量的贡献最大,小清河次之;受径流量影响,入海口T P 通量呈现出夏季>春季>秋季>冬季的季节变化;春㊁夏㊁秋季入海T P 均以P P 为主,春季D P 以D O P 为主,夏㊁秋季以P O 4-P 为主;冬季T P 以D P 为主,D P 以P O 4-P 为主㊂(2)海域内T P 浓度及构成受陆源输入和浮游生物活动共同调控,呈现出春季>夏季ʈ秋季>冬季的季节变化㊁近岸高远岸低的分布特征,高值区基本位于黄河口及小清河口㊂T P 中,春季以D P 为主,夏㊁秋季P P 与D P 相当,冬季以P P 为主;D P 中,春㊁秋季以D O P 为主,夏㊁冬季以P O 4-P 为主㊂海域内浮游植物普遍受到P O 4-P 的绝对限制和相对限制,夏季 利奇马 台风带来的强降雨及风浪使海域内磷限制得到缓解;黄河秋汛可能向湾内汇入大量的氮,加快了P O 4-P 的消耗,磷限制程度被加重㊂(3)海域内A P A 处于较高水平,秋季A P A t o t a l 主要由A P A p h y 贡献,冬季A P A t o t a l 主要由A P A f r e e 贡献;当水体中D I N ʒP O 4-P 极高㊁P O 4-P 严重缺乏时,A P A p h y ㊁A P A b a c 较高,浮游植物及细菌主要依赖细胞周质及细胞膜上的A P 来水解D O P 以维持新陈代谢㊂(4)P O 4-P 通过诱导-抑制机制对A P 进行调控,A P A 随P O 4-P 的增加而降低,当P O 4-P 低于0.05μm o l ㊃L -1时,浮游植物迅速分泌出大量的碱性磷酸酶来应对低磷胁迫;当高于阈值时,A P A 维持在较低水平;湾内浮游植物为适应磷限制环境可能已降低了对磷的需求㊂参考文献:[1] B j ör k m a n K M ,K a r l D M.B i o a v a i l a b i l i t y o f d i s s o l v e d o r ga n i c p h o s p h o r u s i n t h e e u ph o t i c z o n e a t s t a t i o n A L O H A ,N o r t h P a c i f i c S u b t r o p i c a l G y r e [J ].L i m n o l o g y a n d O c e a n o g r a p h y,2003,48(3):1049-1057.[2] D y h r m a n S T ,R u t t e n b e r g K C .P r e s e n c e a n d r e gu l a t i o n o f a l k a -l i n e p h o s p h a t a s e a c t i v i t y i n e u k a r y o t i c p h y t o pl a n k t o n f r o m t h e c o a s t a l o c e a n :I m p l i c a t i o n s f o r d i s s o l v e d o r g a n i c p h o s p h o r u s r e m i -n e r a l i z a t i o n [J ].L i m n o l o g y a n d O c e a n o g r a p h y,2006,51(3):1381-1390.[3] D y h r m a n S ,A m m e r m a n J ,V a n M o o y B.M i c r o b e s a n d t h e m a r i n e p h o s p h o r u s c y c l e [J ].O c e a n o g r a p h y (W a s h i n gt o n ,D .C .),2007,20(2):110-116.[4] C u r r i e D J ,K a l f f J .T h e r e l a t i v e i m p o r t a n c e o f b a c t e r i o pl a n k t o n a n d p h y t o p l a n k t o n i n p h o s p h o r u s u p t a k e i n f r e s h w a t e r [J ].L i m -n o l o g y a n d O c e a n o g r a p h y,1984,29(2):311-321.[5] C h r ós t R J .E n v i r o n m e n t a l C o n t r o l o f S y n t h e s i s a n d A c t i v i t y of A q u a t i c M i c r o b i a l E c t o e n z y m e s [M ].N e w Y o r k :S p r i ng e r -V e r l a g,1991:29-54.[6] C o n t e r J B ,W e t z e l R G .U p t a k e o f d i s s o l v e d i n o r g a n i c a n d o r ga n i c p h o s p h o r u s c o m p o u n d sb y p h y t o p l a n k t o n a n d b ac t e r i o pl a n k t o n [J ].L i m n o l o g y a n d O c e a n o g r a p h y,1992,37(2):232-243.[7] C o t n e r J B ,A m m e r m a n J W ,P e e l e E R ,e t a l .P h o s ph o r u s l i m i t e d b a c t e r i o p l a n k t o n g r o w t h i n t h e S a r g a s s o S e a [J ].A q u a t i c M i c r o b i a l E c o l o g y,1997,13:141-149.[8] L o m a s M W ,B u r k e A L ,L o m a s D A ,e t a l .S a r ga s s o S e a p h o s -p h o r u sb i o g e oc h e m i s t r y :A n i m p o r t a n t r o l e f o rd i s s o l ve d o r ga n i c p h o s p h o r u s (D O P )[J ].B i o ge o s c i e n c e s ,2010,7:695-670.[9] L o m a s M W ,B o n a c h e l a J A ,L e v i n S A ,e t a l .I m pa c t o f o c e a n p h y t o p l a n k t o n d i v e r s i t y o n p h o s p h a t e u p t a k e [J ].P r o c e e d i n gs o f t h e N a t i o n a l A c a d e m y o f S c i e n c e s ,2014,111(49):17540-17545.[10] M a l f a t t i F ,T u r k V ,T i n t a T ,e t a l .M i c r o b i a l m e c h a n i s m s c o u -p l i n g c a r b o n a n d p h o s p h o r u s c y c l e s i n p h o s ph o r u s -l i m i t e d n o r t h -e r n A d r i a t i c S e a [J ].S c i e n c e o f T h e T o t a l E n v i r o n m e n t ,2014,470-471:1173-1183.[11] X i n M ,W a n g B ,X i e L ,e t a l .L o n g -t e r m c h a n ge s i n n u t r i e n t r e -g i m e s a n d t h e i r e c o l o gi c a l e f f e c t s i n t h e B o h a i S e a ,C h i n a [J ].M a -r i n e P o l l u t i o n B u l l e t i n ,2019,146:562-573.[12] B j o r k m a n .B i o a v a i l a b i l i t y o f i n o r g a n i c a n d o r g a n i c p h o s ph o r u s c o m p o u n d s t o n a t u r a l a s s e m b l a g e s o f m i c r o o r ga n i s m s i n H a w a i i a n 49Copyright ©博看网. All Rights Reserved.11期段晓萌,等:黄河口及莱州湾海域磷的时空分布及浮游生物对低磷胁迫的响应c o a s t a l w a t e r s[J].M a r i n e E c o l o g y P r o g r e s s S e r i e s,1994,111:265-273.[13] Y o u n g C L,I n g a l l E D.M a r i n e d i s s o l v e d o r g a n i c p h o s p h o r u sc o m p o s i t i o n:I n s i g h t s f r o m s a m p l e s r e c o v e r ed u s i n g c o m b i ne de l e c t r o d i a l y s i s/r e v e r s e o s m o s i s[J].A q u a t i c G e o c h e m i s t r y,2010,16(4):563-574.[14] P a y t a n A,M c l a u g h l i n K.T h e o c e a n i c p h o s p h o r u s c y c l e[J].C h e m i c a l R e v i e w s,2007,107(2):563-576.[15] H o p p e.P h o s p h a t a s e a c t i v i t y i n t h e s e a[J].H y d r o b i o l o g i a,2003,493:187-200.[16] H u a n g B,O u L,W a n g X,e t a l.A l k a l i n e p h o s p h a t a s e a c t i v i t y o fp h y t o p l a n k t o n i n E a s t C h i n a S e a c o a s t a l w a t e r s w i t h f r e q u e n th a r m f u l a l g a l b l o o m o c c u r r e n c e s[J].A q u a t i c M i c r o b i a l E c o l o g y,2007,49:195-206.[17] N a u s c h M.A l k a l i n e p h o s p h a t a s e a c t i v i t i e s a n d t h e r e l a t i o n s h i p t oi n o r g a n i c p h o s p h a t e i n t h e P o m e r a n i a n B i g h t(s o u t h e r n B a l t i c S e a)[J].A q u a t i c M i c r o b i a l E c o l o g y,1998,16:87-94. [18] O u L,H u a n g B,L i n L,e t a l.P h o s p h o r u s s t r e s s o f p h y t o p l a n k-t o n i n t h e T a i w a n S t r a i t d e t e r m i n e d b y b u l k a n d s i n g l e-c e l l a l k a-l i n e p h o s p h a t a s e a c t i v i t y a s s a y s[J].M a r i n e E c o l o g y P r o g r e s s S e-r i e s,2006,327:95-106.[19] T h o m s o n B,W e n l e y J,C u r r i e K,e t a l.R e s o l v i n g t h e p a r a d o x:C o n t i n u o u s c e l l-f r e e a l k a l i n e p h o s p h a t a s e a c t i v i t y d e s p i t e h i g hp h o s p h a t e c o n c e n t r a t i o n s[J].M a r i n e C h e m i s t r y,2019,214:1-6.[20] T h o m s o n B,W e n l e y J,L o c k w o o d S,e t a l.R e l a t i v e i m p o r t a n c eo f p h o s p h o d i e s t e r a s e v s.p h o s p h o m o n o e s t e r a s e(a l k a l i n e p h o s p h a-t a s e)a c t i v i t i e s f o r d i s s o l v e d o r g a n i c p h o s p h o r u s h y d r o l y s i s i n e p i-a n d m e s o p e l a g i c w a t e r s[J].F r o n t i e r s i n E a r t h S c i e n c e,2020,8:1-8.[21] K a m e n n a y a N A,G e r a k i K,S c a n l a n D J,e t a l.A c c u m u l a t i o n o fa mb i e n t p h o s p h a t e i n t o t h e p e r i p l a s m o f m a r i n e b ac t e r i a i s p r o t o nm o t i v e f o r c e d e p e n d e n t[J].N a t u r e C o m m u n i c a t i o n s,2020,11(1):1-14.[22]栾青杉,康元德,王俊.黄河口邻近海域浮游植物群落及其多样性的长期变化(1960 2010)[J].中国水产科学,2017,24(5): 913-921.L u a n Q i n g s h a n,K a n g Y u a n d e,W a n g J u n.L o n g-t e r m c h a n g e s o f p h y t o p l a n k t o n c o m m u n i t y a n d d i v e r s i t y i n a d j o i n i n g w a t e r s o f t h e Y e l l o w R i v e r e s t u a r y(1960 2010)[J].J o u r n a l o f F i s h e r y S c i-e n c e s of C h i n a,2017,24(5):913-921.[23]苏芝娟,王玉珏,董志军,等.调水调沙后黄河口邻近海域浮游植物群落响应特征[J].海洋学报,2015,37(4):62-75.S u Z h i j u a n,W a n g Y u y u,D o n g Z h i j u n,e t a l.R e s p o n s e o f p h y t o-p l a n k t o n a s s e m b l a g e s t o t h e w a t e r-s e d i m e n t r e g u l a t i o n i n t h e a d-j a c e n t s e a o f t h e Y e l l o w R i v e r m o u t h[J].H a i y a n g X u e b a o,2015, 37(4):62-75.[24]夏斌,张晓理,崔毅,等.夏季莱州湾及附近水域理化环境及营养现状评价[J].渔业科学进展,2009,30(3):103-111.X i a B i n,Z h a n g X i a o l i,C u i Y i,e t a l.E v a l u a t i o n o f t h e p h y s i c o-c h e m i c a l e n v i r o n m e n t a nd n u t r i t i o n s t a t u s i n L a i z h o u B a y a n d a d-j a c e n t w a t e r s i n s u m m e r[J].P r o g r e s s i n F i s h e r y S c i e n c e s,2009, 30(3):103-111.[25]由丽萍,赵玉庭,孙珊,等.2018年春季和夏季莱州湾营养盐结构及限制特征[J].渔业科学进展,2021,42(6):15-24.Y o u L i p i n g,Z h a o Y u t i n g,S u n S a n,e t a l.C h a r a c t e r i s t i c s o f n u-t r i e n t s t r u c t u r e s a n d l i m i t a t i o n s i n L a i z h o u B a y i n t h e s p r i n g a n d s u m m e r o f2018[J].P r o g r e s s i n F i s h e r y S c i e n c e s,2021,42(6): 15-24.[26]张亮,宋春丽,张乃星,等.莱州湾西部海域营养盐分布特征及营养化程度评价[J].广西科学院学报,2017,33(2):82-86.Z h a n g L i a n g,S o n g C h u n l i,Z h a n g N a i x i n g,e t a l.N u t r i e n t d i s t r i-b u t i o n a n d e u t r o p h ic a t i o n a s s e s s m e n t i n w e s t s e a a r e a o f L a i z h o uB a y[J].J o u r n a l o f G u a n g x i A c a d e m y o f S c i e n c e s,2017,33(2):82-86.[27]张锦峰,高学鲁,李培苗,等.莱州湾西南部海域及其毗邻河流水体营养盐的分布特征及长期变化趋势[J].海洋通报,2015,34(2):222-232.Z h a n g J i n f e n g,G a o X u e l u,L i P e i m i a o,e t a l.N u t r i e n t d i s t r i b u-t i o n c h a r a c t e r i s t i c s a n d l o n g-t e r m t r e n d s i n t h e s o u t h w e s t o f t h e L a i z h o u B a y a n d i t s a d j a c e n t r i v e r s[J].M a r i n e S c i e n c e B u l l e t i n, 2015,34(2):222-232.[28]蒋红,崔毅,陈碧鹃,等.渤海近20年来营养盐变化趋势研究[J].海洋水产研究,2005(6):61-67.J i a n g H o n g,C u i Y i,C h e n B i j u a n,e t a l.T h e v a r i a t i o n t r e n d o f n u t r i e n t s a l t s i n t h e B o h a i S e a[J].M a r i n e F i s h e r i e s R e s e a r c h, 2005(6):61-67.[29]刘义豪,杨秀兰,靳洋,等.莱州湾海域营养盐现状及年际变化规律[J].渔业科学进展,2011,32(4):1-5.L i u Y i h a o,Y a n g X i u l a n,J i n Y a n g,e t a l.D i s t r i b u t i o n a n d i n t e r a-n n u a l v a r i a t i o n o f n u t r i e n t s i n L a i z h o u B a y[J].P r o g r e s s i n F i s h-e r y S c i e n c e s,2011,32(4):1-5.[30]赵玉庭,苏博,李佳蕙,等.2013年春季莱州湾海域理化环境及水质状况分析[J].渔业科学进展,2016,37(4):74-80.Z h a o Y u t i n g,S u B o,L i J i a h u i,e t a l.E v a l u a t i o n o f p h y s i c o c h e m-i c a l e n v i r o n m e n t a n d w a t e r q u a l i t y i n t h e L a i z h o u B a y i n S p r i n g o f2013[J].P r o g r e s s i n F i s h e r y S c i e n c e s,2016,37(4):74-80.[31]孙军,刘东艳,徐俊,等.1999年春季渤海中部及其邻近海域的网采浮游植物群落[J].生态学报,2004,24(9):2003-2016.S u n J u n,L i u D o n g y a n,X u J u n,e t a l.T h e n e t z-P h y t o p l a n k t o nc o m m u n i t y o f t h e c e n t r a l B o h a i S e a a nd i t s a d j a ce n t w a t e r s i ns p r i n g,1999[J].A c t a E c o l o g i c a S i n i c a,2004,24(9):2003-2016.[32]宋秀凯,程玲,付萍,等.莱州湾金城海域网采浮游植物年际变化及与环境因子的关系[J].生态学报,2019,39(8):2784-2793.S o n g X i u k a i,C h e n g L i n g,F u P i n g,e t a l.C h a n g e s o f n e t-c o l l e c t-e d p h y t o p l a n k t o n c o m m u n i t y i n J i n c h e n g a r e a of L a i z h o u B a y a n di t s r e l a t i o n s h i p w i t h e n v i r o n m e n t a l f a c t o r[J].A c t a E c o l o g i c a S i n-i c a,2019,39(8):2784-2793.[33]王彩霞.渤海海域微生物群落结构的时空变化及其对环境压力的响应[D].烟台:中国科学院烟台海岸带研究所,2018.W a n g C a i x i a.S p a t i a l a n d T e m p o r a l V a r i a t i o n o f M i c r o b i a l C o m-m u n i t y S t r u c t u r e i n t h e B o h a i S e a A n d I t s R e s p o n s e t o E n v i r o n-m e n t a l[D].Y a n t a i:Y a n t a i I n s t i t u t e o f C o a s t a l Z o n e R e s e a r c h,C h i n e s e A c a d e m y o f S c i e n c e s,2018.[34]杨琳.渤海微微型浮游植物的时空分布[D].青岛:中国海洋大学,2012.Y a n g L i n.T h e S p a t i a l a n d T e m p o r a l D i s t r i b u t i o n o f P i c o p h y t o-p l a n k t o n i n B o h a i S e a[D].Q i n g d a o:O c e a n U n i v e r s i t y o f C h i n a, 2012.59Copyright©博看网. All Rights Reserved.。
1000 0569/2022/038(01) 0091 08ActaPetrologicaSinica 岩石学报doi:10 18654/1000 0569/2022 01 07江南造山带万古金矿床含金硫化物组构与金沉淀机制沈关文1 张良1 孙思辰1,2 宇天伟1 李增胜3 吴圣刚4 陈俊辉4 申颖3SHENGuanWen1,ZHANGLiang1 ,SUNSiChen1,2,YUTianWei1,LIZengSheng3,WUShengGang4,CHENJunHui4andSHENYing31 中国地质大学地质过程与矿产资源国家重点实验室,北京 1000832 福建省地质矿产勘查开发局,福州 3500033 山东省地质科学研究院,自然资源部金矿成矿过程与资源利用重点实验室,济南 2500134 湖南黄金洞矿业有限责任公司,岳阳 4145071 StateKeyLaboratoryofGeologicalProcessesandMineralResources,ChinaUniversityofGeosciences,Beijing100083,China2 FujianBureauofGeologyandMineralResourcesExplorationandDevelopment,Fuzhou350003,China3 MNRKeyLaboratoryofGoldMineralizationandResourceUtilization,ShandongAcademyofGeologicalSciences,Jinan250013,China4 HunanHuangjindongMingCoLtd,Yueyang414507,China2021 08 01收稿,2021 11 24改回ShenGW,ZhangL,SunSC,YuTW,LiZS,WuSG,ChenJHandShenY 2022 Texturesofgold bearingsulfidesandgoldprecipitationmechanism,Wangugolddeposit,JiangnanOrogen ActaPetrologicaSinica,38(1):91-108,doi:10 18654/1000 0569/2022 01 07Abstract TheWangugolddeposit,withaprovengoldresourceof~85t,islocatedinthemiddlesectionoftheJiangnanOrogenandoccursinmetamorphicrockseriesoftheNeoproterozoicLengjiaxiGroup ItformedunderthecontroloftheNNE NE trendingChangsha PingjiangfaultzoneandtheEW trendingJiuling Qingshuiductileshearzone Themainoretypesofthedepositincludearsenopyrite ,pyrite ,sericite ,andquartz alteredslateandquartz sulfideveins,followedbyslatebrecciawithinhydrothermalquartz Arsenopyriteandpyritearethemaingoldbearingmineralsinthedeposit,whicharewidelydistributedinthedeposit Goldmineralizationcanbedividedintofourstages:I,themilkyquartz muscovite scheelite;II,thesmokygrayquartz muscovite arsenopyrite pyrite gold;III,thesmokygrayquartz muscovite pyrite arsenopyrite polymetallicsulfide gold;andIV,themilkyquartz calcite Amongthem,theIIandIIIarethemainmineralizationstages BasedontheresultsofElectronProbeMicroAnalysis(EPMA)ofarsenopyritefromthemainmineralizationstages,thecontentsofAsinarsenopyriteatstageIIrangefrom42 19%to44 84%,withanaverageof43 42%(n=56).ThecontentsofAsinarsenopyriteatstageIIIrangefrom40 08%to43 36%,withanaverageof42 08%(n=19).Accordingtothephasediagramofarsenopyritethermometer,theformationtemperatureofApy 1inthestageIIisestimatedtobe364±21℃withasulfurfugacityvaryingwithin10-9 7~10-7 TheformationtemperatureofApy 2inthestageIIIis319±22℃,anditssulfurfugacityis10-11 5~10-8 6 Thecontentsofinvisiblegoldingold bearingarsenopyriteandpyritewere0 01%~0 66%and0 01%~0 11%,respectively,revealedbyEPMAdata TheAu Asdataofpyriteareplottedonbothsidesofthegoldsolubilitycurve,indicatingthatgoldinpyritemainlyexistsintheformofnano scaleparticlesandsolidsolutionorlatticegold TheproportionofnanometergoldparticlesinPy 1pyriteis73 33%,morethanthatinPy 2(67 80%).Theabovedataindicatethatintheprocessofwater rockreaction,theiron bearingmineralsinsurroundingrocksreactwithH2Sinore formingfluidtoformarsenopyriteandpyrite Accompaniedbyastrongwater rockreaction,themaingold bearingsulfideschangedfromarsenopyritetopyriteduringmineralizationfromstageIItoIIIwiththedecreaseofmineralizationtemperatureandsulfurfugacity,i e ,strongsulfurizationresultsininstabilityofthegold sulfurcomplexandthereleaseofgold Asaresult,goldisdepositedintothesulfidelattice本文受国家重点研发计划(2019YFA0708603)、国家自然科学基金项目(41702070)、高等学校学科创新引智计划(BP0719021)、中央高校基本科研业务费项目(2 9 2020 044)和中国地质大学地质过程与矿产资源国家重点实验室专项基金(MSFGPMR201804)联合资助.第一作者简介:沈关文,男,1998年生,硕士生,资源与环境专业,E mail:sgwxue24@163.com通讯作者:张良,男,1988年生,副教授,硕士生导师,主要从事矿物学与矿床学教学和科研工作,E mail:zhangliangcugb@126.combysubstitutionorintheformofmicroscopic ultramicroscopicgoldparticlestoformgold bearingsulfide Therefore,sulfidationisthemainmechanismoftheprecipitationofinvisiblegoldinsulfidesattheWangugolddepositKeywords Occurrencestateofgold;Mineralthermometer;Goldprecipitationmechanism;Wangugolddeposit;JiangnanOrogen摘 要 万古金矿床位于江南造山带中部,赋存于新元古界冷家溪群浅变质岩系中,受NNE NE向长沙 平江断裂带和近EW向九岭 清水韧性剪切带联合控制,金资源量约85t。
◆综 述◆黄酮类中药单体干预Nrf2/HO -1通路治疗糖尿病微血管并发症研究进展刘映君1,阴永辉2,王文宽1,张金涛1,孔 畅1(1.山东中医药大学,山东 济南 250355; 2.山东中医药大学附属医院,山东 济南 250011)[摘要] 糖尿病微血管并发症是糖尿病最主要的慢性并发症。
研究表明,核转录因子红系2相关因子2/血红素加氧酶-1(Nrf2/HO -1)信号通路是治疗糖尿病微血管并发症的重要靶点。
黄酮、黄酮醇、二氢黄酮、异黄酮、二氢异黄酮、查耳酮、黄烷醇、花青素等黄酮类中药单体可以通过调控Nrf2/HO -1信号通路发挥缓解氧化应激、减少炎症反应、抑制细胞凋亡、减轻纤维化、改善血流动力学等作用,从而达到治疗糖尿病视网膜病变、糖尿病心肌病、糖尿病肾病、糖尿病周围神经病变等多种糖尿病微血管并发症的目的。
参考文献66篇。
[关键词] 糖尿病微血管并发症;核转录因子红系2相关因子2/血红素加氧酶-1信号通路;黄酮类中药单体;氧化应激;炎症反应;细胞凋亡;纤维化;血流动力学[中图分类号] R259.872 [文献标志码] A [文章编号] 1007-659X (2024)02-0246-08DOI :10.16294/ki.1007-659x.2024.02.019Research Progress of Flavonoid Monomer in Treating Diabetic Microangiopathy by Regulating Nrf2/HO -1 Signaling PathwayLIU Yingjun 1,YIN Yonghui 2,WANG Wenkuan 1,ZHANG Jintao 1,KONG Chang 1(1.Shandong University of Traditional Chinese Medicine ,Jinan 250355,China ;2.Affiliated Hospital of Shandong University of Traditional Chinese Medicine ,Jinan 250011,China )Abstract Diabetic microangiopathy is the most important chronic complication of diabetes mellitus (DM ). Studies have shown that nuclear factor -erythroid 2-related factor 2 /heme oxygenase -1(Nrf2/HO -1) signaling pathway is an important target for the treatment of diabetic microangiopathy. Flavonoids ,flavonols ,dihydro⁃flavonoids ,isoflavones ,dihydroisoflavones ,chalcones ,flavanols ,anthocyanins and other flavonoid monomers of Chinese materia medica can alleviate oxidative stress ,reduce inflammatory response ,inhibit apoptosis ,alleviatefibrosis and improve hemodynamics ,etc. by regulating Nrf2/HO -1 signaling pathway ,so as to achieve the purpose of treating diabetic retinopathy ,diabetic cardiomyopathy ,diabetic nephropathy ,diabetic peri -pheral neuropathy and other diabetic microvascularcomplications. There are 66 references in total.Keywords diabetic microangiopathy ;nuclear factor -[收稿日期] 2023-11-15[基金项目] 国家自然科学基金青年科学基金项目(编号:82104727);山东省自然科学基金项目(编号:ZR2021MH088)[作者简介] 刘映君(1997—),女,山东寿光人,2021年级硕士研究生,研究方向:中医药防治内分泌代谢性疾病。
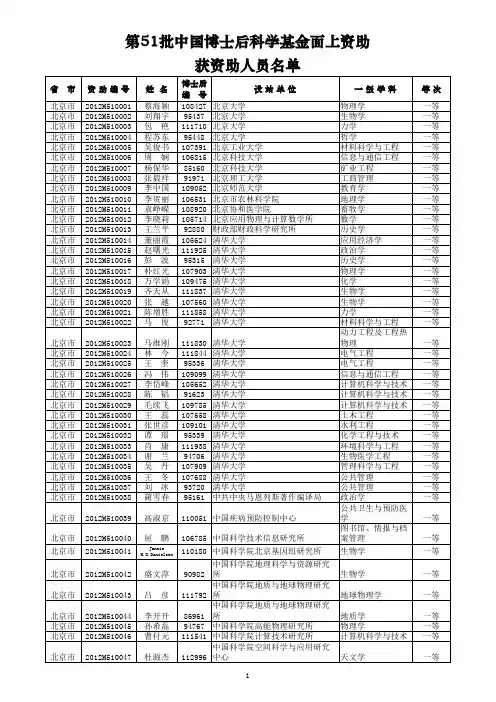
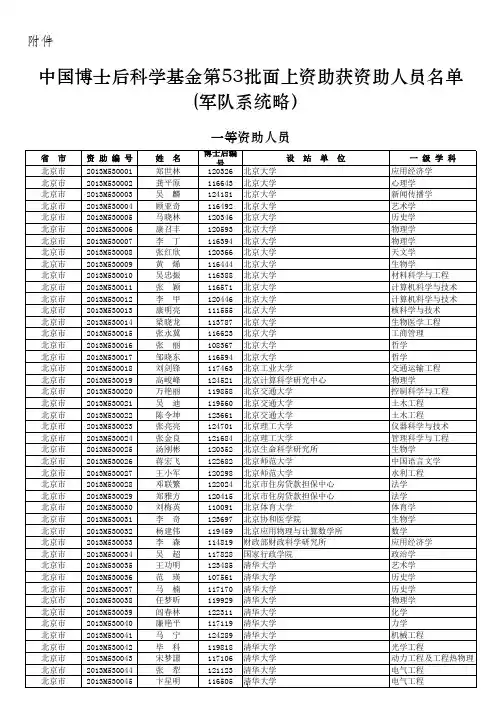
2013年第54批中国博士后基金面上资助人员名单2013年第54批中国博士后基金面上资助人员名单随着科技的不断发展和创新的推动,博士后研究成为各大科研机构和高校中广泛且重要的职位。
为了鼓励和支持博士后的科研工作,中国科学院和中国工程院联合设立了中国博士后科研资助计划。
每年,该计划都会公布一批博士后基金面上资助人员名单,以表彰他们在各个领域的杰出贡献。
在本文中,我将深入探讨2013年第54批中国博士后基金面上资助人员名单,并介绍他们的成就以及对相关领域的贡献。
2013年第54批中国博士后基金面上资助人员名单是一个标志性的名单,因为它代表着当时中国科学研究领域中顶尖人才的集结和荣誉。
这些博士后具有丰富的科研经验和出色的研究成果,在各自的领域中取得了重要的突破和创新。
他们的研究涵盖了物理学、化学、生物医学、工程技术、信息技术等多个领域,为我国科学研究的发展做出了重要贡献。
在物理学领域中,2013年第54批中国博士后基金面上资助人员名单中有许多杰出的博士后。
以王杰为例,他的研究成果在纳米科学和纳米技术领域取得了重要突破。
他通过独特的材料设计和合成方法,成功制备了具有优异电池性能的纳米材料,为新能源的研究和应用带来了重要的启示。
他的论文不仅在高水平学术期刊上发表,还被其他研究人员广泛引用,为同行们提供了宝贵的研究参考。
化学领域也有一批优秀的博士后在2013年第54批中国博士后基金面上资助人员名单中脱颖而出。
以陈华为例,他的研究集中在新型有机合成方法的开发和应用上。
他提出了一种基于多组分反应的新型合成策略,能够高效地合成复杂结构的有机化合物。
他的研究成果为有机合成领域提供了有力的工具和方法,为天然产物的合成以及医药和材料科学的发展做出了重要贡献。
生物医学研究领域也有许多值得关注的博士后在2013年的名单中获得资助。
以刘慧为例,她主要研究针对肿瘤的新型治疗方法。
她在肿瘤免疫治疗领域做出了重要贡献,通过独特的药物设计和开发,成功开发出一种针对特定癌症类型的新型免疫治疗药物。