
实验报告
说明:1、实验报告务必独完成,对抄袭者将按不及格处理;
2、实验报告的格式请按下面的各项要求来填写,不要改动;
3
、正文字体统一用“仿宋-GB2312”、,小四号,单倍行距,小标题加黑;
4、下面的“替换这里”字体底纹在完成后去除;
5、实验报告按时上传,上传时文件名统一按照网上说明来命名;
实验名称:神经干动作电位的引导和观察/动作电位传导速度的测定同组姓名:实验日期:
室温:气压:
成绩:教师:
一、实验结果
(一)神经干动作电位的引导和观察
(二)动作电位传导速度的测定
姓名:学号:
二、分析与讨论
分析:
(一)神经干动作电位的引导和观察
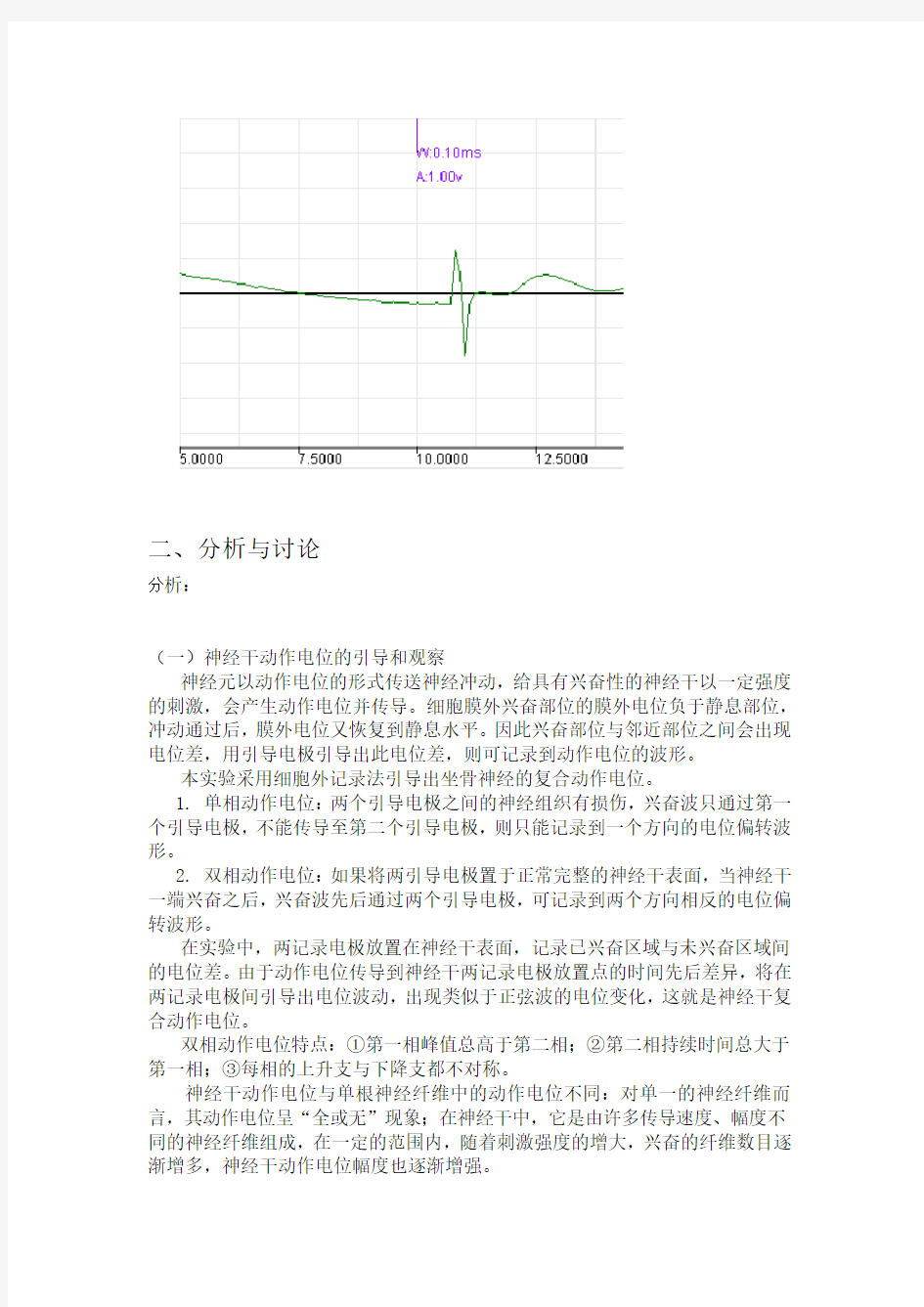
神经元以动作电位的形式传送神经冲动,给具有兴奋性的神经干以一定强度的刺激,会产生动作电位并传导。细胞膜外兴奋部位的膜外电位负于静息部位,冲动通过后,膜外电位又恢复到静息水平。因此兴奋部位与邻近部位之间会出现电位差,用引导电极引导出此电位差,则可记录到动作电位的波形。
本实验采用细胞外记录法引导出坐骨神经的复合动作电位。
1. 单相动作电位:两个引导电极之间的神经组织有损伤,兴奋波只通过第一个引导电极,不能传导至第二个引导电极,则只能记录到一个方向的电位偏转波形。
2. 双相动作电位:如果将两引导电极置于正常完整的神经干表面,当神经干一端兴奋之后,兴奋波先后通过两个引导电极,可记录到两个方向相反的电位偏转波形。
在实验中,两记录电极放置在神经干表面,记录已兴奋区域与未兴奋区域间的电位差。由于动作电位传导到神经干两记录电极放置点的时间先后差异,将在两记录电极间引导出电位波动,出现类似于正弦波的电位变化,这就是神经干复合动作电位。
双相动作电位特点:①第一相峰值总高于第二相;②第二相持续时间总大于第一相;③每相的上升支与下降支都不对称。
神经干动作电位与单根神经纤维中的动作电位不同:对单一的神经纤维而言,其动作电位呈“全或无”现象;在神经干中,它是由许多传导速度、幅度不同的神经纤维组成,在一定的范围内,随着刺激强度的增大,兴奋的纤维数目逐渐增多,神经干动作电位幅度也逐渐增强。
(二)动作电位传导速度的测定
神经组织在接受一次刺激产生兴奋后,其兴奋性将会发生规律性的变化,依次经过绝对不应期、相对不应期、超常期和低常期,然后回到正常水平。
坐骨神经干包括多种类型的神经纤维,记录到的动作电位是它们电位变化的总和。动作电位在神经纤维上的传导有一定的速度,不同类型的神经纤维动作电位传导速度各不相同,其传导速度取决于神经纤维的直径、有无髓鞘等因素。可通过测定神经冲动在神经干上传导的距离(d)与通过这段距离所需的时间(t),然后根据V=d/t即可求出神经冲动的传导速度。
本实验采用两个通道同时记录由两对引导电极记录下的动作电位来计算动作电位传导速度。先分别测量从刺激伪迹到两个动作电位起始点的时间,设上线为t1,下线为t2,求出t2~t1的时间差值。然后再测量标本屏蔽盒中两对引导电极起始电极之间的距离d,则神经冲动的传导速度V=d/(t2~t1)。
通过实验,我们测得t1=2330.76ms,t2=2330.79ms,d=1cm,V=333m/s。
思考与讨论:
1、为什么两对引导电极相距越远,测定的神经纤维兴奋传导速度就越准确?
答:动作电位有一定的时程,当两个电极间的距离没达到足够远时,上相动作电位复极未完成,下相除极已开始,会出现双相动作电位上下相幅值不等,上相幅值较大。两电极间距离大,超过动作电位的波长,则记录到的是对称的双相动作电位波形。其次,距离越大测量的误差就越小,可以减小系统误差。
2、蛙类坐骨神经干主要以Aα类纤维为主,传导速度(V)大约为35~40 m/s,而我们实测得的为333m/s,数值偏差比较大,对此思考其原因?
答:计算结果比理论值偏高,可能是实验中我们将神经干搭在引导电极上时,未将神经干拉成直线,神经干有点下垂或斜放,实际的d>1cm, 使数值偏大。此外,实验仪器和信息处理系统的误差也是造成结果偏差的主要原因。
小结:
1、制备标本时,神经纤维应尽可能长一些,将附着于神经干上的结缔组织膜及血管清除干净,但不能损伤神经干。
2、神经干不能与标本盒壁相接触,也不要把神经干两端折迭放置在电极上,以免影响动作电位的波形。
3、将神经干搭在引导电极上时,尽量将神经干拉成直线,且无下垂或斜向置放,这样会影响测量神经干长度的准确性,最终影响传导速度的准确性。
4、测定动作电位传导速度时,如果神经干长度足够,则尽量将两对引导电极的距离拉远一些,距离越远,测定的传导速度就越准确。
5、在实验中,电位的图像倒置,对此可通过视图→当前通道信号反相,以改变图像。
三、结论
神经干受刺激后,记录的动作电位波形呈双相。神经干动作电位是复合动作电位,相的上升支与下降支都不对称。
本实验测得的神经干动作电位的传导速度为333m/s,较理论值偏大。
(注:文档可能无法思考全面,请浏览后下载,供参考。可复制、编制,期待你的好评与关注)
神经干动作电位、传导速度及不应期的测定 【目的和原理】 神经纤维的兴奋表现为动作电位的产生和传导,神经纤维上传导的动作电位通常称为神经冲动。在神经细胞外表面,已兴奋部位带“负电”,未兴奋部位带“正电”,用引导电极引导出此电位差,输入到示波器,则可记录到动作电位的波形。本实验用细胞外记录法,可引导出坐骨神经的复合动作电位。 神经纤维兴奋的标志是产生一个可以传导的动作电位,它依局部电流或跳跃传导的方式沿神经纤维传导。其传导速度取决于神经纤维的直径、内阻、有无髓鞘等因素,可用电生理学方法来记录和测量。 神经纤维在一次兴奋过程中,其兴奋性可发生周期性变化,包括绝对不应期、相对不应期、超常期和低常期。本实验主要目的是学习电生理仪器的使用方法,掌握离体神经干动作电位的细胞外记录法及其基本波形的判断和测量。掌握神经干动作电位传导速度及其不应期的测定方法,通过调整条件刺激和测试刺激之间的时间间隔,来测定坐骨神经干的绝对不应期。 【实验对象】 蟾蜍或蛙。 【实验器材和药品】 蛙类手术器械一套、电子刺激器、示波器(或计算机实时分析系统)、神经屏蔽盒、任氏液。 【实验步骤】 1.制备坐骨神经——胫、腓神经标本操作方法详见3.8。 2.连接装置(见图8-1-1)。 3.准备仪器: (1)刺激器:调节刺激器各项参数:刺激方式连续刺激,频率16Hz,刺激强度0.5v,波宽0.1ms。调节延迟使动作电位的图像位于示波器荧光屏的中央。 (2)示波器:灵敏度:1~2mv/cm,扫描速度:1~2ms/cm,引导电极输入到示波器的“AC”端,双边输入,刺激器的“同步输出”接示波器“外触发输入”,触发选择设置为“同步触发”。 4.观察项目:
静息电位和动作电位产生的具体原因 伴随生命活动的电现象,称为生物电。关于生物电在生命活动中所起的作用,目前还不十分清楚。本节着重以神经纤维为例讨论细胞水平生物电的表现形式,即静息电位和动作电位。 一、静息电位及其产生机制 (一)静息电位 静息电位是指细胞在安静状态下,存在于细胞膜的电位差。这个差值在不同的细胞是不一样的,就神经纤维而言为膜外电位比膜内电位高70~90mv。如规定膜外电位为0,则膜内电位当为负值(-70~-90mv)。细胞在安静状态时,保持比较稳定的外正内负的状态,称为极化。极化状态是细胞处于生理静息状态的标志。以静息电位为准,膜内负电位增大,称为超极化。膜内负电位减小,称为去或除极化。细胞兴奋后,膜电位又恢复到极化状态,称为复极化。 (二)静息电位产生的机制 “离子学说”认为,细胞水平生物电产生的前提有二:①细胞内外离子分布和浓度不同。就正离子来说,膜内K 浓度较高,约为膜外的30倍。膜外Na 浓度较高约为膜内的10倍。从负离子来看,膜外以Cl-为主,膜内则以大分子有机负离子(A-)为主。②细胞膜在不同的情况下,对不同离子的通透性并不一样,如在静息状态下,膜对K 的通透性大,对Na 的通透性则很小。对膜内大分子A-则无通透性。 由于膜内外存在着K 浓度梯度,而且在静息状态下,膜对K 又有较大的通透性(K 通道开放),所以一部分K 便会顺着浓度梯度向膜外扩散,即K 外流。膜内带负电荷的大分子A-,由于电荷异性相吸的作用,也应随K 外流,但因不能透过细胞膜而被阻止在膜的内表面,致使膜外正电荷增多,电位变正,膜内负电荷增多,电位变负。这样膜内外之间便形成了电位差,它在膜外排斥K 外流,在膜内又牵制K 的外流,于是K 外流逐渐减少。当促使K 流的浓度梯度和阻止K 外流的电梯度这两种抵抗力量相等时,K 的净外流停止,使膜内外的电位差保持在一个稳定状态。因此,可以说静息电位主要是K 外流所形成的电一化学平衡电位。 二、动作电位及其产生机制 (一)动作电位 细胞受刺激时,在静息电位的基础上发生一次短暂的扩布性的电位变化,这种电位变化称为动作电位。 实验观察,动作电位包括一个上升相和一个下降相(图2-3)。上升相代表膜的去极化过程。以0mv电位为界,上升相的下半部分为膜的去极化,是膜内负电位减小,由-70~-90mv.变为0mv;上升相的上半部分是膜的反极化(超射),是膜电位的极性发生倒转即膜外变负,膜内变正,由0mv上升到20~40mv。上升相膜内电位上升幅度约为90~130mv。下降相代表膜的复极化过程。它是膜内电位从上升相顶端下降到静息电位水平的过程。由于动作电位幅度大、时间短不超过2ms,波形很象一个尖峰,故又称峰电位。在峰电位完全恢复到静息电位水平之前,膜两侧还有微小的连续缓慢的电变化,称为后电位。 (二)动作电位产生的机制 动作电位产生的机制与静息电位相似,都与细胞膜的通透性及离子转运有关。 l.去极化过程当细胞受刺激而兴奋时,膜对Na 通透性增大,对K 通透性减小,于是细胞外的Na 便会顺其波度梯度和电梯度向胞内扩散,导致膜内负电位减小,直至膜内电位比膜外高,形成内正外负的反极化状态。当促使Na 内流的浓度梯度和阻止Na 内流的电梯度,这两种拮抗力量相等时,Na 的净内流停止。因此,可以说动作电位的去极化过程相当于Na 内流所形成的电一化学平衡电位。 2.复极化过程当细胞膜除极到峰值时,细胞膜的Na 通道迅速关闭,而对K 的通透性增
人体解剖及动物生理学实验报告 神经干复合动作电位 【实验题目】 神经复合动作电位 1、蟾蜍坐骨神经干复合动作电位(CAP)阈值和最大幅度的测定 2、蟾蜍坐骨神经干复合动作电位(CAP)传导速度的测定 3、蟾蜍坐骨神经干复合动作电位(CAP)不应期的测定 【实验目的】 确定蟾蜍坐骨神经干复合动作电位(CAP)的 1、临界值和最大值 2、传导速度 3、不应期(包括绝对不应期和相对不应期) 【实验原理】 神经系统对维持机体稳态起着重要作用,动作电位(AP)是神经系统进行通信联系所采用的信号。多个神经元的轴突集结成束形成神经,APs沿感觉神经经外周传向中枢或沿运动神经由中枢传向外周。坐骨神经干由上百根感觉神经和运动神经组成,分别联系腿部的感受器和效应器(骨骼肌)。如果电刺激一根离体的坐骨神经干,通过细胞外引导方式,就能记录到神经干复合动作电位(CAP)。一个CAP是一系列具有不同兴奋性的神经纤维产生的多个AP的总和。刺激强度越大,兴奋的神经纤维数目就越多,CAP的幅度也就越大。与胞内引导得到的单细胞AP相比,CAP是双相电位,逐级递增(非全或无),并且幅度较小。 阈电位是指一个刚刚能观测到的CAP,所对应的刺激为阈刺激。在一定范围内增加刺激强度,CAP幅度相应增大。最大CAP所对应的最小刺激电位即最大刺激。 动作电位可以沿神经以一定的速度不衰减地传导,传导速度的快慢基于多种因素,这些因素决定了生物体对其坏境的适应性。它们包括神经的直径、有无髓鞘、温度等等。
神经在一次兴奋过程中,其兴奋性将发生一个周期性的变化,最终恢复正常。兴奋的周期性变化,依次包括绝对不应期、相对不应期等等。绝对不应期内,无论多么强大的刺激都不能引起神经再一次兴奋;相对不应期内,神经兴奋性较低,较大的刺激能够引起兴奋。绝对不应期决定了神经发放冲动(动作电位)的最高频率,保证了动作电位不能叠加(区别于局部电位),以及单向传导(只能有受刺激部位向远端传导,不能返回)的特性。不应期的产生依赖于细胞膜上特定离子通道的特点,如钠、钾离子通道。 【实验方法】 1、制作蟾蜍坐骨神经干标本 (1)双毁髓处死蟾蜍后,剥去皮肤,暴露腰骶丛神经,游离大腿肌肉之间的坐骨神经干及其下行到小腿的两个分支:胫神经和腓神经,三段结扎,剪去无关分支后离体。注意保持神经湿润。 (2)将神经搭于标本盒内,保证神经与电极充分接触,中枢端接触刺激电极S1和S2,外周端接触记录电极R1-R5,之间接触接地电极。 (3)刺激输出线两夹子分别连接标本盒的刺激电极S1和S2,插头接生物信号采集系统RM6240的刺激输出插口;信号输入倒显得红色和绿色夹子分别连接记录电极(绿色夹子在前,引导出正向波形,即出现的第一个波峰向上),黑色夹子连接接地电极,插头接通道A、蟾蜍坐骨神经干复合动作电位(CAP)临界和最大幅度的确定 (1)打开信号采集软件,从“实验”菜单中选取“神经干动作电位”,出现自动设置的界面,各项参数已设置好,界面中只有一个采集通道,对应仪器面板上的通道1(因此信号输入线应连接在通道1)。 (2)检查装置链接正确,确定装置是否正常工作,以及神经是否具有活性。采用刺激强度1V,刺激时程0.2ms,延时5ms,刺激模式为单刺激。选择“同步触发”,按下“开始刺激”后,正常情况下屏幕上会出现一个双相电位即CAP。 (3)降低刺激强度,确定CAP的阈电位。记录刺激阈值及CAP幅度(波峰与波谷之间的差值)。 (4)以0.05V或更小的间隔,逐渐增大刺激强度,观察CAP幅度的变化,同时,记录刺激电位及对应的CAP幅度,直到CAP达到稳定,即最大值(神经标本在正常生理活性时,1V 以内的刺激强度即可引起最大的CAP)。
动作电位微专题复习 教学反思:动作电位有关的知识是高考的高频考点,也是教学的重点和难点,需要进一步进行微专题复习。 1.动作电位产生的机制 (1)阈刺激或阈上刺激使膜对Na+的通透性增加,Na+顺浓度梯度及电位差内流,使膜去极化,形成动作电位的上升支。 (2)Na+通道失活,而K+通道开放,K+外流,复极化形成动作电位的下降支。 2.动作电位的测量 静息电位常见的测定方式是将电流表的两个电极一个放在神经纤维的外侧,另一个放在神经纤维的内侧,由于内外两侧存在电势差,因此电流表指针会发生偏转。 在一个神经纤维上的测定:是指将电流表的两个电极放在同一个神经纤维的外侧(A处和B 处),来测定两个电极处是否有电位差。 3.动作电位产生的影响因素 主要是Na+的平衡电位,此外,其它离子如Ca2+和Cl-,离子通道阻断剂,细胞的代谢等因素。 4.动作电位的传导 动作电位的传导实际上就是兴奋膜向前移动的过程。在受到刺激产生兴奋的轴突与周围静息膜之间都可以产生局部电流,因此可以向两个方向传导,被称之为动作电位的双向传导。动作电位在传导过程中是不衰减的,其原因在于动作电位在传导时,实际上是去极化区域的移动和动作电位的逐次产生,每次产生的动作电位幅度都接近于钠离子的平衡电位,可见其传导距离与幅度是不相关的,因此动作电位幅度不会因传导距离的增加而发生变化。 神经纤维的传导速度极快,但不同的神经纤维的传导速度变化很大。例如,人体的一些较粗的有髓纤维传导速度可达100m/s,而某些较细的无髓纤维的传导速度甚至低于1m/s。 光在空气中的速度:
电流速度为什么就和光速相等 电流是以电场的方式传递的,就是光速.但导线中电子的速度却是很慢的. 在金属导线中,电能的传输速度是每秒三十万公里,与光速同,而我们在大型直线加速器中只能把电子加速到接近光速,其质量已达电子静止质量的四万倍以上,消耗的能量够一座小城镇的用量.从重力场理论中知道,光速是光能传导速度,是能量空间的调整速度,电流速度就是电能传导速度. “电”的传播过程大致是这样的:电路接通以前,金属导线中虽然各处都有自由电子,但导线内并无电场,整个导线处于静电平衡状态,自由电子只做无规则的热运动而没有定向运动,当然导线中也没有电流.当电路一接通,电场就会把场源变化的信息,以大约光速的速度传播出去,使电路各处的导线中迅速建立起电场,电场推动当地的自由电子做漂移运动,形成电流.那种认为开关接通后,自由电子从电源出发,以漂移速度定向运动,到达电灯之后,灯才能亮,完全是一种误解.
人体解剖及动物生理学实验报告 实验名称神经干复合动作电位 姓名 学号 系别 组别 同组姓名
实验室温度20℃ 实验日期2015年4月24日 一、实验题目 蟾蜍坐骨神经干复合动作电位(CAP) A蟾蜍坐骨神经干CAP阈值和最大幅度的确定 B蟾蜍坐骨神经干CAP传导速度的确定 C蟾蜍坐骨神经干CAP不应期的确定 二、实验目的 确定蟾蜍坐骨神经干复合动作电位(CAP)的 (1)临界值和最大值 (2)传导速度 (3)不应期(相对不应期、绝对不应期) 三、实验原理 神经系统对维持机体稳态起着重要作用,动作电位(AP)是神经系统进行通信联系所采用的信号,多个神经元的轴突集结成束形成神经,APs沿感觉神经有外周传向中枢或沿运动神经由中枢传向外周。坐骨神经干由上百根感觉神经和运动神经组成,分别联系腿部的感受器和效应器(骨骼肌)。如果电刺激一根离体的坐骨神经干,通过细胞外引导方式,就能记录到神经干复合动作电位(CAP)。一个CAP是一系列具有不同兴奋
性的神经纤维产生的多个AP的总和。刺激强度越爱,兴奋的神经纤维数目就越多,CAP 的幅度也就越大。与胞内引导得到的单细胞AP相比,CAP是双相电位,逐级递增(非全或无),并且幅度较小。 阈电位是指一个刚刚能观测到的CAP,所对应的刺激为阈刺激。在一定范围内增加刺激强度,CAP幅度相应增大。最大CAP所对应的最小刺激电位即最大刺激。 动作电位可以沿神经以一定的速度不衰减地传导,传导速度的快慢基于多种因素,这些因素决定了生物体对其坏境的适应性。它们包括神经的直径、有无髓鞘、温度等等。 神经在一次兴奋过程中,其兴奋性将发生一个周期性的变化,最终恢复正常。兴奋的周期性变化,依次包括绝对不应期、相对不应期等等。绝对不应期内,无论多么强大的刺激都不能引起神经再一次兴奋;相对不应期内,神经兴奋性较低,较大的刺激能够引起兴奋。绝对不应期决定了神经发放冲动(动作电位)的最高频率,保证了动作电位不能叠加(区别于局部电位),以及单向传导(只能有受刺激部位向远端传导,不能返回)的特性。不应期的产生依赖于细胞膜上特定离子通道的特点,如钠、钾离子通道。 四、实验方法 蟾蜍坐骨神经标本的制作 1.双毁髓处死蟾蜍后,剥去皮肤,暴露腰骶丛神经,游离大腿肌肉之间的坐骨神经 干及其下行到小腿的两个分支:胫神经和腓神经,三段结扎,剪去无关分支后离体。注意保持神经湿润。 2. 将神经搭于标本盒内,保证神经与电极充分接触,中枢端接触刺激电极S1和S2, 外周端接触记录电极R1-R2,之间接触接地电极。 3. 刺激输出线两夹子分别连接标本盒的刺激电极S1和S2,插头接生物信号采集系 统RM6240的刺激输出插口;信号输入倒显得红色和绿色夹子分别连接记录电极(绿色夹子在前,引导出正向波形,即出现的第一个波峰向上),黑色夹子连接接地电极,插头接通道1.
【提问】窦房结P细胞跨膜电位及产生机理? 【回答】学员dbss9ffe42,您好!您的问题答复如下:外Ca2+浓度的影响,可被Ca2+通道抑制剂(如维拉帕米、Mn2+)阻断。当膜电位由最大复极电位自动去极化到阈电位时,膜上L型Ca2+抖通道被激活,引起Ca2+。内流,导致0期去极化。 祝您学习愉快! 【追问】那么请问窦房结P细胞的复极化是受什么影响【回答】学员nflalihh,您好!您的问题答复如下: 窦房结细胞的动作电位具有以下特点: ①最大复极电位与阈电位的绝对值小; ②0期去极化的幅度小、时程长、去极化速率较慢; ③没有明显的复极1期和2期; ④4期自动去极化速度快。 1.去极化过程:0期去极L型Ca2+通道激活,Ca2+内流。 2.复极化过程:3期复极L型Ca2+通道逐渐失活,Ca2+内流相应减少,及Ik通道的开放,K+外流增加。 3.4期自动去极化机制:①IK:复极至-60mV时,因失活逐渐关闭,导致K+外流衰减,是最重要的离子基础;②Ica-T:
在4期自动去极化到-50mV时,T型Ca2+通道激活,引起少量Ca2+内流参与4期自动去极化后期的形成;③If:窦房结细胞最大复极电位只有-70mY,If不能充分激活,在P细胞4期自动去极化中作用不大。 【追问】老师这道题还是不明白 【回答】学员zhulipeng,您好!您的问题答复如下:窦房结细胞的生物电特点是没有稳定的静息电位。动作电位复极至3期末进入第4期,便自动缓慢去极。 窦房结的最大舒张电位约-60mV,阈电位约-40mV。 0期去极化速度缓慢,主要是Ca2+缓慢内流引起。复极化无明显的l期和2期平台,随即转入复极化3期,后者主要是K+外流形成。4期的自动去极化主要是由于K+通道逐渐关闭,Na+、Ca2+内流逐渐增多而引起。
人体解剖及动物生理学实验报告实验名称神经干复合动作电位 姓名 学号 系别 组别 同组姓名 实验室温度20℃ 实验日期2015年4月24日一、实验题目 蟾蜍坐骨神经干复合动作电位(CAP) A蟾蜍坐骨神经干CAP阈值和最大幅度的确定 B蟾蜍坐骨神经干CAP传导速度的确定 C蟾蜍坐骨神经干CAP不应期的确定 二、实验目的 确定蟾蜍坐骨神经干复合动作电位(CAP)的 (1)临界值和最大值
(2)传导速度 (3)不应期(相对不应期、绝对不应期) 三、实验原理 神经系统对维持机体稳态起着重要作用,动作电位(AP)是神经系统进行通信联系所采用的信号,多个神经元的轴突集结成束形成神经,APs沿感觉神经有外周传向中枢或沿运动神经由中枢传向外周。坐骨神经干由上百根感觉神经和运动神经组成,分别联系腿部的感受器和效应器(骨骼肌)。如果电刺激一根离体的坐骨神经干,通过细胞外引导方式,就能记录到神经干复合动作电位(CAP)。一个CAP是一系列具有不同兴奋性的神经纤维产生的多个AP的总和。刺激强度越爱,兴奋的神经纤维数目就越多,CAP的幅度也就越大。与胞内引导得到的单细胞AP相比,CAP是双相电位,逐级递增(非全或无),并且幅度较小。 阈电位是指一个刚刚能观测到的CAP,所对应的刺激为阈刺激。在一定范围内增加刺激强度,CAP幅度相应增大。最大CAP所对应的最小刺激电位即最大刺激。 动作电位可以沿神经以一定的速度不衰减地传导,传导速度的快慢基于多种因素,这些因素决定了生物体对其坏境的适应性。它们包括神经的直径、有无髓鞘、温度等等。 神经在一次兴奋过程中,其兴奋性将发生一个周期性的变化,最终恢复正常。兴奋的周期性变化,依次包括绝对不应期、相对不应期等等。绝对不应期内,无论多么强大的刺激都不能引起神经再一次兴奋;相对不应期内,神经兴奋性较低,较大的刺激能够引起兴奋。绝对不应期决定了神经发放冲动(动作电位)的最高频率,保证了动作电位不能叠加(区别于局部电位),以及单向传导(只能有受刺激部位向远端传导,不能返回)的特性。不应期的产生依赖于细胞膜上特定离子通道的特点,如钠、钾离子通道。 四、实验方法 蟾蜍坐骨神经标本的制作 1.双毁髓处死蟾蜍后,剥去皮肤,暴露腰骶丛神经,游离大腿肌肉之间的坐骨神经干及其下行到小腿 的两个分支:胫神经和腓神经,三段结扎,剪去无关分支后离体。注意保持神经湿润。 2. 将神经搭于标本盒内,保证神经与电极充分接触,中枢端接触刺激电极S1和S2,外周端接触记录 电极R1-R2,之间接触接地电极。
讨论神经细胞动作电位形成的机制及影响因素。 动作电位是可兴奋细胞受到刺激时,在静息电位的基础上爆发的一次迅速的,可逆的,并且是可传导的电位变化。 1、神经细胞动作电位形成的机制: ①当细胞受到刺激时,细胞膜上少量Na+通道被激活而开放,Na+顺浓度差,少量内流,导致膜内外电位差下降,产生局部电位。 ②当膜内电位变化到阈电位时,Na+通道大量开放。 ③Na+顺电化学差和膜内负电位的吸引,引发再生式内流。 ④膜内负电位减小到零并变为正电位,形成动作电位(AP)上升支。 ⑤Na+通道关闭,Na+内流停止的同时K+通道被激活而开放。 ⑥由于K+顺浓度差和膜内正电位的吸引,K+迅速外流。 ⑦膜内电位迅速下降,恢复到静息电位(RP)水平,即AP 下降支。 ⑧钠泵的作用,将进入膜内的钠离子泵出膜外同时将膜外多余的钾离子泵入膜内,恢复兴奋前时离子分布的浓度。 2、影响动作电位形成的因素: 主要是Na+的平衡电位,此外,还有其它离子如Ca2+和Cl-,离子通道阻断剂,细胞的代谢等因素。 主要为Na+的平衡电位:
①细胞膜两侧存在离子浓度差,细胞膜内钾离子浓度高于细胞膜外,而细胞外钠离子、钙离子、氯离子高于细胞内,这种浓度差的维持依靠离子泵的主动转运。(主要是钠-钾泵(每3个Na+流出细胞, 就有2个K+流入细胞内。即Na+:K+ =3:2)的转运)。 ②细胞膜在不同状态下对不同离子的通透性不同,例如,安静时主要允许钾离子通透,而去极化到阈电位水平时,又主要允许钠离子通透。 ③可兴奋组织或细胞受阈刺激或阈上刺激。 在细胞膜上任何一点产生的动作电位会不衰减地传播到整 个细胞膜上,这称之为动作电位的传导。如果是发生在神经纤维上,传导的动作电位又称为神经冲动。 以神经元为例,动作电位沿轴突的传导是通过跨膜的局部电流实现的。给轴突的某一位点以足够强的刺激,可使其产生动作电位。此时该段膜内外两侧的电位差发生暂时的翻转,即由安静时膜内为负、膜外为正的状态转化为兴奋时的膜内为正、膜外为负的状态,称其为兴奋膜。 兴奋膜与周围的静息膜(未兴奋的膜)无论在膜内还是膜外均存在有电位差,同时细胞膜的两侧的溶液都是导电的,所以兴奋膜与静息膜之间可发生电荷移动,这种电荷移动就是局部电流。在膜外侧,电流从静息膜流向兴奋膜;在膜内侧,电流由兴奋膜流向静息膜。
2.神经干动作电位是神经兴奋的客观标志,给具有兴奋性的神经干以一定强度的刺激,会产生动作电位并传导。在神经细胞外面,已兴奋部位的膜外电位负于静息部位。当神经冲动通过后,兴奋处的膜外电位又恢复到静息时的水平。所以兴奋部位和邻近部位之间可出现电位差,用引导电极引导出此电位差,输入到示波器,则可记录到动作电位的波形。本实验采用细胞外记录法,可引导出坐骨神经的复合动作电位。 3.经纤维兴奋的标志是产生一个可以传导的动作电位,它以局部电流或跳跃式传导的方式沿神经纤维传导。其传导速度取决于神经纤维的直径、内阻、有无髓鞘等因素。坐骨神经-腓神经为一混合神经干,其动作电位是由一群不同兴奋阈值、传导速度和幅值的电位总和而成,为复合动作电位。蛙类坐骨神经干中以Aa类纤维为主,传导速度大约35~40m/s。测定神经冲动在神经干上传导的距离和通过这些距离所需的时间,即可计算出该神经干兴奋传导的速度。 4.动作电位在神经纤维上的传导有一定的速度。不同类型的神经纤维,其传导速度各不相同,取决于神经纤维的直径、有无髓鞘、环境温度等因素。蛙类坐骨神经干中以Aα类纤维为主,传导速度大约35~40m/s。测定神经冲动在神经干上传导的距离(d)与通过这一距离所需的时间(t),即可根据V=d/t 求出神经冲动的传导速度。 5.神经纤维的兴奋部位相对于未兴奋部位来说呈负电位,两点之间存在电位差,通过单极或双极电极的引导在记录系统上进行显示和分析。由于采用的是胞外记录的方法,因而在单极记录时,测得的动作电位实际上是组成神经干中的每根神经纤维兴奋后的超射值在神经干表面的叠加。即此动作电位是一复合波,其上升相、下降相及峰值不是相应的单一动作电位波形的去极化相、复极化相及峰电位。在双极记录时,测得的波形实际上是两个记录电极的电位差,与单一动作电位波形相差更大,这使问题的分析更加复杂。动物实验制作的坐骨神经 腓肠肌标本中,神经干是由具有不同生理特性的不同种类神经纤维所组成,故复合动作电位记录的是复合波。然而,每种纤维兴奋后传导速度各不一样,波长也各不相等,加上引导方式不同,这也增加了我们分析复合双相动作电位的复杂性及带来传导速度测定的困难。 6.对于单根神经纤维,其兴奋后产生负波。对于某一点,负波的产生和终止不是突然的,而需要一定的时间才能达到最高点,故记录曲线的上升和下降都具有一定的斜率。神经干受刺激后,由于不同神经纤维兴奋产生了不同的负波,它们波长不等,传导速度也不相等,所以
静息电位,动作电位的产生机制及影响其大小的主要因素 一、静息电位(resting potential, RP) 1、概念:静息电位:细胞在静息(未受刺激)状态下膜两侧的电位差称静息电位(膜电位) 2、静息时细胞的特点 静息时细胞内外离子的特点:①细胞内[K+]一般比细胞外液高30倍;②细胞内带负电荷的生物大分子(主要是蛋白质)比细胞外液高10倍;③细胞外液中[Na+]和[CL-]都比细胞内高20倍。所以,细胞内正离子主要为K+,负离子主要为带负电荷的蛋白质分子。细胞外正离子主要为Na+,负离子主要为CL- 。 静息时细胞膜的选择通透性:①带负电荷的蛋白质分子完全不可通过;②Na+和CL-通透性极小;③K+有较大的通透性。3、静息电位形成的机理:细胞内的K+在细胞膜内外浓度差(内高外低)作用下携带正离子外流,当膜内外K+浓度差(K+外流动力)和K+外流所形成的电位差(K+外流阻力)达到动态平衡时,K+的净通量为零,此时所形成的电位差稳定于某一数值而不再增加,即形成静息电位;所以说静息电位实质为K+外流所形成的跨膜电位。细胞内外的K+不均衡分布和静息状态下细胞膜对K+的通透性是细胞在静息状态下保持极化状态的基础。 二、动作电位 1. 动作电位的概念动作电位(action potential):可兴奋组织接受刺激而发生兴奋时,细胞膜原有的极化状态立即消失,并在膜的内外两侧发生一系列的电位变化,这种变化的电位称为动作电位。 2. 动作电位形成的机理 证明:①人工地改变细胞外液Na+浓度,动作电位上升支及其幅度也随之改变,*海水实验; ②用河豚毒阻断Na+通道后,动作电位幅度↓或消失;③膜片钳实验。3.动作电位组成动作电位的扫描波形包括升支和降支两部分。如采用慢扫描并高度放大,则升支和降支的开始部分显示为尖锐的剑锋状,故动作电位又称为锋电位。动作电位的升支代表细胞受到刺激后膜的去极化和反极化过程,即膜内电位由静息时的-70毫伏逐渐减小到-55毫伏(由于这一膜电位可以激发动作电位产生,故把-55毫伏的膜电位称为阈电位);然后,膜电位再减小到0毫伏(去极化结束);最后膜电位由0毫伏迅速上升到+35毫伏(反极化)。通常把膜电位超出0的正值部分称为超射。动作电位的降支代表细胞的复极化过程。在此过程中,膜电位还要发生变化,先出现微弱的去极化,接着出现超极化;前者称为负后电位,后者称为正后电位。负后电位使膜电位减小,临近阈电位而容易被激发动作电位,故也称之为超常期后电位或去极化电位;正后电位使膜电位增大,远离阈电位而不易发生动作电位,故也称之为低常期后电位或超极化后电位。动作电位出现时间与细胞兴奋性变化时间是相吻合的。动作电位的升支所占时间相当于绝对不应期,降支前半段所占时间相当于相对不应期,负后电位所占时间相当于超常期,正后电位所占时间相当于低常期。通常所说的神经冲动,就是指一个沿着神经纤维传导的动作电位或锋电位 1 / 1
神经干动作电位实验报Experimental report of neUhtstem action potential 告 Intern ship report 实验报告
一、实验目的: 1. 学习蛙坐骨神经干标本的制备 2. 观察坐骨神经干的双相动作电位波形,并测定最大刺激强度 3. 测定坐骨神经干双相动作电位的传导速度 4. 学习绝对不应期和相对不应期的测定方法 5. 观察机械损伤或局麻药对神经兴奋和传导的影响 二、实验材料 1. 实验对象:牛蛙 2. 实验药品和器材:任氏液,2%普鲁卡因,各种带USB接口或插头的连接导线,神经屏 蔽盒,蛙板,玻璃分针,粗剪刀,眼科剪,眼科镊,培养皿,烧杯,滴管,蛙毁髓探针,BL-420N 系统 三、主要方法和步骤: 1. 捣毁脑脊髓 2. 分离坐骨神经 3. 安放引导电极 4. 安放刺激电极 5. 启动试验系统 6. 观察记录 7. 保存 8. 编辑输出 四、实验结果和讨论 1.观察神经干双相动作电位引导(单通道,单刺激) 如图,观察到一个双相动作电位波形。
Pm驴:i SQOQOKi 2.0 ms 7 射¥ 也00z 时间 一—j .................... : .................. 频率: 最大值- ...... ' ........ ' ......... [ ........ ;...... [协小值: -15 - -20 _ 1 OOY oo: oo. m兀卫EQ创 2.神经干双相动作电位传导速度测定(双通道,单刺激) -ID kUUUChz L.U ns ZlT m¥ii J.ttmz j ................. ■:- I 2? 1. WV 1 I --------------- 14 I I 4 I I I ooTio mo oa nr iins on oo oru oom coe co nr n o日on m nn oo oo ni2 DO on rtu OO CIJ ri^ oo oc OIA (1) 选择“神经骨骼肌实验”一“…传导速度测定” (2) 改变单刺激强度 (3) 传导速度=传导距离(R1--R2-)/传导时间(t 2-t 1) 如图所示,两个波峰之间的传导时间△ t = (t 2-t 1) = 0.66ms 实验中,我们设定在引导电极1和3之间的距离△ R = (R 1--R2-) = 1cm 故传导速度v = △ R/ △ t = 1cm / 0.66ms = 15.2 m/s 释: 最 大ii; ■小 值: 平均值: 嶂赠但? 面租 BJ祠; 最知宜. 环值: 平均值: 而租
动作电位、静息电位等的产生机制及特征: 静息电位产生的原理是这样的:神经元在静息情况下,细胞膜对K +具有较高的通透性,而对Na +等的通透性很低,并且胞内K +的浓度要远远高于胞外,因此在浓度差的驱动下,K +从胞内流向胞外,而由于K +带有1个正电荷的电量,因此随着K +的流动,膜两侧会形成一个逐渐增大的电位差,这个电位差则会阻止K +进一步进行跨膜扩散。当促进K +向外流动的浓度差与阻止K +向外流动的电位差相等时,离子的净移动就会停止,这是跨膜的电位差称为K +离子的平衡电位(equilibrium potential ),可以根据能斯特(Nernst )方程计算出K +的平衡电位, [K]ln [K]o K i RT E ZF 以上的能斯特方程中,E K 为K +的平衡电位,R 为气体常数,T 为绝对温度,Z 为离子价数,F 为法拉第常数,[K]o 和 [K]i 分别为钾离子在胞外和胞内的浓度,我们将上述参数的值代入后可以计算出K +的平衡电位为-75mV ,而同样的也可以计算出Na +的平衡电位为+55mV 。根据这一能斯特理论,1902年这一静息电位产生机制的“膜假说”被提出了,尽管多数人们接受这一理论,但一直未能得到证实。直到1939年,生物学家Hodgkin 和Huxley 从枪乌贼的巨大神经轴突中第一次精确记录到了静息电位,结果为-60 mV ,与计算推测的K +的平衡电位接近,证实了“膜假说”的可靠性。但实际的静息电位E m 并不完全等于E K ,而是介于E K 和E Na 之间。这说明静息电位的形成主要是K +跨膜流动形成的,但Na +的流动也参与其中。 我们在理解了静息电位产生的机制之后,进一步来探讨动作电位的机制。我们知道电位的变化,归根到底就是膜两侧的离子快速跨膜流动的结果。经过近20年的时间,随着实验技术特别是电压钳、膜片钳(patch clamp technique)等技术的发展,生物学家通过不断的实验研究,才逐渐明确了动作电位的产生机制。动作电位的去极化相是由带正电荷的离子从胞外向胞内移动(例如Na +和Ca 2+的内流)产生的,称为内向电流(inwar current ),相反动作电位的复极化相是由带正电荷的离子(K +)从胞内向胞外移动产生的,称为外向电流(outward current )。但外向电流也可以由带负电荷的离子从胞外流向胞内形成,例如Cl -,那么介导动作电位生成的离子成分是什么?它们是如何被准确控制进行流动的? 最早是由Hodgkin 和Huxley 提出了“钠学说”,由于他们记录到动作电位的峰值达到+50 mV ,非常接近Na + 的平衡电位,因此他们认为在动作电位爆发时,Na + 的一过性内流使得膜电位出现快速、短暂的去极化。而后又设计了
在静息电位的基础上,细胞受到一个适当的刺激,其膜电位所发生的迅速、一过性的极性倒转和复原,这种膜电位的波动称为动作电位。动作电位的升支和降支共同形成的一个短促、尖峰状的电位变化,称为锋电位。锋电位在恢复至静息水平之前,会经历一个缓慢而小的电位波动称为后电位,它包括负后电位和正后电位。 细胞的动作电位具有以下共同特征:①动作电位具有“全或无” 特性,动作电位是由刺激引起细胞产生的去极化过程。而且刺激必须达到一定强度,使去极化达到一定程度,才能引发动作电位。对于同一类型的单细胞来说一旦产生动作电位,其形状和幅度将保持不变,即使增加刺激强度,动作电位幅度也不再增加,这种特性称为动作电位的全或无 ( allornone )现象,即动作电位要么不产生要产生就是最大幅度;②动作电位可以进行不衰减的传导,动作电位产生后不会局限于受刺激的部位,而是迅速沿细胞膜向周围扩布,直到整个细胞都依次产生相同的电位变化。在此传导过程中,动作电位的波形和幅度始终保持不变;③动作电位具有不应期。细胞在发生一次兴奋后,其兴奋性会出现一系列变化,包括绝对不应期、相对不应期、超常期和低常期。绝对不应期大约相当于锋电位期间,相对不应期和超常期相当于负后电位出现的时期;低常期相当于正后电位出现的时期。 (二)动作电位的产生机制 动作电位上升支主要由Na吶流形成,接近于Na啲电-化学平衡电位。 1.细胞内外Na+和K+的分布不均匀,细胞外高Na+而细胞内高K+。 2.细胞兴奋时,膜对Na+有选择性通透,Na+顺浓度梯度内流,形成锋电位的上升支。 3.K+外流增加形成了动作电位的下降支。 在不同的膜电位水平或动作电位发生过程中,Na+通道呈现三种基本
For personal use only in study and research; not for commercial use 神经干动作电位传导速度的测定 实验对象:蟾蜍 一实验目的 掌握坐骨神经标本的制备方法。 掌握引导神经干复合动作电位和测定其传导速度的基本原理。 二相关知识 (一)兴奋及兴奋性的概念 (二)动作电位的潜伏期、动作电位时程和幅值 1、动作电位:各种可兴奋细胞在受到刺激而兴奋时,可以在细胞膜静息电位的基础 上发生一次短暂的,可向周围扩布的电位波动。这种电位波动称为动作电位。(三)、动作电位的传导 局部电流的形式 1、细胞外记录 2、神经干的动作电位 神经干是由许多粗细不等的有髓和无髓神经纤维组成的混合神经,故神经干动作电位与单根神经纤维的动作电位不同,它是由许多神经纤维的动作电位合成的一种复合电位。 三实验原理 (一)、单根神经纤维动作电位的引导及其传导 1、记录出了一个先升后降的双相动作电位的原理 当神经纤维未受刺激时,膜外与电极所接触的两点之间没有电位差,所以两电极之间也无电位差存在,扫描线为一水平基线。在神经干左端给予电刺激后,则产生一个向右传导的冲动(负电位),当冲动传到1电极(负电极)下方时,此处电位较2处为低,产生了电位差,扫描线向上偏转,记录出一个向上的波形(在电生理实验中,为了便于观察,习惯上规定负波向上)。随后,冲动继续向右侧传导,离开1电极传向2电极处。当它到达2电极(正电极)下方时,因1电极处神经差不多已恢复到原来的状态,于是2电极处又较1电极处为负,引起扫描线向下偏转,记录出一个向下的波形。这样,在神经冲动向右传导的过程中,就记录出了一个先升后降的双相动作电位。 负电极在前时,它首先记录到神经干表面由正变负的电位变化,经历了由正到负再到正的过程,因此记录出动作电位的上相。当在后的正电极记录到这种同样的电位变化过程时,显示相反的情况,记录出动作电位的下相。如果互换正、负电极的位置,则记录到先降后升的双相动作电位。 C.?? A点神经纤维多于B点(次要原因)。 (二)、神经干动作电位的引导及其传导 四实验步骤 (一)、制备蛙类坐骨神经-胫腓神经标本 通过观看录象让学生学习制作方法
一目的要求: 1.学习蛙类动物单毁髓与双毁髓的方法。 2.学习并掌握蛙类坐骨神经干标本的制备方法。 3.学习电生理学实验方法。 4.观察蟾蜍坐骨神经干复合动作电位的波形,了解其产生的基本原理。 二基本原理: 神经干在受到有效刺激后,可以产生动作电位,标志着神经发生兴奋。如果在神经干另一端引导传来的兴奋冲动,可以引导出双相的动作电位,如在两个引导电极之间将神经麻醉或损坏,则引导出的动作电位即为单相向动作电位。 神经细胞的动作位是以”全或无”方式发生的。坐骨神经干是由很多不同类型的神经纤维组成的,所以,神经干的动作电位是复合动作电位。复合动作电位的幅值在一定刺激强度下是随刺激强度的变化而变化的。 三动物与器材: 蟾蜍、常用手术器械(手术剪、手术镊、金冠剪、眼科剪、毁髓针和玻璃分针)、蛙板、固定针、不锈钢盘、污物缸、粗棉线、任氏液、计算机生理信号处理系统、神经屏蔽盒。 四方法步骤: 1.蟾蜍的单毁髓与双毁髓 一手握住蛙或蟾蜍(可用纱布包裹蟾蜍躯干部),背部向上。用拇指压住蛙或蟾蜍的背部,食指按压其头部前端,使头端向下低垂; 另一手持毁髄针,由两眼之间沿中线向后触划,当触及到两耳中间的凹陷处(此处与两眼的联机成等边三角形)时,持针手即感觉针尖下陷,此处即是枕骨大孔的位置。将毁髄针由凹陷处垂直刺入,即可进入枕骨大孔(图t-1)。然后将针尖向前刺入颅腔,在颅腔内搅动,以捣毁脑组织。如毁髄针确在颅腔内,实验者可感到针尖触及颅骨。此时的动物为单毁髓动物。再将毁髓针退至枕骨大孔,针尖转冋后方,与脊柱平行刺入椎管,以捣毁脊髓。彻底捣毁脊髓时,可看到动物的后肢突然蹬直,而后瘫痪如棉(图t-2),此时的动物为双毁髓动物。如动物仍表现肢肌肉紧张或活动自如,必须重新毁髓。操作过程中应注意使蟾蜍头部向外侧(不要挤压耳后腺),防止耳后腺分泌物射入实验者眼内(如被射入,则需立即手生理盐水冲洗眼睛)。 2.坐骨神经干标本制备 (1) 剥制后肢标本(图t-3) (2) 分离两后肢(图t-4)
静息电位与动作电位 一、静息电位(RP)的产生机制:在静息状态下,细胞膜对K+具有较高的通透性是形成静息电位的最主要因素。细胞膜内K+浓度约相当于细胞外液的30倍,K+将顺浓度梯度跨膜扩散,但扩散的同时也在细胞膜的两侧形成逐渐增大的电位差,且该电位差造成的驱动力与浓度差的驱动力的方向相反,阻止K+进一步跨膜扩散。当逐渐增大的电位差驱动力与逐渐减小的浓度差驱动力相等时,便达到了稳态。此时的膜电位处于K+的平衡电位(EK+=-90~-100mv),电位差的差值即平衡电位,平衡电位决定着离子的流量。当细胞外液中K+浓度增加(高钾)时,膜内外K+的浓度差减小,K+因浓度差外移的驱动力降低,K+外流减少。故达到稳态时,K+平衡电位的绝对值减小;反之亦然。而细胞膜对Na+亦有一定的通透性,扩散内流的Na+可以部分抵消由K+扩散外流所形成的膜内负电位。所以,EK+=-90~-100mv,而RP=-70~-90mv。可见,细胞外液Na+浓度对RP的影响不大。除了以上两个方面,还有钠泵的生电作用。钠泵使细胞内高钾、细胞外高钠。若钠泵受抑制,膜内外K+的浓度差减小,K+外流减少,K+平衡电位的绝对值减小,静息电位的绝对值也减小。综上所述,影响静息电位水平的因素:(1)细胞膜对K+和Na+的相对通透性;(2)细胞外液K+的浓度;(3)钠泵的活动。 二、动作电位(AP)的产生机制:在静息状态下,细胞膜外Na+浓度约为细胞内液的10倍余,Na+有向膜内扩散的趋势;并且静息时膜内存在着相当数量的负电位,吸引着Na+向膜内移动。但由于静息时细胞膜对Na+相对不通透,因此,Na+不能大量内流。 当刺激引起去极化达到阈电位,细胞膜上的电压门控Na+通道大量被激活,细胞膜对Na+的通透性突然增大,Na+大量内流,造成细胞膜的进一步去极化;而膜的进一步去极化,又将导致更多的Na+通道开放,有更多的Na+内流,引起细胞膜迅速、自动地去极化。 Na+的大量内流,以至膜内负电位因正电荷的增加而迅速消失。又因为细胞膜外Na+浓度约为细胞内液的10倍余,使得Na+内流在膜内负电位绝对值减小到零时仍可以继续,进而出现正电位,直至膜内正电位增大到足以对抗浓度差所引起的Na+内流,便达到了平衡电位(顶点),此时膜对Na+的净通量为零。但是膜内电位并不停留在正电位状态,很快Na+通道失活,膜对Na+变为相对不通透,而对K+的通透性增加。于是膜内K+在浓度差和电位差的驱动力下外流,使膜内电位由正电位又向负电位发展,以后再逐渐恢复到静息电位水平(动作电位的幅度由静息电位的绝对值和Na+的平衡电位值相加决定)。 当细胞外液Na+浓度降低时,膜内外Na+的浓度差减小,将导致去极化时Na+内流减少,Na+的平衡电位减小,动作电位峰值降低;反之亦然。
反射时测定和反射弧分析 神经干动作电位的测定 2013级生命科学3班张柏辉学号:20132501076 1.实验目的 1.观察蛙坐骨神经干动作电位的基本波形,并了解其产生的基本原理; 2.学习测定反射时的方法,了解反射弧的组成; 3.了解脊髓反射的功能特性。 2.实验原理 (一)反射时测定和反射弧分析 反射是指对某一刺激无意识的应答。反射活动的结构基础称为反射弧,包括感受器、传入神经、神经中枢、传出神经和效应器。从皮肤接受刺激至机体出现反应的时间称为反射时。反射时是反射通过反射弧所用的时间。反射弧的任何一部分缺损,原有的反射不再出现。中枢的兴奋和抑制同时存在又相互影响。在脊髓反射的中枢之间或高位脑和脊髓对低位脊髓反射中枢均存在抑制作用,这些抑制作用保证了机体活动的协调性。 (二)神经干动作电位的测定 神经干在受到有效刺激后可以产生复合动作电位,标志着兴奋的产生。如果在立体神经干的一端施加刺激,从另一端引导传来的兴奋冲动可以记录出双相动作电位,假如在引导的两个电极之间将神经干麻醉或损坏,阻断其兴奋传导能力,此时可以记录到单相动作电位。 3.实验对象与实验材料 (一)材料:虎纹蛙 (二)器具:手术剪、手术镊、手术刀、金冠剪、眼科剪、毁髓针、玻璃分针、木质蛙板、固定针、锌铜弓、瓷盘、污物缸、滴管、纱布、粗棉线、滤纸片、支架、蛙嘴夹、小烧杯、秒表、神经屏蔽盒、PowerLab、刺激线、USB线、电脑 (三)试剂:任氏液、2%普鲁卡因、0.5%及1%硫酸溶液 4.实验方法与步骤 (一)反射时与反射弧的测定 1. 屈反射
取一只虎纹蛙,只毁脑髓制成脊蛙(只毁脑),用蛙嘴夹夹住蛙下颌悬挂在支架上,右后肢最长趾浸入0.5%硫酸溶液中2~3mm(<10s),同时开始计时。当出现屈反射时立即停止计时,并用清水冲洗受刺激皮肤,纱布擦干,重复测屈反射时3次。同样方法测左后肢最长趾的屈反射时。 2.损毁感受器 用手术剪自后肢最长趾基部环切皮肤,后用手术镊剥净长趾上的皮肤,用0.5%硫酸溶液刺激去皮皮肤,并记录侧时结果。 3.对照没损毁感受器 改换同侧后趾有皮肤趾,将其浸入0.5%硫酸溶液中,测定反射时。 4.擦或抓反射 取一浸有0.5%硫酸溶液的滤纸片贴于虎纹蛙腹部,记录抓或擦反射的反射时。 5.麻醉神经 右侧坐骨神经滴加普鲁卡因液,加药同时开始计时,每2min重复步骤3,并记录加药时间。 屈反射消失后,重复步骤4,记录加药时间。 6.测左后肢最长趾屈反射时,并与步骤1比较。 7.毁坏脊髓,重复步骤7. (二)神经干动作电位的测定 1. 标本制备:坐骨神经干(双毁髓->剥制后肢->分离两后肢),分离坐骨神经到踝关节附近,将标本搭置在神经屏蔽盒各金属极上; 2.设置实验装置:连接神经屏蔽盒各接线; 3.设置CH3 BioAmp和Stimulator:打开PowerLab电源,打开Scope软件(或Chart5),设置通道3: Ch3 BioAmp:Range 5mV, Filter 20Hz,High Pass 10Hz (调零用DC档);设置Stimulator:单刺激Delay 120ms ,波宽Duration:10mS,振幅Ampt:100mV;设置Overlay on Top 点右下角Start ,即可看到刺激输出后得到的动作电位波形图。每点一次START,记录号增加在图下方,调节单刺激持续时间Duration和振幅大小,以及调节放大器HighPass等参数,均对实验结果有影响。得到双相电位后,以普鲁卡因液或棉线结扎法损伤神经,调参数至振幅Amp5.000mV,,观察单相电位。