
*中国科学院知识创新项目,KZCX3-SW -214号。吴玉霖,研究员,E -mail:YLWu@https://www.doczj.com/doc/c93053498.html,
收稿日期:2005-04-22,收修改稿日期:2005-06-30
环境长期变化对胶州湾
浮游植物群落结构的影响*
吴玉霖 孙 松 张永山
(中国科学院海洋研究所海洋生态与环境科学重点实验室 青岛 266071)
提要 主要依据1991年5月)1995年11月和1997年2月)2004年11月连续多年每季度月(2、5、8、11月)采用/浅水ó型0浮游生物网在胶州湾10个站位自海底至表垂直拖取的浮游植物样品,同时搜集自1954年以来约50年有关胶州湾环境因子及浮游植物的历史监测资料,分析研究胶州湾环境因子长期变化及其对浮游植物群落结构的影响。结果表明,近50年来由于大规模人类活动给胶州湾环境带来很大影响,大量工农业生产废水和生活污水源源不断进入湾内,造成水域营养盐含量迅速增加,近3年来NO 3-N 和NH 4-N 比1962年分别提高14.5倍和9.5倍,沿岸带的开发导致胶州湾水域面积不断缩小,纳潮量下降,水域环境容量和自净能力减弱,水域环境趋向退化。环境因子的变化对胶州湾浮游植物群落结构造成明显的影响,近50年来,浮游植物数量呈增加趋势;优势种更替明显,原先一些优势种不再居优势地位,而比较喜肥的种类如中肋骨条藻数量显著增加,成为最重要的优势种;浮游植物多样性指数有所下降,表明胶州湾浮游植物群落处于演化之中,同时由于营养盐含量迅速增加,为赤潮的形成与发展提供了丰富的物质基础,自1997年以来,赤潮几乎连年暴发,并有愈演愈烈的发展趋势,反映出胶州湾生态环境已呈现异常的征兆。关键词 胶州湾,环境变化,浮游植物,群落结构中图分类号 P593
浮游植物个体微小,种类繁多,不同种类对环境的适应能力有一定差异,因而受环境因子变化的影响,浮游植物群落结构诸如种类组成、丰度等可能出现某些变化。作为海洋中最主要初级生产者的浮游植物在海洋生态系统中具有重要功能,因此开展环境长期变化对浮游植物群落结构的演变及演化方向的研究成为海洋科学研究的热点之一,国内外已有若干相关报道(沈志良,2002;吴玉霖等,2004;赵淑江等,2001a;Zhao et al ,2005;Copper et al ,1993;Andren,1999;Patsch et al ,1997;Smayda,1984)。本文中作者利用1953年以来约50年对胶州湾浮游植物调查结果,结合自然因素及人类活动引起的环境长期变化资料进行综合分析,探讨浮游植物对环境长期变化的响应。
1 调查材料与方法
1.1 浮游植物调查资料
1.1.1 1953年3月)1956年2月对青岛近海浮游矽藻的调查(李冠国等,1956) 在栈桥设采样点,每星期用标准25号筛绢(网目为64L m)制作的小网采样1次。
1.1.2 1981年2月)1981年11月对胶州湾浮游植物的调查(郭玉洁等,1992) 湾内共设调查站位11个,湾外3个(图1),每季度(2、5、8、11月)采样1次,以浅水ó型网从海底至表垂直拖取。1.1.3 1991年5月)1995年11月、1997年2月)2004年11月对胶州湾浮游植物进行的连续调查(作者) 在湾内设调查站位9个,湾外1个(图2)。采样时间、间隔、网具及方法同1.1.2。
第36卷 第6期海 洋 与 湖 沼
Vol.36,No.62005年11月
OCE ANOLOGIA E T LIMNOLOGIA SINIC A
Nov.,2005
图1 1981年采样站位
Fig.1 Location of sampling stations in
1981
图2 1992)2004年采样站位
Fig.2 Location of sampling stations(1992)2004)
对胶州湾浮游植物多样性采用Shannon -Wiener 指数(H c )进行分析,其公式为:
H c =-E S
i=1
P i log Z P i
其中:H c 为生物多样性指数,S 为样品中的种类总数,P i 为第i 种的个体数与总个体数的比值。1.2 环境因子的历史资料
水文气象、海水化学资料来自中国生态系统
研究网络(CE RN)胶州湾生态系统研究站数据库,部分营养盐资料由沈志良研究员提供。
海水养殖及其它人类活动的相关资料搜集自青岛市统计局所编的5统计数据61949)2003年年度资料。
2 结果和讨论
2.1 胶州湾浮游植物分布与种类组成的一般特征
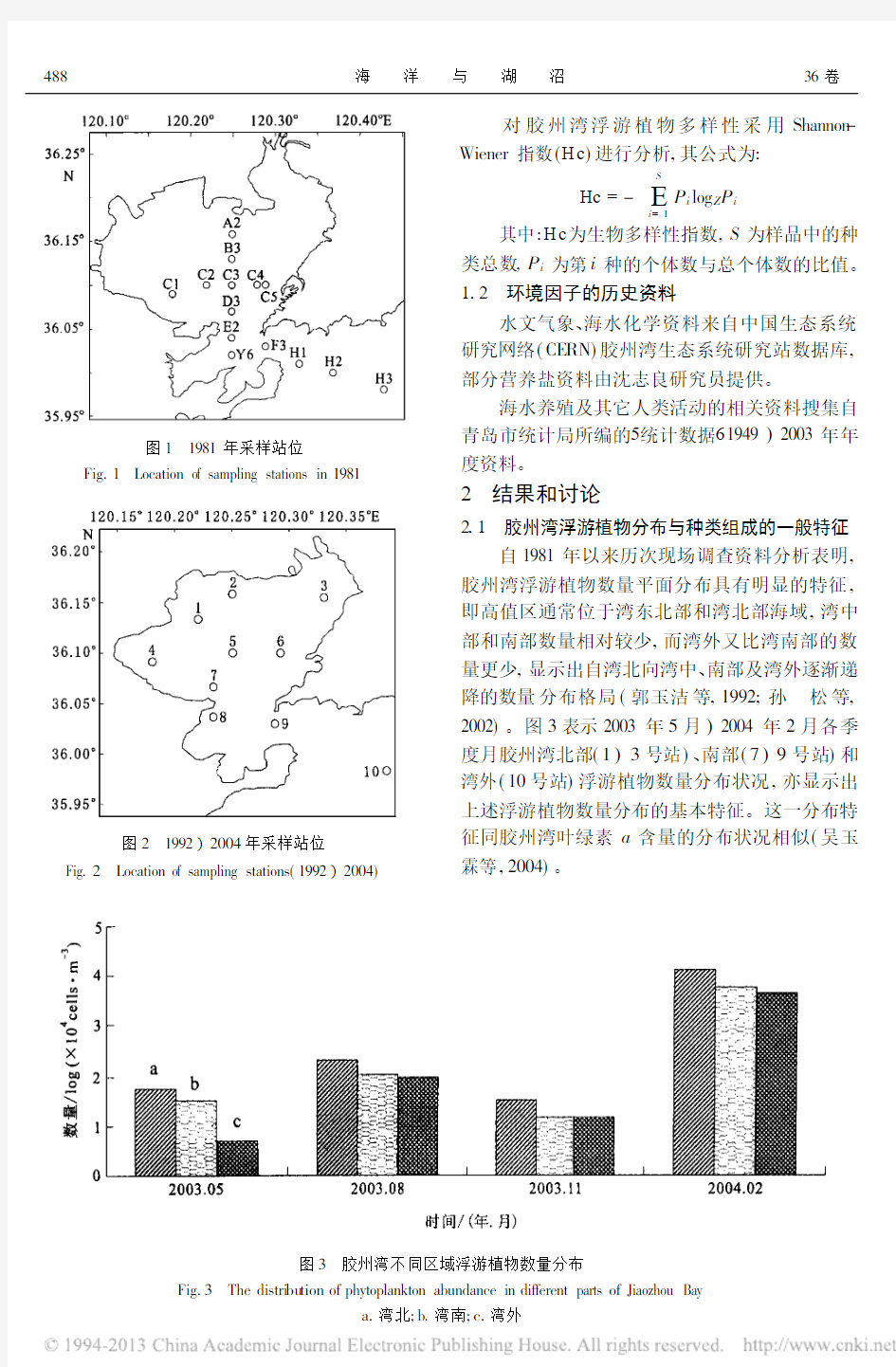
自1981年以来历次现场调查资料分析表明,胶州湾浮游植物数量平面分布具有明显的特征,即高值区通常位于湾东北部和湾北部海域,湾中部和南部数量相对较少,而湾外又比湾南部的数量更少,显示出自湾北向湾中、南部及湾外逐渐递降的数量分布格局(郭玉洁等,1992;孙 松等,2002)。图3表示2003年5月)2004年2月各季度月胶州湾北部(1)3号站)、南部(7)9号站)和湾外(10号站)浮游植物数量分布状况,亦显示出
上述浮游植物数量分布的基本特征。这一分布特征同胶州湾叶绿素a 含量的分布状况相似(吴玉霖等,2004)
。
图3 胶州湾不同区域浮游植物数量分布
Fig.3 The distri bu tion of phytoplankton abundance in different parts of Jiaozhou Bay
a.湾北;
b.湾南;
c.湾外
488 海 洋 与 湖 沼36卷
胶州湾浮游植物这种分布格局同营养盐的分布状况基本一致。表层海水中NO 3-N 和NO 2-N 浓度均是北部和东北部高、由北向南递减、湾外最低;NH 4-N 、PO 4-P 和SiO 3-Si 表、底层浓度均为东北部和东部高,自东北向西南降低,而湾外低于湾内(沈志良等,1994;沈志良,2002),表明氮、磷等营养盐对浮游植物数量分布具有相当重要的影响。另外,湾东北部水体稳定度相对较高亦有利于浮游植物聚集。
胶州湾地处暖温带,四季分明,湾内浮游植
物有着明显的季节数量动态变化特征,数量高峰通常出现在冬季,夏季常有一个次高峰(图4)。春、秋季数量较少,这符合温带海域一年有双峰的一般现象。如若把1954至今的13个年份调查加以平均,则冬季高达1119.01@104cells/m 3,夏季为690.08@104cells/m 3,春、秋季分别为160.47@104
cells/m 3
和203.77@104
cells/m 3
。浮游植物这种数量季节变化特征同胶州湾叶绿素a 含量季节变化状况基本一致(吴玉霖等,2004)
。
图4 不同季节浮游植物数量的年际变化Fig.4 The seasonal variation in phytoplankton abundance
a.2月;
b.5月;
c.8月;
d.11月
关于胶州湾浮游植物种类组成,1981年调查共鉴定116种,其中硅藻100种,甲藻15种,金藻1种(郭玉洁等,1992);1999年调查共鉴定117种,其中属硅藻104种,甲藻12种及金藻1种(孙 松等,2002);2003年调查共鉴定120种,其中属硅藻类的有95种,甲藻21种,绿藻2种,金藻和裸藻各1种。大部分浮游植物种类属于暖温带性和广布性种类,夏、秋季有若干热带近岸性种类出现,如波状石丝藻(Litho desmium undulatum )等。此外近岸偶尔能采到半咸水性种。中肋骨条藻(Skeletonema costatum )为胶州湾最重要的优势种。其它较重要的种类还有加氏星杆藻(Asterionella kariana)、日本星杆藻(A.japonica)、旋链角毛藻(Chaetoceros curvisetus )、尖刺拟菱形藻(Pseudonitzschia pungens)、奇异菱形藻(Nitzschia paradoxa )、星脐圆筛藻(Coscinodiscus asterom phalus)及短角弯角藻(Eucam pia zoodiacus)等。胶州湾浮游植物多样性指数H c 平面分布状况与数量分布的格局明显不同,湾北部多样性指数通常较低。2003年1)3号站平均生物多样性指数为0.780,而湾南部(7)9号站)平均值为0.926,湾外(10号站)平均为0.936。湾南
部和湾外比较接近(图5)。这反映出三部分水域浮游植物植物群落结构有所差异。造成这种
局面的主要原因是湾东北部和北部营养盐含量较高,水质肥沃,水体相对较为稳定,导致部分喜肥的种类如中肋骨条藻大量繁殖,往往成为浮游植物密集区。因生物多样性指数同群落中生物种类数及生物丰度关系密切,所以湾北部浮游植物多样性指数相对较低。湾南部及湾外处于湾内水同外海水的混合带,浮游植物种类相对多些,是造成这两部分水域多样性指数较高的主要原因。
2.2 胶州湾浮游植物的长期变化
2.2.1 浮游植物数量的长期变化 图4和图6表示1954年及其后若干年份各季度月与年平均浮游植物数量动态变化状况。总体上看,近50年来胶州湾浮游植物数量有呈增加的变化趋势,但年际间波动较大。进一步分析表明,可以划分两个不同的演变阶段,即1981)1997年浮游植物数量呈逐渐下降阶段和1998年至今为迅速增长时期。浮游植物数量这种年际变化趋势可能同胶州湾大规模滤食性贝类筏式养殖业发展状况有关。
6期吴玉霖等:环境长期变化对胶州湾浮游植物群落结构的影响489
图5 胶州湾浮游植物多样性指数分布
Fig.5 The distribution of phytoplankton di versity index H c in Jiaozhou Bay
a.湾北;
b.湾南;
c.
湾外
图6 胶州湾浮游植物数量的长期变化
Fig.6 Long -term changes of phytoplank ton abundance in Jiaozhou Bay
胶州湾自80年代中期起迅速发展贝类筏式养殖,至90年代中期达到高峰,这期间浮游植物数量明显地呈现出逐年下降的变化趋势,反映出滤食性贝类对浮游植物数量有很强的控制作用(详细分析见3.3.2节)。1997年开始,因病害等原因,扇贝大规模灾难性死亡,筏式养殖业出现大滑坡并迅速萎缩,1998年后胶州湾浮游植物数量得到恢复并呈迅速增加的态势。胶州湾浮游植物数量动态变化比较容易受人类活动如海水养殖及城市排污等影响的一个重要原因是因为该湾水域面积较小,约为374.4km 2,平均水深仅7m,几乎为青岛市区所包围,湾口狭小,与湾外水交换不很顺畅;营养盐含量持续增加,导致某些浮游植物迅速繁殖且时而形成赤潮。1999年浮游植物平均数量之
所以高达2393.8@104cells/m 3
,就是因为该年夏季先后出现短角弯角藻和中肋骨条藻赤潮的缘故(详细分析见/2.30节)。
2.2.2 浮游植物种类组成的变化 胶州湾各年四个季度月调查所获的浮游植物种类数见图7,可以看出,年际间有一定的波动,但没有明显的长期变化趋势。以1997年出现的种类最多,各季度月均在65种以上。另外2003年四个季度平均种数亦达70种。应该说明的是,季度月所采到的浮游植物种类数同调查站位的多少之间有一定关系,调查站位多,不同浮游植物种类被采到的几率就高,反之则低。1954年仅在栈桥设1个站,该年各季度月调查所获种类数约为其它年份的一半。
490 海 洋 与 湖 沼36卷
图7 四个季度月浮游植物种类数年际变化
Fig.7 Annual variation in phytoplankton species number in di fferent seasons
a.2月;
b.5月;
c.8月;
d.11月
近50年来,胶州湾浮游植物优势种构成的变化是比较明显的(表1),中肋骨条藻是最常见的优势种,在上个世纪50年代和80年代的冬季或夏季常成为优势种群,而进入90年代至今,该藻数量有明显增加,且在一年中的大部分季节常居优势地位,2001年夏季,全湾该藻平均密度高达28059@103cells/m 3,优势度达到96.7%;90年代以来,加氏星杆藻和诺登海链藻(Thalassiosira nordenskioldii)常在冬季大量出现,成为重要优势种,如1999年2月,加氏星杆藻平均数量高达19714@103cells/m 3,优势度为97.6%,而诺登海链藻在2002年冬季全湾平均数量达到20243@103cells/m 3,优势度为32.6%;近几年来圆筛藻属
的星脐圆筛藻在夏季有较大量出现,成为优势种,2003年7月曾在胶州湾北部急速增殖而形成赤潮;进入90年代后,尖刺拟菱形藻和奇异菱形藻常在春、秋季较多量出现,由于春、秋两季胶州湾浮游植物数量相对较少,这两种硅藻成为重要的优势种。同上述那些90年代新出现的优势种呈鲜明对比的是,原先(1954年和1981年)胶州湾的若干浮游植物优势种,诸如窄隙角毛藻(C h.a ffi -nis)、菱形海线藻(Thalassionema nitzschioides )、刚毛根管藻(Rhizosolenia setigera )和辐射圆筛藻(Cos.radiacus)等数量相对下降,已不再成为优势种群,显示出胶州湾浮游植物群落在种类构成上存在长期变化的重要现象。
表1 胶州湾浮游植物优势种长期变化
Tab.1 Long -term changes of dominant phytoplankton species in Jiaozhou Bay
年 份冬 季
春 季
夏 季
秋 季
1954
Coscinodiscus asteromphalus (25.6%)
T halassione ma nitzschioides (16.7%)
A ste rionella j a p on ica(17.6%)Rhizosolenia stolterfothii (33.3%)
Coscinodiscus radiatus (12.5%)
Skeletonema costatum (79.3%)
Coscinodiscus asteromphalus (16%)
Chaetoceros a ffinis (9.3%)
1981
Chaetoceros a ffinis (12.8%)Chaetoceros ompressus (11.9%)
Chaetoceros densus(40.3%)
Rhizosolenia setigera(10.3%)Clmacodium biconcavum
(9.6%)
Skeletonema costatu m (25.4%)
Asterionella j a p onica (11.9%)
Chaetoceros a ffinis (10.7%)
1992
Skeletonema costa tum (40.4%)
Rhizosolenia alata f .indica (17.9%)
Asterionella j a ponica (36.8%)
Coscinodiscus granii(88.4%)Biddulphia aurita(21.5%)
Chaetoceros debilis (20.3%)Pseudonitzschia pungens (14.3%)
6期吴玉霖等:环境长期变化对胶州湾浮游植物群落结构的影响491
续表年份冬季春季夏季秋季
1993Rhizosolenia delicatula
(70.1%)
A ste rionella j a p on ica(8.9%)Pseu don it z schia pungens
(69.5%)
Chaetoceros curvisetus
(62%)
Rhizosolen ia styli f ormis
(12.6%)
Chaetoceros compressus
(10.5%)
1994Rhizosolenia delicatula
(96.2%)Asterionella j a ponica
(36.6%)
Pseudonitzschia pun gens
(30.9%)
Skeletonema costatum
(22.3%)
Chaetoceros curvisetus
(15.9%)
Skeletonema costatu m
(32.1%)
Coscinodiscus asteromphalus
(18.4%)
Eucampia zoodiacus(11%)
1995Skeletonema costa tum
(46.3%)
A ste rionella kariana(22.5%)Ditylu m brigh twellii(30.6%)
Coscinodiscus perforatus
(24%)
Chaetoceros curvisetus
(71.9%)
Actinocyclus ehrenbergii
(12.7%)
Rhizosolen ia styli f ormis
(30.3%)
Asterionella j a p onica
(15.7%)
1997Chaetoceros com pressus
(46.3%)
Chaetoceros densus(14.5%)Pseu don it z schia pungens
(28.4%)
Rhiz osolenia fr a gilissima
(17.6%)
Skeletonema costatum
(17.2%)
Skeletonema costatu m
(73.5%)
1998A ste rionella kariana(57.8%) Skeletonema costa tum
(30.9%)Asterionella ka riana(74.4%)
Ske letonema costatum
(21.3%)
Actinocyclus ehrenbergii
(75.7%)
Skeletonema costatum
(13.2%)
Coscinodiscus perf oratus
(39.7%)
Chaetoceros curvisetus
(10.8%)
1999A ste rionella kariana(97.6%)Chaetoceros cu rvisetus
(40%)
Actinoc yclus ehrenbergii
(17.4%)
Eucam p ia zoodiacus(16.7%)Skeletonema costatum
(91.8%)
Euca mpia z oodiacus(7%)
Rhizosolen ia styli f ormis
(19%)
Chaetoceros loren zianus
(14.2%)
Skeletonema costatu m
(13.3%)
2000Pseudonitzschia pungens
(45.3%)
Eucampia zoodiacus(24.2%)
A ste rionella j a p on ica(14%)Chaetoceros densus(74.3%)Chaetoceros curvisetus
(81.3%)
Coscinodiscus wailesii(9.5%)
Skeletonema costatu m
(57.2%)
Nit z schia p a r a doxa(16.6%)
2001T halassiosira nordenskioldii
(76.9%)
Skeletonema costa tum
(18.1%)Pseu don it z schia pungens
(65.2%)
Chaetoceros eibenii(24.8%)
Skeletonema costatum
(96.7%)
Skeletonema costatu m
(55.1%)
Tha lassiosira rotula(30.1%)
2002T halassiosira nordenskioldii
(32.6%)
Skeletonema costa tum(56%)Ske letonema costatum
(15.5%)
Chaetoceros debilis(13.5%)
Nitzschia p a radoxa(11.5%)
Coscinodiscus asteromphalus
(79.4%)
Chaetoceros curvisetus
(11.3%)
Rhizosolen ia delicatula
(18.5%)
Pseudonitzschia pungens
(15.2%)
Nit z schia p a r a doxa(14%)
Skeletonema costatu m
(11.2%)
492海洋与湖沼36卷
续表
年 份冬 季
春 季
夏 季
秋 季
2003
Skeletonema costa tum (71.3%)
Chaetoceros deci p iens (27.3%)
Asterionella j a ponica (24.5%)
Ske letonema costatum (14.5%)
Chaetoceros curvisetus (21.4%)
Coscinodiscus asteromphalus (37.1%)
Nit z schia p a r a doxa(27.1%)Skeletonema costatu m (11.8%)
Chaetoceros loren zianus (11.1%)
注:括号内数据为该藻数量占浮游植物总量的百分比
2.2.3 浮游植物多样性的长期变化 生物群落是一定生境里各生物种群相互依存和相互制约的组合结构单元,某些生存环境因子的变化可能影响这种组合结构特征的改变。为了解胶州湾浮游植物群落结构演化状况,应用Shannon -Wiener 生物多样性指数对胶州湾浮游植物群落稳定度进行分析。该指数的主要依据是群落中物种丰度和生物种群丰度。从1954年以来历次浮游植物调查资料进行生物多样性指数统计结果(图8)可以看出,几十年来,胶州湾浮游植物多样性指数有下
降趋势,1981年平均多样性指数为1.094,而90年代至今3个时间段(1991)1995、1997)2000、2001)2003)分别平均为0.758、0.733和0.694。应该指出的是,1954年因仅设置1个站位进行采样,代表性差些,如以1981年及90年代以来的3个时间段的浮游植物多样性指数变化状况进行比较,四个季度月的变化趋势基本一致,均有所下降,但以冬季最为明显。生物多样性指数呈下降趋势反映出胶州湾浮游植物群落结构受环境因子
变化的影响已处于不断演化之中。
图8 胶州湾浮游植物多样性指数H c 长期变化
Fig.8 Long -term changes of phytoplankton diversity index H c in di fferent seasons
a.2月;
b.5月;
c.8月;
d.11月
2.2.4 胶州湾的赤潮 赤潮是一大海洋灾害,
是在异常的环境条件由某种或几种浮游植物(有时由原生动物或细菌)暴发性增殖或高度聚集引起水体变色的生态异常现象。自上世纪70年代起,在我国沿海发生的频率、规模和危害程度有愈演愈烈的发展趋势(吴玉霖等,2001),然而在1990年之前,胶州湾未见过发生赤潮的报道,1990年6月首次发现由原生动物中缢虫(Meso dinium rubrum)引起的面积约8万m 2
的小范围赤潮。但是自90年代中期开始,胶州湾赤潮迅猛发展,几乎年年发生,甚至一年内发生多次,赤潮原因种由
早先的中缢虫增加了6种硅藻和1种甲藻,赤潮面积亦越来越大,2004年3月赤潮面积达70km 2(表2)。在我国北方海域包括渤海和我国的边缘海)))黄海,已记录到的赤潮均发生在夏季和秋季,而胶州湾2004年冬季就暴发赤潮实属异常。国内外对赤潮成因研究结果表明,水体中大量营养盐的存在为赤潮形成提供重要物质基础,适宜的水文气象因子是促成赤潮暴发的一个重要条件(周成旭等,2000)。由此说明,近10年来胶州湾水域赤潮频频发生是与水域生态环境的变化密切相关。
6期吴玉霖等:环境长期变化对胶州湾浮游植物群落结构的影响493
表2 胶州湾赤潮发生状况Tab.2 Even ts of red tide in Jiaozhou Bay
时 间海域和面积 原因种
1990年6月22号锚地附近,8万m 2红色中缢虫(Mesodinium rubrum)1997年8月胶州湾中部中肋骨条藻(Skeletonema costatum)
1998年7月3)8日姑山附近海域中肋骨条藻和高贵盒形藻(Biddulphia regia)1999年6月中旬胶州湾东北部
短角弯角藻(Eucam pia z oodiacus)1999年7月小港、团岛嘴、沧口水道,26k m 2中肋骨条藻和短角弯角藻2000年7月
胶州湾中部,2km 2夜光藻(Noctiluca scintillans )2001年7月8)12日胶州湾口,9.8km 2红色中缢虫2003年7月4)11日团岛、大麦岛红色中缢虫
2003年7月胶州湾北部星脐圆筛藻(Cos.asterom pha lus)2004年2上旬胶州湾东北部
柔弱根管藻(Rh.delicatula)
2004年3月25日
胶州湾北部,面积约70km 2
诺登海链藻(Th.nordenskioldii)和中肋骨条藻
2.3 主要环境因子的长期变化及影响
近50年来,青岛市工农业生产迅速发展,市区人口不断增加,进入90年代以来,社会经济发展更是日新月异。2002年全市工农业总产值达到2848.29亿元,是1965年的118.3倍,市区人口亦由1965年的147万人增加到2002年的242万人。随着市区人口不断增加和大规模经济活动的开展,大量生活污水和工农业生产产生的废水源源不断地进入胶州湾,造成水域环境发生退化,营养化水平不断增高,导致浮游植物群落结构的演化。
2.3.1 营养盐 据收集到的调查资料显示,自1962年来,胶州湾营养盐含量发生了很大变化(图9)。90年代NO 3-N 和NH 4-N 含量分别是60
年代的7.26倍和5.1倍,同期PO 4-P 的平均含量亦提高了2.43倍。近3年(2001)2003年)NO 3-N 和NH 4-N 含量又比上世纪90年代几乎翻了一番。
2001)2003年SiO 3-Si 平均浓度为5.79L mol/L,亦比90年代的平均值高约1倍。可以看出,近50年来,胶州湾氮、磷和硅等营养盐呈迅速增加的态势。近3年全湾平均无机氮含量高达22.40L mol/L,PO 4-P 含量为0.54L mol/L 远高于DIN=1L mol/L 和PO 4-P=0.1L mol/L 的浮游植物生长阈值(Justic et al ,1995),亦超过我国一类海水水质标准(GB.3097-97)。水域的富营养化为浮游植物大量繁殖提供重要的物质基础,是造成浮游植物数量趋向增加的主要原因,亦成为导致近10年来胶州湾赤
潮的发生并呈愈演愈烈趋势的关键因素。
图9 胶州湾营养盐含量的长期变化
Fig.9 Lon g -term changes in PO 4-P,Si O 3-Si,NO 3-N,NH 4-N and DIN concentration in Jiaozhou Bay
a.PO 4-P;
b.SiO 3-Si;
c.NO 3-N;
d.NH 4-N;
e.DIN
494 海 洋 与 湖 沼36卷
依据青岛市统计局所编的5统计数据61949)2003年年度资料,1997年青岛市废水排放总量为18545.2万t,到2002年就增加到23291.41万t,其中仅氨氮的排放量就达到7312.14t,COD 为57495.5t,这是造成胶州湾营养盐增加的一个重要原因。胶州湾是几乎被青岛市区所包围的半封闭的小海湾,近几十年来,更由于包括人类活动(如围海造陆)在内的种种原因而造成水域面积逐渐变小,由1935年的559km 2减少到1988年的374.4km 2,缩小了近三分之一,纳潮量亦随之显著减小,由原先的12.667@108m 3缩小为9.48@108m 3
(赵淑江等,2001a),因而大大降低了胶州湾水域环境容量与自净能力,加速了水域富营养化进程。2.3.2 海水养殖 胶州湾海水养殖业十分发达,海水养殖面积从1978年的1330ha 发展至2001年的13737ha(依据城阳区和黄岛区海水养殖面积统计),增长了约10倍,近10年来发展尤为迅速(图10)。胶州湾海水养殖方式主要有四种:一是浅海筏式养殖,主要养殖栉孔扇贝和贻贝等;二是沿岸池塘养殖,主要养殖对虾;三是滩涂底播护滩养殖,主要养殖蛤仔等;四是网箱养鱼,主要养殖鲈鱼、牙鲆和黑
等。
图10 胶州湾海水养殖面积长期变化
Fig.10 Long -term changes of mariculture area in Jiaozhou Bay
海水养殖的自身污染问题相当突出,被养殖动物摄食而未消化的饵料作为粪便排出,而被消化和吸收的部分中有些则通过代谢作为氨和尿素排入水体。以筏式贝类养殖为例,据李元山等
(1996)报道,栉孔扇贝每个体每天排粪量为179mg (干重),以通常每公顷养殖扇贝1.5@106个计算,则每公顷每年向水体排放的粪便量达到97.95t 。胶州湾四万亩筏式贝类养殖每年向水域排入粪量高达2.6万t,如此巨量的生物性沉积为微生物包括某些病源生物大量孳生提供有利条件,给胶州湾生态环境带来不良影响。养殖生物排泄物经微生物代谢矿化后以营养盐形式进入水体,导致水域营养盐含量增加和富营养化。利用1981年及1992)2002年的调查资料进行数理统计得出,胶州湾DI N 、NH 4-N 和NO 3-N 及SiO 3-Si 同海水养殖面积变化间的相关系数分别为0.81、0.78、0.71和0.67,显示出海水养殖同水域营养
盐含量之间的重要关系。所以海水养殖是除上述城市排污外的又一个造成水域营养盐迅速增加的源泉,当然亦是导致浮游植物数量增加和赤潮频
频发生的一个重要原因。实际上,近10多年来,我国北方沿海赤潮的发生多与海水养殖有关(吴玉霖等,2001)。
滤食性贝类具有另一个生态功能,即通过其强劲的滤食活动对浮游植物数量的控制作用,栉孔扇贝的滤水率可高达5.24L/(ind #h)(方建光等,1996)。80年代初庙岛海峡年平均浮游植物数量为3.0@106cells/m 3,由于发展扇贝养殖,至80年代末,数量下降至0.54@106
cells/m 3
,约为原先的六分之一(项福亭等,1996)。有大规模扇贝养殖的烟台四十里湾海域,在扇贝迅速生长的11月份,浮游植物密度仅为5.23@104cells/m 3(吴玉霖等,2001),这是对在2. 2.1节中揭示的1981)1997年伴随筏式贝类养殖业的迅速发
6期吴玉霖等:环境长期变化对胶州湾浮游植物群落结构的影响495
展,胶州湾浮游植物数量呈下降趋势现象的又一个明证。随着1997年后因病害等原因扇贝大量死亡,出现养殖业大滑坡,仅黄岛区由原先12000亩养殖扇贝下降至2000年仅剩8000亩(刘元刚等,2001)1)。此后出现的现象是自1998年开始胶州湾浮游植物数量迅速增加,并常暴发赤潮,显示出滤食性贝类既有污染水环境,促进
水域富营养化的作用,同时又对浮游植物数量动态变化有强大的生态调控机能。
2.3.3 水温 自1900年至今100年来,胶州湾地区四季平均气温呈上升趋势,其中以冬季气温上升幅度较大,上世纪90年代冬季平均气温比20世纪初上升2.64e ,秋季次之,相对说,夏季的升幅最小,为0.56e (表3)。
1)刘元刚,谢佃臣,2001.黄岛区海珍品养殖现状及发展对策.第三届全国海珍品养殖研讨会论文集,137)141
表3 胶州湾地区20世纪初和20世纪末四季平均气温(e )及其变化1)
Tab.3 Average air temperatures(e )and their changes in Jiaozhou Bay area i n the early and late 20th century 季节冬季春季夏季秋季20世纪初-0.0115.4824.938.3990年代 2.6316.4924.499.81上升幅度
2.64
1.01
0.56
1.42
1)引自赵淑江等,2001a
受气温变化的影响,胶州湾水温亦存在一定的变化。以1981、1992)2000年和2001)2003年3个时间段四个季节月平均水温进行比较(表4),可以看出,冬、春季水温均有上升趋势,20年来,上升幅度约2e ,而夏、秋季变化小,无明显趋势。由于各种浮游植物对水温适应能力不同,所以水温的变化会引起浮游植物群落结构的改变,正如3.2.2节中所述的,胶州湾原先若干优势种如窄隙角毛藻、菱形海线藻、刚毛根管藻等至90年代数量明显下降,不再是优势种,而一些广温性种类如尖刺拟菱形藻、奇异菱形藻和中肋骨条藻近年来数量明显上升,特别是一些暖水性种类,如波状石丝藻,1981年6月就开始出现在胶州湾南部和
中部;9月遍布全湾,以湾口数量最多,亦不过15@103
cells/m 3
;10月份数量急剧下降,仅为9月
份的1/3。但近年来,该藻自5月份起就在湾内出现,且数量明显增多,2003年6月上旬,该藻已遍布全湾,平均数量高达298@103
cells/m 3
,比1981年的数量增加约20倍。洛氏角毛藻(Ch.loren -zianus)为热带和温带沿岸性种类,经常出现在暖海中,近些年来,该藻在胶州湾的数量有明显增加,如2003年9月全湾平均数量高达9046@103cells/m 3,占该月浮游植物总量的42.9%,至11月仍为胶州湾主要优势种之一。由此可见,水温的变化对胶州湾浮游植物各种群数量及分布状况均有一定影响,并导致浮游植物群落结构的变化。
表4 胶州湾水温(e )的长期变化
Tab.4 Long -term changes of the average water temperatures(e )in Jiaozhou Bay
年份冬季春季夏季秋季1981 2.6113.5625.7812.501992)2000 3.9915.9426.1814.162001)2003
4.40
16.27
25.65
12.71
496 海 洋 与 湖 沼36卷
3小结
3.1大规模的人类活动给胶州湾环境带来很大
影响
由于越来越多的生活污水和工农业生产(包括海水养殖业)的废水源源不断进入湾内,造成近40年来水域营养盐含量迅速增加,近3年来NO3-N和NH4-N比1962年分别提高14.5倍和9.5倍,加上沿岸带开发,胶州湾水域面积不断缩小,纳潮量下降,水域环境容量和自净能力减弱,导致胶州湾环境正向富营养化方向蜕变。
3.2胶州湾水环境的长期变化对浮游植物群落
结构产生明显的影响
3.2.1自1954年以来,浮游植物数量总体上看呈增加的趋势,特别是在1997年开始大规模滤食性贝类养殖业因病害等原因而出现大滑坡至今,浮游植物数量有明显增长的势头。
3.2.2近50年来,胶州湾浮游植物优势种更替明显,一些原先优势种数量下降,不再居优势地位,而比较喜肥的种类如中肋骨条藻数量显著增加,成为最重要的优势种;一些暖水性种类如波状石鼓藻提前在湾内出现,且数量上有很大增加。
3.2.3胶州湾浮游植物多样性指数近50年来有所下降,反映出种类数量与各物种种群丰度间的变化,表明胶州湾浮游植物群落结构处在演化之中。
3.2.4由于营养盐含量迅速增加,为赤潮的形成与发展提供丰富的物质基础。自1997年以来赤潮几乎连年暴发,有愈演愈烈的发展趋势,反映出胶州湾生态环境已出现异常的征兆。
参考文献
方建光,匡世焕,孙慧玲等,1996.桑沟湾栉孔扇贝养殖容量的研究.海洋水产研究,17(2):18)31[Fang J G, Kuang S H,Sun H L et al,1996.Study on the carrying capaci ty of Sanggou Bay for the culture of scallop Chla mys farre ri.M arine Fisheries Research,17(2):18)31]
孙松,刘桂梅,张永山等,2002.90年代胶州湾浮游植物种类组成和数量分布特征.海洋与湖沼(浮游动物研究专辑),37)44[Sun S,Liu G M,Zhang Y S et al, https://www.doczj.com/doc/c93053498.html,munity composition and distribution character of phytoplankton in the Jiaozhou Bay in the1990s.
Oceanologia et Limnologia Sinica(Special Issue for China GLOBEC Zooplankton Population Dynamics Research),
37)44]
吴玉霖,孙松,张永山等,2004.胶州湾浮游植物数量长期动态变化的研究.海洋与湖沼,35(6):518)523
[Wu Y L,Sun S,Zhang Y S et al,2004.Quantitative study on long-term variation of phytoplankton in Jiaozhou Bay.Oceanologia et Li mnologia Sinica,35(6):518)523]吴玉霖,周成旭,张永山等,2001.烟台四十里湾海域红色裸甲藻赤潮发展过程及其成因.海洋与湖沼,32(2): 159)167[Wu Y L,Zhou C X,Zhang Y S et al,2001.
Evolution and causes of formation of G ymnodin ium san-guineum bloom in Yantai Sishili Bay.Oceanologia et Li m-nologia Sinica,32(2):159)167]
李元山,牟绍敦,冯月群等,1996.海珍品综合增养殖中的种间关系和生态容纳量研究.海洋湖沼通报,(1):
24)29[Li Y S,Mu S D,Feng Y Q et al,1996.Study on
the interspecific relationship and ecological capacity in rare marine animals culture.Transactions of Oceanology and L i mnology,(1):24)29]
李冠国,黄世玫,1956.青岛近海浮游矽藻季节变化研究的初步报告.山东大学学报,2(4):119)143[L i G G, Huang S M,1956.Seasonal variation of plankton diatoms at Tsingtao.Acta Scientiarum Naturalium,2(4):119)
143]
沈志良,2002.胶州湾营养盐结构的长期变化及其对生态环境的影响.海洋与湖沼,33(3):322)331[Shen Z L, 2002.Long-term changes in nutrient structure and i ts influences on ecology and environment in Jiaozhou Bay.
Oceanologia et Li mnologia Sinica,33(3):322)331]
沈志良,陆家平,刘兴俊等,1994.胶州湾水域的营养盐.
海洋科学集刊,35:115)129[Shen Z L,Lu J P,Liu X J et al,1994.Nutrien ts in Jiaozhou Bay Waters.Studia Ma-rina Sinica,35:115)129]
周成旭,吴玉霖,2000.赤潮发生机理.苏纪兰,秦蕴珊主编.当代海洋科学学科前沿.北京:学苑出版社, 359)362
赵淑江,沈志良,张铭汉等,2001a.胶州湾理化因子的长期变化.焦念志,赵卫红,赵淑江等著.海湾生态过程与持续发展.北京:科学出版社,207)240
赵淑江,吴玉霖,王克等,2001b.胶州湾生物因子的长期变化.焦念志,赵卫红,赵淑江等著.海湾生态过程与持续发展.北京:科学出版社,241)256
项福亭,曲维功,张益额等,1996.庙岛海峡以东浅海养殖结构调整的研究.齐鲁渔业,13(2):1)4[Xiang F T, Qu W G,Zhan g Y E et al,1996.On aquaculture structure adjustment in Shallow Sea Eas t to Miaodao Strait.Shandong Fi sheries,13(2):1)4]
郭玉洁,杨则禹,1992.浮游植物.见:刘瑞玉主编.胶州湾生态学和生物资源.北京:科学出版社,136)169 Andren E,1999.Changes in the composition of the diatom flora during the last century indicate increased entrophication of the Oder Estuary,south-western Baltic Sea.Estuarine,
6期吴玉霖等:环境长期变化对胶州湾浮游植物群落结构的影响497
Coastal and Shelf Science,40:339)351
Copper S R,Bruch G S,1993.A2500-year history of anoxia and eutrophication Chesapeake Bay.Estuarines,16:617)626 Jus tic D,Rabalais N N,Turner R E et al,1995.Changes i n nu trient structure of river-dominated coastal waters:stoichio-metric nutrient balance and its consequences.Estuarine, Coastal and Shelf Science,40:339)356
Patsch J,Radach G,1997.Long-term simulation of the eatroph-ication of the North Sea temporal development of nutrients,
chlorophyll and pri mary production i n comparison to obser-vations.Journal of Sea Research,38:275)310Smayda T J,1984.Variations and long-term changes in Narra-gansett Bay,a phytoplankton-based coastal marine eacosys-tem;relevance to field monitoring for pollution assessment.
In:Whife H H ed.Concepts in Marine Pollation M easure-ments.Maryland Sea Grant College University of Maryland Press,663)679
Zhao S J,Jiao N Z,Shen Z L et al,2005.Causes and conse-q uences of nutrient structure in the Jiaozhou Bay.Journal of Integrative Plant Biology(Formerly Acta Bot Si n),47(4): 396)410
LONG-TER M C HANGE OF ENVIRONMENT A ND IT.S INFLUENCE ON
PHY TOPLANKTON COMMUNITY STRUCTURE IN JIAOZHOU BAY
WU Yu-Lin,SUN Song,Z HANG Yong-Shan
(Key Laboratory o f Marine Ecology and En vironmental Sciences,
Institute o f Oceanology,Chinese Academy o f Sc iences,Qingdao,266071)
Abstract Long-term variation of environmental factors and their influences on phytoplankton were revealed by data obtained in51quarterly cruises(February,May,August and Nove mber)during1991)2004in Jiaozhou Bay combined with the historical data over the past50years.Phytoplankton were sampled with nets(37cm diameter, 140cm length,77L m mesh)from bottom to surface for subsequent counting of cell number under microscope.
Jiaozhou Bay is a sem-i enclosed bay situated in the western part of Shandong peninsula,C hina,which is sur-rounded by Qingdao City,with an area of about390km2and an average water depth of about7m.The bay mouth is narrow,only about2.5km wide.The distribution of phytoplankton that characterized by dense area appeared main-ly in northeastern and northwestern parts near-shore area,and in the middle and southern parts,the density was low.The phytoplankton abundance was higher inside the bay than outside the bay.Clear seasonal variation of phyto-plankton was shown.The peak in phytoplankton abundance occurred frequently in winter and summer;and the trough value occurred mainly in autumn.Human ac tivities have greatly impacted the environment of Jiaozhou Bay. Large amount of industrial and residential wastewater concentrated with N,P and organic matters discharged into the bay every https://www.doczj.com/doc/c93053498.html,rge-scale maricultures of filter-feeding bivalve,fish and so on have caused serious ecological prob-lems in the bay.Nutrient concentrations had been continuously rising since1960s.NO3-N,NH4-N and PO4-P were doubled by7.26,5.10and2.43times from1960s to1990s,respectively.The mean contents of these nutrients in 2001)2003were obviously higher than that in1990s.Significant long-term environmental changes have affected the succession of phytoplankton community constructure.The phytoplankton species composition,abundance and dom-i nant species have also experienced obvious variation since1954.Dominant species have considerably alternated. Some species such as Chaetoceros affinis,Thalassionema nitzschioides,Rhizosolenia setigera,Coscinodiscus radia-cus have been replaced by Asterionella kariana,Thalassiosira no rdenskioldi,Nitzschia parado xa and so on.Some eutrophic species such as Skeletonema costatum and a warm water species Lithodesmium undulatum increased abun-dantly.The phytoplankton diversity inde x(Shannon.s H c)has slightly decreased during past50years.From1977 to present,red tide events have been increasing in number,frequency and intensity in Jiaozhou Bay.Marine eutrophication is widely believed to be the major cause of red tide.
Key words Jiaozhou Bay,Environmental change,Phytoplankton,Community structure 498海洋与湖沼36卷
杭州疏林草地优秀案例之一花港观鱼雪松大草坪案例分析 “乱花渐欲迷人眼,浅草才能没马蹄”,这是古代诗人对杭州西湖景色的描述,道出了杭州草坪是那么欣欣然而有生意。在杭州的园林绿地里,草坪占有相当大的比重,如花港观鱼、柳浪闻莺两个公园中,开阔的草坪约占整个公园陆地面积的40%左右[4]。但杭州夏季炎热,不宜多采用大面积的空旷草坪,以疏林草地为主,可提高草坪在夏季的利用率。疏林草地是杭州园林植物造景的主要特色之一,涌现出许多疏林草地的植物造景佳作。本专题将对杭州优秀的疏林草地案例进行分析。 花港观鱼雪松大草坪平面图 花港观鱼雪松大草坪 花港观鱼雪松大草坪面积约14080m2,是花港观鱼公园内最大的草坪活动空间,也是杭州疏林草地景观的杰出代表。 雪松大草坪以高大挺拔的雪松作为主要的植物材料,在体量上相互衬托,十分匹配。雪松单一树种的集中种植体现树种的群体美;适当的缓坡地形,更强调了雪松伟岸的树形。四角种植的方式,既明确限定了空间,又留出了中央充分的观景空间和活动空间,景观效果与功能都得到了极大的满足。 根据植物景观的平面布局,雪松大草坪可基本划分为3组植物,下面分别分析各组植物的景观特色。 A组植物(图02)
为强调公园的休闲性质、适当缓和雪松围合形成的肃穆气氛,设计者在本组雪松林缘错落种植了8棵樱花,春季景观效果突出。 该组植物结构简单、层次分明。 雪松深绿色的背景为盛开的樱花提供了极佳的背景,折线状自然种植的单排樱花恰似一片浮云,蔚为壮观。 其合理的间距与冠幅体现了整体性与连续性。由于樱花的观赏时间较短,故大多数时间仍以欣赏雪松群植的形体美为主。 樱花的平均高度约为雪松平均高度的1/3,上下层次清晰。樱花间距5~8m,为现有平均冠幅的1倍以上,三三两两的组合彼此呼应,体现了视觉上的连续性,并预留了较大的生长空间。 在平面图上还可以发现,8株樱花的疏密变化与12株雪松的组合颇为类似,中间紧,两头松,模拟自然界从密林至林缘的生长模式产生自然的景观效果,并以类似的组合方式使两种植物具有内在的联系,和谐统一。 B组植物(图03-04)
第八章植物群落结构 第一节:植物群落及其种类组成 一、植物群落的概念与基本特征: (一)概念 1. 生物群落 在特定的时间、空间或生境下,具有一定的生物种类组成,外貌结构,各种生物之间,生物与环境之间彼此影响,相互作用,并具特定功能的生物集合体。 2. 植物群落 特定空间或特定生境下,植物种群有规律的组合。 即一定地段上,群居在一起的各种植物种群所构成的一种有规律的集合体。 群落生态学(synecology)是研究群落与环境相互关系的科学。 二、植物群落的基本特征 ?具有一定的物种组成物种数和个体数。 ?不同物种之间的相互影响:必须共同适应它们所处的无机环境;它们内部的相互关系必须 取得协调和发展(种群构成群落的二个条件)。 ?具有形成群落环境的功能:定居生物对生活环境的改造结果。 ?具有一定的外貌和结构:形态结构、生态结构、营养结构。 ?具有一定的动态特征 :季节动态、年际动态、演替与演化。 ?具有一定的分布范围:特定的地段或特定的生境。 ?具有边界特征:或明确或不明确的边界。 群落的物种组成 ?实验原理: ?1. 取样方法——样方法 ?取样就是代表性群落的选取或确定,包括样地设置的方法、范围大小等。 ?样地大小的确定一般采用巢式样方法,通过绘制种——面积曲线来确定。 ?样地大小 不同群落类型最小面积经验值 样地形状 a.传统形状——方形 故称样方(quadrat)
b.圆形 也称样圆 c.矩形 也称样带(belt)或样条(transect) (二)种类组成的性质分析 根据各个种在群落中的作用不同,将其划分为几个不同的群落成员型。植物群落研究中,常用的群落成员型有以下几类: 1.优势种和建群种 (1)优势种:在群落中能有效控制能量流动和物质循环并对群落的结构和群落环境的形成具有明显的控制作用的生物种。 特征:个体数量多,投影盖度大,生物量高,体积大,生活能力强。(2)建群种:吧优势层的优势种称为建群种,其中决定着群落的外貌,而且一控制着群落的生态环境和群落的其他组成成员。 如果群落中的建群种只有一个,则称为“单建群种群落”或“单优种群落”。如果具有两个或两个以上同等重要的建群种,则称为“共建种群落”或“共优种群落”。 2.亚优势种(subdominant species) 指个体数量与作用都次与优势种,但在决定群落性质和控制群落环境方面仍起着一定作用的植物种。 3.伴生种(companion species) 伴生种为群落的常见种类,它与优势种相伴存在,但不起主要作用。 4.偶见种或罕见种(rare species) 是那些在群落中出现频率很低的种类,往往是由于种群自身数量稀少的缘故。偶见种可能是偶然的机会由人带入、或伴随着某种条件改变而侵入,也可能是衰退中的残遗种。 (三)种类组成的数量特征 ?个体数量指标 ?综合数量指标 个体数量指标 1 多度(abundance)与密度(density) 多度是对植物群落中物种个体数目多少的一种估测指标。 德鲁提(Drude)的七级制多度。即:查。国内多采用Drude七级制多度,即: Soc 极多,植物地上部分郁闭,形成背景 Cop3 数量很多 Cop2 数量多 Cop1 数量尚多 Sp 数量不多而分散 Sol 数量很少而稀疏 UN 个别或单株
[案例分析]生物教学:环境污染对生物的影响1 教学活动对象:高一学生 教学活动准备:开放生物实验室,并准备学生活动所需的各类仪器装置;实验所需各种生物、各类污染物等主要由学生自己采集、准备。 教学活动过程:该主题的教学活动过程主要分为以下步骤: (1) 教师提出课题“环境污染对生物的影响”。 (2) 学生调查学校周围环境中的主要污染现象,分析污染原因。如让学生走访区环保局和环境监测站,随同专业人员采集黄浦江水样、测定水样,调查学校周围环境的空气、水质和绿化现状等。 (3) 学生经过对周边环境的各类污染因素与常见生物的关系的调查和分析后,组成若干课题研究小组(每组3-5人),各自选定实验研究项目。 (4) 各小组相互评议实验研究项目,进行可行性论证,然后确定实验研究项目。 (5) 各小组设计具体的实验研究方案。实验方案中应包括以下内容:①研究题目;②研究目的;③实验原理;④所需材料(应具有可行性);⑤具体实验步骤;⑥预期结果。 (6) 师生分别作实验准备。 (7) 在课堂内,各组学生按照自己的实验方案进行操作。小组成员之间应相互协作,相互切磋,共同解决实验中出现的问题。 (8) 各组间相互交流实验研究的过程和结果,相互进行评议和质询,提出自己的不同看法。 各组在听取评议的基础上进一步完善实验或提出进一步研究的方案。 (9) 学生写出实验研究报告,提出自己对实验研究结果的见解。 在“环境污染对生物的影响”教学案例中,学生的探究活动分为形成概念和问题、制定学习计划、开展探究活动、总结发现四个阶段。在第一阶段,教师就“环境污染对生物产生的影响”这一现象要求学生进行多种体验,通过调查活动学生形成一系列概念和问题,从而引发学生探究的兴趣。第二阶段开始划分学习小组并进行小组讨论,以选定各自的实验研究项目,制定实验研究计划。第三阶段主要依靠学生自己开展探究活动,教师给予学生适度的辅导。探究的最后阶段是以实验报告的形式来进行总结活动,教师明确提出了实验报告的格式和要求等,并预先制定了相应的量规用于评价学生的整个学习和探究过程。 1.研究课题:环境污染对生物的影响。 2.活动目标: 在活动中提高学生的环保意识和科研意识; 在实验研究的过程中促进学生发展创造性思维; 培养学生设计和操作实验的能力; 培养学生相互合作的精神。 3.参加活动对象:高-年级部分学生(由学生自由报名)。 4,活动的准备: 开放生物实验室,并准备学生活动所需的各类仪器装置。实验所需各种生物、各类污染物等主要由学生自己采集、准备。 5.活动过程: (1)教师就课题"环境污染对生物的影响"概述进行科学实验与研究的基本方法。 (2)学生调查学校周围环境中的主要污染现象,分析污染原因。如让学生走访区环保局和环境监测站,随同1案例来源:上海故业中学费循蛟老师https://www.doczj.com/doc/c93053498.html,/3_anli/3_jijin/jijin_008.htm
城市园林设计植物群落的构建 (一)人工群落 在生态系统中,由于受遗传、受粉及物种间相互作用的生物机制的影响,物种的多样性只有在群落的水平上才能得到很好地维持和保护。研究表明,由乔灌草组成的立体式群落结构具有较好的生态效益,能有效提高生态系统稳定性,同时也为昆虫、鸟类等野生动物的引入提供良好的条件。然而当前多数绿地系统所谓的乔、灌、草复层混交林模式往往只是三者的简单组合,物种单一,群落结构单调,难以实现群落正常的生态过程。人工植物群落构建应参考当地自然植被类型、生活型谱分析,确定城市树种规划中乔、灌、草的比例;依据群落树种的重要值高低,确定拟建人工群落中建群种、优势种和伴生种,使拟建人工群落中各树种的数量比例更加合理,在此基础上,依据当地自然植被中群落类型的多寡,提出城市绿地系统可以构建的人工群落类型数量。 (二)自然群落 根据人为活动对植被影响的程度,把城市植被分为人工栽培群落、残存自然群落和城市杂草群落3个类型人工栽培群落是指人为引入城市区域的群落类型,主要有市区的花坛、绿地以及郊区的农田和人工防护林等。残存自然群落指人为活动影响之前就已经存在,在城市化过程中未被清除的原生或次生的自然群落,如寺庙周围的风水林。城市杂草群落是城市化后,不受人的意识支配而出现的植物群落,主要
包括归化植物和当地的乡土植物。其中,后两者在城市中所占比例已经不多,但是在生物多样性的维持方面却能发挥很大作用。过去,城市自然遗留地和自然植被有时被看作是视觉差、景观低劣的荒芜地,但它们常具有地带性群落特征,含有丰富的乡土植物,与野生动物也有较好的生态关系,生物多样性潜力很大。 在城市生物多样性规划中,应更多考虑如何恢复和重建城市自然生态景观,在确保景观美学需要的前提下,使它们更好地发挥生态功能,丰富城市景观,增加城市的生物多样性。自然绿化在我国尚属于探索阶段,但是在发达国家,"近自然"群落建设的理念和技术已被广泛运用和推广,以草坪为例,人工草坪不仅维护费用高,而且农药化肥的投入会给环境造成一定程度的污染。即便是在拥有几十年甚至上百年草坪建设经验的欧美发达国家,最终放弃了人工草坪建设,重新选择低养护成本的野草作为自然绿化。既保护了环境,又能吸引昆虫、鸟类和其他小动物来栖息,能很好地增加城市的生物多样性,同时方便市民在草地上休闲娱乐,真正体现"以人为本"的设计宗旨。
第三节城市绿地植物群落结构特征 一、植物群落结构相关概念 1、自然植物群落与人工植物群落 植物群落指在某一地段内全部植物在时空分布上的综合,在一定的生境条件下,具有相对的种类组成与数量比例和特定的结构与外貌,发挥着一定的功能。植物群落按照其形成可分为自然植物群落和人工植物群落。自然植物群落指植物在长期的历史发育过程中自然形成的群落。人工植物群落是指通过人为干涉,按照人们意愿与功能需求,在模拟与借鉴自然植物群落结构的基础上,通过对植物进行选择、配置、营造与管理而形成的植物群落,城市绿地植物群落是人工植物群落的重要组成部分。著名学者王伯荪将城市植被定义为城市里覆盖的生活植物,是完全不同于自然植被的特点、性质以及生境的植物群落。通过概念的解析,城市绿地植物群落与自然植物群落两者之间的异同在于:首先,植物群落所处生境不同;“城市”为人工植物群落限定了范畴,是以城市环境为背景,伴随着城市发展而形成的具有人工化典型特征的群落。所涉及的城市环境包含有城市的建筑,小气候、地形、土壤等综合因素,这些都有别于自然植物群落所具有的特征。其次,植物群落生长发育过程存在一定差异;城市绿地植物群落多是出自多元化的功能与服务等方面考虑,按照人们意愿进行的植物配置,人为选择决定了植物群落的空间结构特征。 2、植物种植设计与植物群落结构 植物种植设计,与之含义相近的概念有植物配置、植物造景、植物景观设计。从以下几个方面来理解植物种植设计:从美学角度出发,植物种植设计是
利用不同种类的植物来营造景观空间,使其充分发挥植物群落或个体的形态、色彩、线条等方面的自然美。从生态学角度出发,植物种植设计是依据立地条件与植物自身的生长规律而营造的植物群落,使其满足植物最佳生长状态所需的环境资源,从而很好地发挥生态效益与服务功能以达到改善生态环境的目的。从行为心理学角度出发,植物种植设计是通过植物的巧妙组合来满足城市中的人们不同的生理与心理方面的需求。由此可见,植物种植设计是满足不同功能需求的不同植物种类在空间上的布置。植物种植设计是城市绿地植物群落结构形成的前提,绿地类型、功能以及种植设计形式的不同,都将会引起城市绿地植物群落结构的异同。群落结构指某个等级或尺度(种群、群落)的生态系统中不同性状与大小的组成单元在空间上的分布与排列。植物群落结构体现了群落内个体间的空间分布格局,主要包括以下几个方面:第一,群落中个体在数量上的聚集程度,如密度、多度、尺度等;第二,群落中个体在空间上的组合关系,如水平分布与垂直结构等;第三,群落中个体在形态上的分化程度,如胸径、冠幅、树高等。此外,植物群落历经长时间的进化与演替,形成了相对稳定具有一定逻辑关系的结构与形态。因此,植物群落结构是不同生态学过程在不同时间与空间尺度上综合作用的结果。 二、植物群落各组件结构特征与属性 1、冠层结构特征 植物的生长发育与树冠有着密切联系,树冠不仅对太阳能等资源的分配起到关键作用,还对林下植物群落的组成及生长变化产生直接或间接的影响。作为光合作用的主要场所,树冠的形态、尺度以及健康状况与否对植物活力及生产力起到关键作用。植物群落中不同植物种类所组成的冠层结构在植物群落结
植物群落调查 考察植物群落有各种方法,如样地法、样线法、距离抽样法、点样法等。 其中样地法是基础方法,用样地法进行调查的方法步骤说明如下: (一)样地的设置 样地不是群落的全部面积,它仅是代表群落的基本持征的一定地段。对植物群落考察应在确定的样地内进行,通过详细调查,以此来估计推断整个群落的情况。 样地选择的方法:选择样地应遵循下列原则:(1)种的分布要有均匀性。(2)结构完整,层次分明。(3)环境条件(尤指土壤和地形)一致。(4)群落的中心部位,避免过渡地段。 1.样地的形状:大多采用方形,又称样方;除此还有样条,样线,弱圆等。可根据不同研究内容具体选择。 小型样方用于调查草本群落或林下草本植物层,大型样方用于调查森林群落或荒漠中的群落。为防止出现闭合差,在森林调查中,样方常沿着预定的测线方向呈菱形设置。其方法是由中心点定出距离为样方对角线长度的两个点,然后从这两点分别拉直长度恰为样方边长的测绳,使其在每一侧都恰好交接,就是样方的边界。 2.样地面积 下列样地面积的经验值可供考察时参考使用:草本群落1~10m2,灌丛16~100m2,单纯针叶林100m2,复层针叶林、夏绿阔叶林400~500m2,亚热带常绿阔叶林1000m2,热带雨林2500m2 3.样地数目 样地数目多少取决于群落结构复杂程度。根据统计检验理论,多于30个样地的数值,才比较可靠。为了节省人力与时间,考察时每类群落根据实际情况可选择3~5个样地;所有样地应依照顺序进行编号,以免混乱。 4.样地布局:一般可选用主观取样法,即选择被认为有代表性的地块作为调查样地。(二)植物群落样地调查内容与方法 样地调查内容主要有环境条件,群落的空间结构,群落的组成特征,群落的外貌。 1.环境条件调查:包括以下五项:(1)地理位置,(2)地形条件。(3)土壤条件。
二氧化硫对植物的影响 张涛 20135937 摘要:近年来SO2污染比较严重,它对植物有着多方面的影响。植物既受到SO2污染的影响,又对SO2的影响具有一定程度的修复能力。本文总结了关于SO2单一污染物对植物生理生化的直接影响以及其适应机制,并提出对这方面研究的展望。 关键词:二氧化硫;植物;抗氧化酶 我国是以煤为主要能源的国家,所以我国的大气污染主要是以SO 2 污染为主。特别是近30年来我国经济的高速发展,更使煤炭以及石油的消耗量达到 了一个前所未有的高度,加剧了SO 2的排放污染。SO 2 是我国当前最主要 的大气污染物,在个别地区污染相当严重。SO 2 可通过气孔进入植物叶片细 胞后快速溶于细胞中,在细胞内释放出H+、HSO 3-和SO 3 2-等,从而对细 胞产生直接或间接的伤害。也可与其它大气污染物进行化学反应,生成各种硫酸盐,这些成分随雨水共同降落成为“酸雨”,能够导致土壤和水系的酸化,干扰植物的代谢,对生态系统有很大的破坏作用,从而间接地危害人类健康。关 于SO 2 污染环境对植物生理生化及生长发育的影响已引起了众多学者的关 注,并己取得了长足的进展。近年来,在SO 2 的植物伤害症状、伤害机理、对生理生化指标、植物组织结构影响等方面取的研究得了许多进展。 1.二氧化硫对植物形态的影响 李利红,仪慧兰[1]等采用室内培养及密闭箱静态熏气方法,研究了不同浓 度SO 2暴露对拟南芥叶片形态的影响,结果显示:SO 2 暴露对拟南芥成熟 叶片的伤害主要是叶面伤害斑的出现和叶片枯死,伤
害程度与暴露浓度和时间呈正相关,暴露于低浓度SO 2 时叶面无伤害斑,随 时间推移有少数叶片边缘卷曲,但在停止暴露后恢复正常;中浓度时暴露的植株叶片出现大小不等的透明斑,随着暴露时间的延长,伤害症状发展为坏死斑, 暴露于高浓度SO 2 的植株,叶片很快出现不规则形的黄色坏死斑,坏 死斑的面积随暴露时间的延长而扩大,之后叶片大量枯死。但在脱离高浓度S O 2 后伤害性斑点不再增加,并能继续生长发育。 SO 2暴露对拟南芥植株的生长发育具有双向作用,较低浓度SO 2 暴露 对植株的生长发育有一定的促进作用,高浓度SO 2 暴露会抑制植株的生长发育,使株高、单株叶片数和单叶面积呈浓度依赖性减少。 2二氧化硫对植物生理生化的影响 2.1二氧化硫对植物气孔的影响 气孔是植物与外界环境间气体交换的主要通道,气体污染物主要通过气孔进入叶组织,因此气孔在大气污染物对植物的影响中占有相当重要的地位。高吉喜 [2]通过试验表明:通常情况下 SO2 促使植物气孔关闭,但也有某些植物经S O 2熏气后气孔关闭。气孔对SO 2 浓度的反应通常是SO 2 浓度越大,气孔 反应越快。 2.2二氧化硫对植物细胞膜的影响 细胞膜是植物细胞的重要组成部分,起着调节控制细胞内外物质交流的屏障作用,当植物处在不利环境条件下时,刺激首先作用于细胞膜。大量观察研 究表明,细胞膜也是SO 2作用的最初部位,在植物接触高浓度SO 2 后,膜 首先受到损伤,继而膜透性发生改变。植物膜透性对SO 2 的反应差异通常与 植物的抗性有关,抗SO 2强的植物,细胞膜对SO 2 的反应不敏感,反之则很
抗生素对土壤环境的污染与植物修复的研究与展望 作者:黄盼盼, 周启星, 董璐玺 作者单位:南开大学环境科学与工程学院,中国,天津,300071 刊名: 科技信息 英文刊名:SCIENCE & TECHNOLOGY INFORMATION 年,卷(期):2010(11) 参考文献(19条) 1.INGERSIEVH TORANGL;LOCKM L;et a1Primary blodegradation of veterinary antibiotics in aerobic and anaerobic surface water simulation systems 2001 2.Diaz2Cruz M S;Larcelo D Environmental behavior and analysis of veterinary and human drugs in soils,sediments and sludge 2003(06) 3.王丽平;章明奎;郑顺安土壤中恩诺沙星的吸附-解吸特性和生物学效应[期刊论文]-土壤通报 2008(02) 4.Angel J.Baguer;John Jensen;Paul Henning Krogh Effects of the antibiotics ox tetracyeline and tyrosine on soil fauna 2000 5.李兆君;姚志鹏;张杰;梁永超兽用抗生素在土壤环境中的行为及其生态毒理效应研究进展[期刊论文]-生态毒理学报 2008(01) 6.Cerd Hamscher;Sake Sczesny;Heinrich Hoper;Heinz Nau Determination of Persistent Tetracycline Residues in Soll Fertilired with Liquid Manure by High-Performance Liquid Chromatography with Electroe prays Ionization Tandem Mass Spectrometry 2002 7.S.Thiele-Bruhn;M-0,Aust effects of pig slurry on the sorption of Sulfonamide Antibiotics in soil 2004 8.曹卫东;王旭;刘传平;封朝晖,刘红芳,李芳柏当前部分有机肥料中的持久性有机污染问题[期刊论文]-土壤肥料2006(02) 9.王丽平;章明奎土霉素污染对土壤生物学性质影响的初步研究[期刊论文]-农业环境科学学报 2009(07) 10.刁晓平;孙英健;孙振钧;沈建忠安普霉对对不同土壤中微生物活动的影响[期刊论文]-生态环境 2004(04) 11.孔维栋;朱永官抗生素类兽药对植物和土壤微生物的生态毒理学效应研究进展[期刊论文]-生态毒理学报2007(01) 12.周启星;宋玉芳植物修复的技术内涵及展望[期刊论文]-安全与环境学报 2001(03) 13.安凤春;莫汉宏;郑明辉;张兵DDT及其主要降解产物污染土壤的植物修复[期刊论文]-环境化学 2003(01) 14.王校常;施卫明;曹志洪重金属的植物修复-绿色清洁的污染治理技术[期刊论文]-核农学摄 2000(05) 15.牛建平;吴泽辉;石起增磺胺二甲嘧啶在土壤中的降解动态研究[期刊论文]-安徽农业科学 2009(04) 16.章明奎;王丽平;郑顺安两种外源抗生素在农业土壤中的吸附与迁移特性[期刊论文]-生态学报 2008(02) 17.Boxall A B A;Blaekwell P;Cavallo R;et a1The sorption and transport of a sulphonamide antibiotic in soil systems 2002 18.李彦文;莫测辉;赵娜菜地土壤中磺胺类和四环素类抗生素污染特征研究[期刊论文]-环境科学 2009(06) 19.王丽平典型外源抗生素在土壤中的转归及其与土壤微生物多样性的相互作用和机理研究 2008 本文链接:https://www.doczj.com/doc/c93053498.html,/Periodical_kjxx201011297.aspx
第十章植物群落类型与分布 陆地植物群落的垂直分布 如在潮湿热带地区,随着海拔的升高逐渐更替出现热带雨林→山地雨林→常绿阔叶林→针阔混交林→寒温带暗针叶林→高山灌丛、高山草甸。 我国的植被分区 原则:是分地理地带性和植被地带性,尤其是水平地带性作为基础,并结合考虑非地带性规律。 ①寒温带针叶林区域:大兴安岭北部山地、用材基地。 ②温带针阔混交林区域:垂直分布带较明显,木材重要基地。 ③暖温带落叶阔叶林区域:地带性植被为落叶阔叶林。 ④亚热带常绿阔叶林区域:是我国面积最大的植被区,占全国总面积的1/4左右。 ⑤热带季雨林、雨林区域:是我国森林类型中植被种类最为丰富的类型。广东分布于湛江、化 州、高州和阳江一线以南。 ⑥温带草原区域:是欧亚草原区域的一部分。 ⑦温带荒漠区域:本区内有一系列巨大山系,如天山、昆仑山等。 ⑧青藏高原高寒植被区域:发育着以山地森林垂直带为代表的植被。 第二节自然植被的群落类型 一、植物群落分类 分类方法: ①按外貌分类:以优势植物的生活型(生长型)作为分类基础。 森林、矮疏林、密灌丛、草地、稀树干草原、灌木稀树干草原、树丛、稀树草地、草甸、干草原、草甸性草原、真草原、灌木干草原、草本沼泽、木本沼泽、荒原 缺点:把生态学关系差异很大的群落混淆在一起。 ②按结构分类:忽视生态特征,没注意群落与环境的统一 ③按植物区系分类:注意植物生态、生活型和地理型 ④按优势度分类:以优势树种的生态学特征作为基础 ⑤按生态分类:主要以植物群落分布的生态条件或环境状况作为分类基础. ⑥按外貌——生态分类:以群落主要层优势种的外貌(群落的结构特征)、生态特征(生活型) 和生态学指标(生态环境)为划分原则。 ⑦按演替分类:强调演替在植物群落分类上的重要性。 (二)中国的植物群落分类 以群落本身的综合特征作为分类依据 1.中国的植物群落分类系统 主要分类单位分三级:植被型(高级单位)、群系(中级单位)和群丛(基本单位)。每一等级之上和之下又各设一个辅助单位和补充单位。高级单位的分类依据侧重于外貌、结构和生态地理特征,中级和中级以下的单位则侧重于种类组成。其系统如下: 植被型组:如草地
二氧化硫污染对绿色植物的影响 上海市园林学校(200051)胡天勤 化学与生活,1996(7) 随着人类对自然资源的不断开发和工农业生产的迅速发展,大量有毒有害物质任意排放,对我们周围环境带来了严重污染。 本文就二氧化硫这一主要大气污染物对绿色植物所产生的影响作一分析和探讨。 (一)二氧化硫的来源 在大氧中有许多污染物质,如二氧化硫、NOx、臭氧、烟尘等,其中以二氧化硫为主要污染源,原因是它来源广、危害大。据统计,全球每年向大气排放的二氧化硫多达2.4亿吨左右,单在我国,就有1400万吨之多,其污染量之大令人吃惊。二氧化硫污染大气,它来自以下凡方面: (1)煤、石油等燃料的燃烧是大气中二氧化硫的主要来源。煤炭中含硫,一般含量在3%~5%左右,燃烧后即被氧化成二氧化硫,由燃料燃烧所产生的二氧化硫大多从烟囱排入大气。 (2)钢铁、炼油、有色金属冶炼、化工、水泥等工厂企业,在生产流程及工艺操作过程中,也会排放相当量的二氧化硫气体。据统计,到本世纪末。全世界二氧化硫排放总量可达3.4吨左右。而当大气中二氧化硫的含量超出0。2~0。3PPm时,一些绿色植物将会受到严重的伤害。 (二)二氧化硫对植物的危害 大气中二氧化硫污染物对植物的危害方式一般有三种: 1。急性危害:高浓度的SO2气体会大大超出植物的承受能力,使植物在短时问内(1~2天或几小时内)发生叶片枯焦脱落,生长发育严重受阻,直到枯
萎死亡。 2。慢性危害:植物因长期在低浓度SO2污染的环境中,逐渐产生不易被人们所觉察的一些症状,使植物出现不同程度的生长不良。 3。隐性危害:植物长期在低浓度SO2影响下,并未表现出任何症状,但植物内部的生理活动已受到侵害,生长发育受阻。 (三)二氧化硫危害植物的化学机理 当二氧化硫通过植物叶片上的气孔进入叶子后,被叶肉吸收,转变成亚硫酸根离子然后又可转变成硫酸根离子,由于在植物体内SO2转变成SO32-的速度要比SO32-转变成SO42-快得多,所以当高浓度的二氧化硫进入植物体内后,会造成高浓度的SO32-的积累,而SO32-对植物的毒性比SO42-扩大30倍,从这一意义上分析,二氧化硫对植物造成的损害,实际上是由于其还原作用所引起的。 (1)对气孔机能的影响 当二氧化硫气体进入叶片以SO32-形式积累起来后;便会对气孔的开启和关闭机能带来影响,使气孔机能瘫痪,从而使大量二氧化硫气体进入植物体的细胞,加重对植物的危害。此外,由于植物气孔机能受阻,还会引起水份大量蒸腾,导致植物组织迅速枯萎。 (2)对叶片组织结构的破坏 当二氧化硫通过开放的气孔进入叶片组织后,溶解在细胞中,致使细胞内含物遭破坏或变形,引起外渗与原生质分离,使叶片组织结构遭到损害,海绵细胞与栅栏细胞发生质壁分离,其主要症状为:细胞失水变形、组织破碎。栅状组织细胞的排列层次紊乱、细胞间隙增大、叶片明显变薄等。 (3)对光合作用的影响
第一节植物群落的外貌和结构 一、生活型组成特征 人们观察和区别植物群落时,首先关注的是群落中占优势的生活型,正是它赋予该群落一定的外貌形象,如森林、草原、荒漠等。 植物群落生活型的组成特征是当地各类植物与外界环境长期适应的反映。研究表明,一个大地域的典型植被,均有一定的生活型谱(表4-1 ),而且一定的植被类型,一般都以某 一两种生活型为主,各拥有较丰富的植物种类。 表4-1 不同群落类型的生活型谱 热带和亚热带湿润森林均以高位芽植物占优势,如对高位芽植物作进一步划分即可比较出差异。 对每一群落,均可作叶级的分析,并作出叶级谱。不同植被类型的叶级谱都有一定的规律性,即往往以某一叶级占优势,并以此与其他类型相区别(表4-2 )。叶面积的大小,与气候带有某种相关性。在热带地区,大叶的比例最高,随着逐渐离开赤道,叶面积较小的类型亦渐增多,而大叶的比例逐渐减少。 表4-2 不同群落类型叶级的比较 表4-3 贝加尔针茅草原生物学类群与生态类群综合分析表 (据内蒙古植被,1985,简化) 对生活型与生态类群组成的综合分析,能够更好地反映群落与所处环境的关系。对贝加尔针茅(Stipa baicalensis )草原作此类分析(内蒙古植被,1985)可显示出它的群落重要属性,即中旱生和旱中生的杂类草(双子叶植物)占有很大比重,也表明它的所在地环境并非十分干旱。 以上只涉及生活型内所含种数的多少,进一步分析还需考虑它们在群落中的各种数量特征,才能判别其所起的作用。 二、植物群落的空间结构和植物环境 群落中的各种植物,在群落内占据一定的生存空间,而全部植物(按所属生活型)的分布状况,构成了植物群落垂直的和水平的结构,并将原有生境改变为特殊的群落内部环境(植物环境)。
环境污染对生物的影响 环境污染是指有害物质或因子进入环境,并在环境中扩散、迁移、转化,使环境结构和功能发生变化,对人类以及其他生物的生存和发展产生不利影响的现象。环境污染有不同的类型,按环境要素可分为大气污染、水体污染和土壤污染。 造成环境污染的污染物发生源称之为污染源。污染源又可分为工业污染源、农业污染源、交通运输污染源、生活污染源等。其中工业污染源是指在原料生产、加工过程、燃烧过程、燃烧过程、加热冷却等过程中所使用的生产设备都有可能成为工业污染源。农业污染源是指在农业生产过程中对环境造成有害影响的农田和各种农业设施。交通运输污染源是指对周围环境造成污染的交通运输设施和设备。这类污染源是运行中发出噪声引起振动、运载的有害物的泄露、汽油柴油煤油燃料燃烧等。生活污染源主要是城市化造成的,由于城市人口密集,是人类消费活动集中地。消费能源排出废气可以造成大气污染,排出的生活污水(包括粪便)可以造成水体污染,城市排出的厨房垃圾、废塑料废纸、金属、煤灰可以造成环境污染。 污染物排入环境后经过环境的迁移、分布、扩散、转化,并通过不同的途径进入生物机体。污染物进入生物机体后,经过生物体内的代谢,一些污染物被代谢成无毒的物质排出体外,另一些污染物或一些污染物的代谢产物通过在生物体内浓缩积累和放大,对生物产生不利影响。污染物对生物机体的最早作用是从生物大分子开始的,然后逐步在细胞----器官----个体---种群----群落----生态系统各个水平上反
映出来。 (1)污染物在生物化学和分子水平上的影响 ①污染物对生物体酶的影响 ◆污染物对酶辅助因子的影响 一些污染物能与酶的辅助因子相互作用,从而使辅助因子失活,影响到酶的活性。例如氰化物等能与细胞色素酶中的铁离子相互结合,形成稳定的络合物,而抑制细胞色素的酶活性,使其不能传递电子,则细胞内的氧化代谢过程中断,使机体不能利用氧,出现窒息性缺氧。 ◆对酶活性中心的影响 污染物还能和酶的其他活性基团结合。例如,汞和砷与某些酶的活性基团结合就很牢固,从而使酶失去活性。 ◆破坏酶的结构 有些污染物能取代酶分子中的某些成分,从而使酶失去活性。 ◆与酶激活剂作用 有些酶需要激活剂才能表现出活性,酶激活剂往往是金属离子,凡是能与激活剂作用的污染物都能抑制酶的活性。 ◆污染物与基质竞争同种酶而抑制酶的作用 污染物与底物具有相似的结构,也能和酶形成复合物。从而与底物竞争没得活性中心。 ②污染物对生物大分子的影响 污染物对生物大分子的影响主要表现在以下方面:
环境污染危害动植物生存 一、水体富营养化 富营养化对生物的危害富营养化是指因水体中N 、P 等植物必需的矿质元素含量过多而使水质恶化的现象。水体中含有适量的N 、P 等矿质元素,这是藻类植物生长发育所必需的。但是,如果这些矿质元素大量地进入水体,就会使藻类植物和其他浮游生物大量繁殖。这些生物死亡以后,先被需氧微生物分解,使水体中溶解氧的含量明显减少。接着,生物遗体又会被厌氧微生物分解,产生出硫化氢、甲烷等有毒物质,致使鱼类和其他水生生物大量死亡。发生富营养化的湖泊、海湾等流动缓慢的水体,因浮游生物种类的不同而呈现出蓝、红、褐等颜色。富营养化发生在池塘和湖泊中叫做“水华”,发生在海水中叫做“赤潮”。工业废水、生活污水和农田排出的水中含有很多N 、P 等植物必需的矿质元素,这些植物必需的矿质元素大量地排到池塘和湖泊中,会使池塘和湖泊出现富营养化现象。池塘和湖泊的富营养化不仅影响水产养殖业,而且会使水中含有亚硝酸盐等致癌物质,严重地影响人畜的安全饮水。 二、重金属污染 重金属对生物的危害有些重金属如Mn 、Cu 、Zn 等是生物体生命活动必需的微量元素,但是大部分重金属如Hg 、Pb 等对生物体的生命活动有毒害作用。生态环境中的Hg 、Pb 等重金属,同样可以通过生物富集作用在生物体内大量浓缩,从而产生严重的危害,科学家们发现,自然界中的Hg 在水体中经过微生物的作用,能够转化成毒性更大的甲基汞。在被甲基汞污染了的海水中,藻类植物改变了颜色,海鱼也大量死亡。科学家们还发现,质量浓度仅为4mg/L 的PbCl 2 溶液,就能明显地抑制菠菜和番茄正常地进行光合作用。可见,Hg 、Pb 等重金属对于生物的正常生命活动是十分有害的。 三、大气污染 大气污染对动物的危害与对人体的危害情况相似。动物和人类共同生存在一个大气环境里。大气污染对人类的伤害,动物也不能幸免于难。凡是对人造成严重危害的大气污染事件,对动物也产生同样的危害和影响。空气污染对动物的危害,除污染物的直接侵入造成伤害之外,还通过污染食品进入体内,导致发病和死亡。因为动物没有能力去选择和鉴别某些剧毒性的食品,所以它们将比人类更容易遭受污染物的伤害和影响。 大气污染物,尤其是二氧化硫、氟化物等对植物的危害是十分严重的。当污染物浓度很高时,会对植物产生急性危害,使植物叶表面产生伤斑,或者直接使叶枯萎脱落;当污染物浓度不高时,会对植物产生慢性危害,使植物叶片褪绿,或者表面上看不见什么危害症状,但植物的生理机能已受到了影响,造成植物产量下降,品质变坏。 四、农药污染 许多农药是不易分解的化合物,被生物体吸收以后,会在生物体内不断积累,致使这类有害物质在生物体内的含量远远超过在外界环境中的含量,这种现象称为生物富集作用。生物富集作用随着食物链的延长而加强。例如,几十年前DDT 作为一种高效农药,曾经广泛用于防治害虫。美国某地曾经使用DDT 防治湖内的孑孓,使湖水中残存有DDT ,而浮游动物体内DDT 的含量则达到湖水的一万多倍。小鱼吃浮游动物,大鱼又吃小鱼,致使DDT 在这些大鱼体内的含量竟高达湖水的八百多万倍。
辽东山地老秃顶子冰缘地貌植物群落类型及结构特征 摘要: 关键词:老秃顶子;冰缘地貌;群落类型;结构特征 引言 冰缘地貌是由冻胀、热融蠕流、热融、雪蚀、风力等寒冻风化和冻融作用形成的地表形态. 一般指无冰川覆盖的气候严寒地区的冻土分布区和季节冻土区发育的冰缘地貌, 因而冰缘地貌又称冻土地貌[ 1]辽东地区冻土环境属于季节性冻土,冰期内地表受到强烈的风化作用,使得山体岩层崩解分离,散落的岩块、碎屑常堆积于缓坡或谷地,形成杂乱无章的乱石堆积,故此,当地人也将该地貌称为“乱石窖”或“跳石塘”。目前植被状况良好,苔藓、地衣、蕨类和灌木覆盖严密,乔木根系固定在岩石缝隙之间,郁闭度在0.7以上,给公顷蓄积量达300立方米。但这种特殊石质生境上的森林植被一旦遭到破坏,不仅恢复困难,还潜在着发生泥石流、啸山等重大地质灾害的危险。加之本区正好处在我国东部2~3级生态敏感带的边缘,生态环境脆弱,存在着生态环境的不稳定性。因此,掌握冰缘地貌上覆植被的分布规律,对提高植物群落的稳定性、丰富性。目前国内外对于冰缘地貌与植被生态学之前的关系研究还比较少见,1983年陈鹏等研究了长白山冰缘地貌与土壤动物之间的关系。【6】对于辽东山地老秃顶子冰缘地貌的研究仅限于对其冰缘地貌的形成和冰缘地貌本身单方面的研究,还没有把辽东山地老秃顶子冰缘地貌与其植被生态学联系到一起进行研究。本文通过对老秃顶子冰缘地貌上的植物群落类型及结构特征的研究,旨在揭示其冰缘地貌上的植物类型,分布、结构等特征与其所在冰缘地貌环境之间的关系,从而为冰缘地貌上的植被保护提供基础的科学依据。 1研究区概况 老秃顶子国家级自然保护区(124°41′13″~125°5′15″ E;41°11′11″~41°21′34″ N)位于辽宁省东部桓仁、新宾两县交界处,主峰老秃顶子位于辽宁老秃顶子自然保护区核心区内,海拔1367.3m,相对高差867m,为辽宁第一峰。其四周群山起伏,各山间谷地均有大量冰缘地貌分布。土壤类型主要以棕壤和暗棕壤为典型代表,多由花岗岩残积母质发育而成,结构疏松,有机质含量高,适宜植被生长【2】。地带性植被为温性落叶阔叶林,植物区系属长白植物区系的西南边缘,兼有华北植物区系的过渡性,生物多样性丰富,查明有真菌植物50 科 44 种 , 地衣植物 13 科 84 种 , 苔鲜植物 50 科 204 种 , 维管束植物 120 科 1141 种。维管束植物中有蕨类植物 19 科 60 种 , 裸子植物4 科23 种, 被子植物97 科1058 种[ 24 ], 主要优势科有菊科(Compositae) 、蔷薇科 (Rosaceae) 、毛茛科(Ranun2culaceae) 、禾本科( Gramineae) 、百合科(Liliaceae) 、豆科(Leguminosae) 、伞形科(Api2aceae) 、杨柳科(Salicaceae) 、莎草科(Cyperaceae) 、唇形科(Labiatae) 等。区内气候类型属于北温带大陆性季风气候,年均温 6.3℃,年降水量为870--1060mm,年平均相对湿度67%【4】;年无霜期133d,冻结期114d,年平均冻土厚度91cm。境内水系发达、河流网布,局部乱石窖之下形成暗河,在巨石
影响植物生长的环境因素有很多。这个问题很复杂~~~通俗地讲就是内在和外在、生物非生物……简单说就是植物个体本身受遗传的影响,外在就是来自于周围的微环境和大环境,大环境就是所谓的气候(光热水风温湿度)、地域(地貌土质土壤类型营养含量等)、,微环境就包括的更细了。除了非生物的干扰外,还受其他植物的干扰,比如植物的他感作用就是通过自身分泌特殊的化学或者生物成分,是其他植物不能正常生长~~还有植物的竞争,生态位相同的植物不等良好的生长~~同时还要考虑微生物的作用……还可以分为包括生物因素和非生物因素.生物因素是指影响它的其它生物.非生物因素如:阳光,空气,水.食物等. 植物的根吸收营养物质,要消耗能量的,而能量的主要来源就是根细胞的有氧呼吸,此过程需要一定得氧气,土壤板结,土壤通透性不好,氧气不足,妨碍植物根的呼吸作用,不利于植物生长。而且土壤板结,土壤肥力状况也不理想,土壤孔隙都不理想,土壤团粒结构少,这些影响土壤微生物活动的同时,也会影响植物生长。 首先需要弄明白的是,生物包括微生物、动物、植物。生物对环境的影响有很多,包括好的和坏的。比如说,土壤里面的蚯蚓,在土壤中的活动,使得土壤中的有机物质得以较快分解成植物能够吸收的养分,另外还有疏松土壤的功能。植物通过吸收二氧化碳,释放氧气。微生物包括各种细菌、病毒,这些东西可以使得枯落物以及动物排泄物及尸体能更快的分解,被环境吸收,重新利用。生物对环境的影响无处不在。人类对环境的影响是最大的,但是我们现在对环境都是负面影响,比如破坏植被、污染大气、污染河流等等,这样造成的后果就是对整个生态环境的破坏,不能使生态系统正常发挥作用,最后祸及人类自身。
环境污染对生物多样性的影响 生物多样性是人类社会赖以生存和发展的环境基础,也是当今国际社会环境和发展研究的热点问题之一。中国是全球生物最丰富的国家之一,生物物种数量约占全球的十分之一,是全球生物多样性保护的重要地区。但是由于自然、人为及制度方面的原因,中国的生物多样性正遭受着严重的损失和破坏,保护生物多样性已成为亟待解决的生态保护的头等大事。环境污染对生物多样性的影响主要表现在这三个方面:遗传多样性的丧失、物种多样性的丧失、生态多样性的丧失及生态系统复杂性的降低。随着科学技术水平的发展和人民生活水平的提高,环境污染也在增加,特别是发展中国家,环境污染亦直接或间接对生物造成了一定的影响。 环境污染并不是一种一般意义上的环境胁迫,生物对污染的适应机制及进化格局与自然胁迫条件下的情形并不相同。环境污染是一种生态破坏! 随着研究工作的不断深入和系统化, 人们越来越发现, 环境染污引起物种丧失实质上也是一种生态破坏, 而且认为当今世界正在经历的物种大灭绝在很大程度上与全球扩散的环境污染有密切联系。以下分别在遗传、物种和生态系统水平上讨论污染对生物多样性的影响。 一、遗传多样性的丧失 遗传多样性强调的是现有种质的遗传变异库存量,它既是生物遗传变异的历史积累, 反映了生物的进化过程, 也是现有生物适应现有环境和未来未知环境的遗传基础。遗传多样性的丧失包括已有的遗传基因库的减小和新的遗传变异来源的降低。遗传变异性的丧失会导致生物对未来环境适应性的降低, 从而意味着人类进一步发展所依托的生物资源的减少。污染条件下遗传多样性水平降低可能有以下3个方面原因: 1)在污染条件下,种群的敏感性个体消失, 从而整个种群的遗传多样性水平降低; 2) 污染引起种群的规模减少, 由于随机的遗传漂变, 降低了种群的遗传多样性水平; 3)污染引起种群数量减小, 以至达到了种群的遗传学瓶颈,即使种群最后实现了完全的适应, 并恢复到原来的种群数量时, 由于建立者效应(founder effects) ,从而造成遗传来源单一, 遗传变异性的来源也大大降低。 $ 二、物种多样性的丧失 环境污染引起物种多样性降低的机理一般为: 1)污染物的直接毒害作用, 阻碍生物的正常生长发育, 使生物丧失生存或繁衍的能力。例如,由于农药的大量使用,在杀灭对农作物有害昆虫的同时, 也杀灭了一些对农作物有益的昆虫。 2) 污染引起生境的改变, 使生物丧失了生存的环境。例如,昆明滇池从50年代到90年代,由于水体污染导致富营养化,高等水生植物种类丧失了36%, 鱼类种类丧失了25%,整个湖泊的物种多样性水平显着降低,生态系统的结构趋于单一。 3)污染物在生态系统中的富集和积累作用, 使食物链后端的生物, 难以存活或繁育。以美国长岛河口区生物对DDT的富集为例, 该地区大气中的DDT含量为3×10- 6mg/kg, 其中溶于水中的更微乎其微。但水中浮游生物体内的DDT含量为0. 04mg/ kg, 富集系数为1. 3万( 以大气中DDT 含量作基数); 浮游生物为小鱼所食, 小鱼体内DDT 增加到kg, 富集16. 7万倍; 其后小鱼为大鱼所食, 大鱼体内DDT浓度增加到2mg/ kg, 富集系数为833万;海鸟捕食鱼, 其体内DDT增加到25mg/kg, 富集系数高达858万。从这个例子可以看出, 空气和水中的DDT 含量很低, 但在生态系统中由于通过食物链逐级富集, 使处于食物链后端的鸟类体内DDT含量大增,导致中毒或死亡。在污染引起物种多样性丧失的研究中,除了应了解物种总的数量动态变化外, 还应注意不同物种对于污染的耐性或抗性水平不同, 从而在同样的污染条件下, 幸存的物种还具有一定的区系或种属特点。一般来说,广域分布的物种生存的机会