
真核细胞常见表达载体 真核细胞, 表达载体 1、pCMVp-NEO-BAN载体 特点:该真核细胞表达载体分子量为6600碱基对,主要由CMVp启动子、兔β-球蛋白基因内含子、聚腺嘌呤、氨青霉素抗性基因和抗neo基因以及pBR322骨架构成,在大多数真核细胞内都能高水平稳定地表达外源目的基因。更重要的是,由于该真核细胞表达载体中抗neo 基因存在,转染细胞后,用G418筛选,可建立稳定的、高表达目的基因的细胞株。 插入外源基因的克隆位点包括Sal1、BamH1和EcoR1位点。注意在此载体中有二个EcoR1位点存在。 2、pEGFP, 增强型绦色荧光蛋白表达载体(Enhanced Fluorecent Protein Vector) 特点: pEGFP表达载体中含有绿色荧光蛋白,在PCMV启动子驱动下,在真核细胞中高水平表达。载体骨架中的SV40origin使该载体在任何表达SV40 T抗原的真核细胞内进行复制。Neo抗性盒由SV40早期启动子、Tn5的neomycin/kanamycin抗性基因以及HSV-TK基因的聚腺嘌呤信号组成,能应用G418筛选稳定转染的真核细胞株。此外,载体中的pUC origin 能保证该载体在大肠杆菌中的复制,而位于此表达盒上游的细菌启动子能驱动kanamycin抗性基因在大肠杆菌中的表达。 用途: 该表达载体EGFP上游有Nde1、Eco47111和Age1克隆位点,将外源基因扦入这些位点,将合成外源基因和EGFP的融合基因。借此可确定外源基因在细胞内的表达和/或组织中的定位。 亦可用于检测克隆的启动子活性(取代CMV启动子,Acet1-Nhe1)。 3、pEGFT-Actin, 增强型绿色荧光蛋白/人肌动蛋白表达载体 特点:pEGFP-Actin表达载体中含有绿色荧光蛋白和人胞浆β-肌动蛋白基因,在PCMV启动子驱动下,在真核细胞中高水平表达。载体骨架中的SV40origin使该载体在任何表达SV40 T 抗原的真核细胞内进行复制。Neo抗性盒由SV40早期启动子、Tn5的neomycin/kanamycin 抗性基因以及HSV-TK基因的聚腺嘌呤信号组成,能应用G418筛选稳定转染的真核细胞株。此外,载体中的pUC origin 能保证该载体在大肠杆菌中的复制,而位于此表达盒上游的细菌启动子能驱动kanamycin抗性基因在大肠杆菌中的表达。 用途: pEGFP-Actin载体在真核细胞表达EGFP-Actin融合蛋白,该蛋白能整合到胞内正在生的肌动蛋白,因而在活细胞和固定细胞中观察到细胞内含肌动蛋白的亚细胞结构。 4、pSV2表达载体 特点:该表达质粒是以病责SV40启动子驱动在真核细胞目的基因进行表达的,克隆位点为Hind111。SV40启动子具有组织/细胞的选择特异性。此载体不含neo基因,故不能用来筛选、建立稳定的表达细胞株。 5、CMV4 表达载体 特点:该真核细胞表达载体由CMV启动子驱动,多克隆区域酶切位点选择性较多。含有氨苄青霉素抗性基因和生长基因片段以及SV40复制原点和fl单链复制原点。但值得注意的是,该表达载体不含有neo基因,转染細胞后不能用G418筛选稳定的表达细胞株。 其他常用克隆Vector: pBluscript II KS DNA 15 ug pUC18 DNA 25 ug pUC19 DNA 25 ug 说明: pBluescript II kS、pUC18 &Puc19载体适合于DNA片段的克隆、DNA测序和对外源基因进行表达等。这些载体由于在lacZ基因中含有多克隆位点,当外源DNA片段扦入,转化lacZ基因缺乏细胞,并在含有IPTG和X-gal的培养基上培养时,含有外源DNA载体的细胞将
原核蛋白表达常见问题解析 1、为什么目的蛋白总是以包涵体的形式出现? 在原核蛋白表达纯化中目的蛋白经常发生错误的折叠,并聚集成为 包涵体。经过诱导,目的蛋白通常可达细胞总蛋白的50%以上。虽然有一定比例的蛋白以可溶的单体形式存在,而多达95%(甚至更多)的蛋白则在包涵体中。实验过程中,可以采取降低诱导温度,例如25–30°C,或降低IPTG浓度(0.01–0.1mM)并延长诱导时间,还有采用特别的 培养基等方法获得更多的可溶蛋白。 2、跨膜蛋白为什么很难表达? 跨膜蛋白的表达成功率相对较低是一个实验结果,究其原理,目前 众说纷纭很多种理论。以我们浅薄的理解层面来看,主要有以下几个 原因: 跨膜蛋白一般都是强疏水性的氨基酸分子和亲水性的分子跳跃式 的连接,形成的亲水疏水的一个最简单的跨膜化学结构,这种结构与 信号肽结构相似,对于原核细胞来说,简单的细胞器很难像真核细胞 一样完成信号肽识别及切除、引导内质网、高尔基体重新包装及分泌 这一复杂过程,有些蛋白是多次跨膜,对于原核细胞来说几乎是不可 能完成的任务。 另外,对于疏水性的片段,在原核细胞中极易形成包涵体,疏水 性多肽会抑制翻译过程,甚至与原核膜结构融合形成毒性,出于生物 自我保护的本能,所有的细胞器都会停止合成蛋白的过程。 3、如何选择蛋白表达宿主菌?
4、我们有哪些原核蛋白纯化方式?如何选择不同的纯化方式? 答:我们公司的蛋白纯化方法大致分为亲和纯化、离子交换、切胶 回收三类。 1、常规情况下,一般携带融合标签(His标签,GST标签,sumo标签,Fc标签),我们可以通过Ni柱、GST柱、Protein A等进行亲和纯化 获得融合蛋白,用亲和纯化的方法一般可以获得85%以上纯度的蛋白, 亲和纯化的方便快捷。 2、如果需要目的蛋白不含有任何标签,怎么选择纯化方式?。 (1)可表达融合蛋白,用蛋白工具酶切割融合蛋白,再进行纯化除去 工具酶。此方法能快速得到蛋白。 (2)可表达不含标签的蛋白,进行离子、分子筛、疏水等纯化,通过AKATA纯化设备获得蛋白。 3、如果需要获得蛋白作为抗原,可以直接通过切胶回收的方式,此方 法获得蛋白纯度较高,进行免疫动物后得到的抗体进行WB反应,灵敏 度较高。 5、表达得到的蛋白是有活性的么? 答:需要让蛋白有活性的条件很复杂,合适的缓冲液体系、盐浓度、蛋白的折叠状态甚至检测活性的方法的细微差别都可能导致活性 的强弱有无,一般情况下,上清表达的蛋白要比包涵体经过变复性纯 化后得到的蛋白活性要好,我们尽量从上清中获得蛋白,期许蛋白形 成的折叠最接近活性状态,这也是我们擅长的。但是在实际实验条件下,我们无法承诺表达纯化的蛋白一定具有客户期望的生理活性。
原核表达遇到瓶颈怎么 办 TPMK standardization office【 TPMK5AB- TPMK08- TPMK2C- TPMK18】
我想表达遇到的第一个瓶颈估计就是为什么我的外源片段插到载体里面,PCR鉴定没问题,双酶切也OK,可是就是不见表达。一般我们是如何判断没有表达呢?大多都是首先进行SDS-PAGE。在跑胶的时候一定要设对照,比较严谨的电泳对照,应该是:Marker,标准品阳性对照(如果有,且打算做Western的话),以及空白载体(诱导)和重组载体(不诱导)2个阴性对照,再加上诱导不同时间的表达结果。Marker用于判断条带大小,标准品用于判断Western体系包括抗体显色剂和操作的可靠性,以及精确判断大小;空白载体(诱导)负对照有助于判断在非诱导条件下的本底表达;而重组载体(不诱导)负对照则有助于判断诱导的效果以及排除诱导剂对宿主菌潜在的干扰,或者是区分细菌内源和外源表达产物——偷懒可不是好习惯,会影响结果判断。注意,接种表达和接种做感受态都类似,一定要用挑单克隆、加足量抗生素、过夜培养的新鲜菌液转种最好,在筛选压力下生长旺盛的种子液远比经过4℃保存的菌液要好得多,也许是因为保存时间长会导致抗生素失效部分细菌丢失质粒或者其他变化。反正就是一种经验做法。 跑SDS-PAGE的话可以用考马斯亮兰染,它灵敏度在100ng左右;但是不能跟着做Western了。银染的灵敏度在0.1~1 ng;有钱还可以用Sypro Red,灵敏度高还可以继续做Western。WesternBlot是蛋白质定性和半定量的最通用的技术,具体细节可参考生物通WesternBlot技术专题,我们有详细介绍的。 在做过Western Blot仍没有检测到表达带,那么就要开始进行下一步的分析了。首先,看看载体的多克隆位点和片断插入的序列,是否有因为酶切连接而意外引入了转录终止信号。有时载体几经多个实验人员的周转,反复插入片断,或者是粘端
载体与表达系统 0001--pIRES --BD Co--真核表达--yshu3507 0002--pECFP-C1--BD Co--真核表达--yshu3507 0003--pShuttle--/--/--victoh 0004--pSBR322--/--/--chujun_hust 0005--pcDNA3.1(+)/CAT--invitrogen--真核表达--wangjun2002274 0006--pQEx--Qiagen--原核表达--wangjun2002274 0007--pIVEX2.3--Roche--体外转录翻译--wangjun2002274 0008--pIRES-EGFP--/--/--luoqingli2003 0009--pET-28a(+)--Novagen--原核表达--wangjun2002274 0010--pSUPER Vector--oligoengine--siRNA--zyh961171 0011--pET-32a(+)--novagen--原核表达--Crazyvirus 0012--pBK-CMV--/--原核表达、真核表达--zrzhong6519 0013--pcDNA3.1/Zeo (+)--invitrogen--原核表达--Crazyvirus 0014--pcDNA3--invitrogen--真核表达--kras 0015--pfastbac1--invitrogene--昆虫表达--cox2wj 0016--pEGFP-C3--clontech--真核表达--smilely 0017--pSecTag2--invitrogen--真核表达--kenmed 0018--PYX212--/--真核表达--Gmail 0019--pET20b--Novagen--原核表达--Gmail 0020--pEGFP-1--BD BIO--转录调控--wyh 0021--pEGFP/U6 --/--siRNA--songbinn
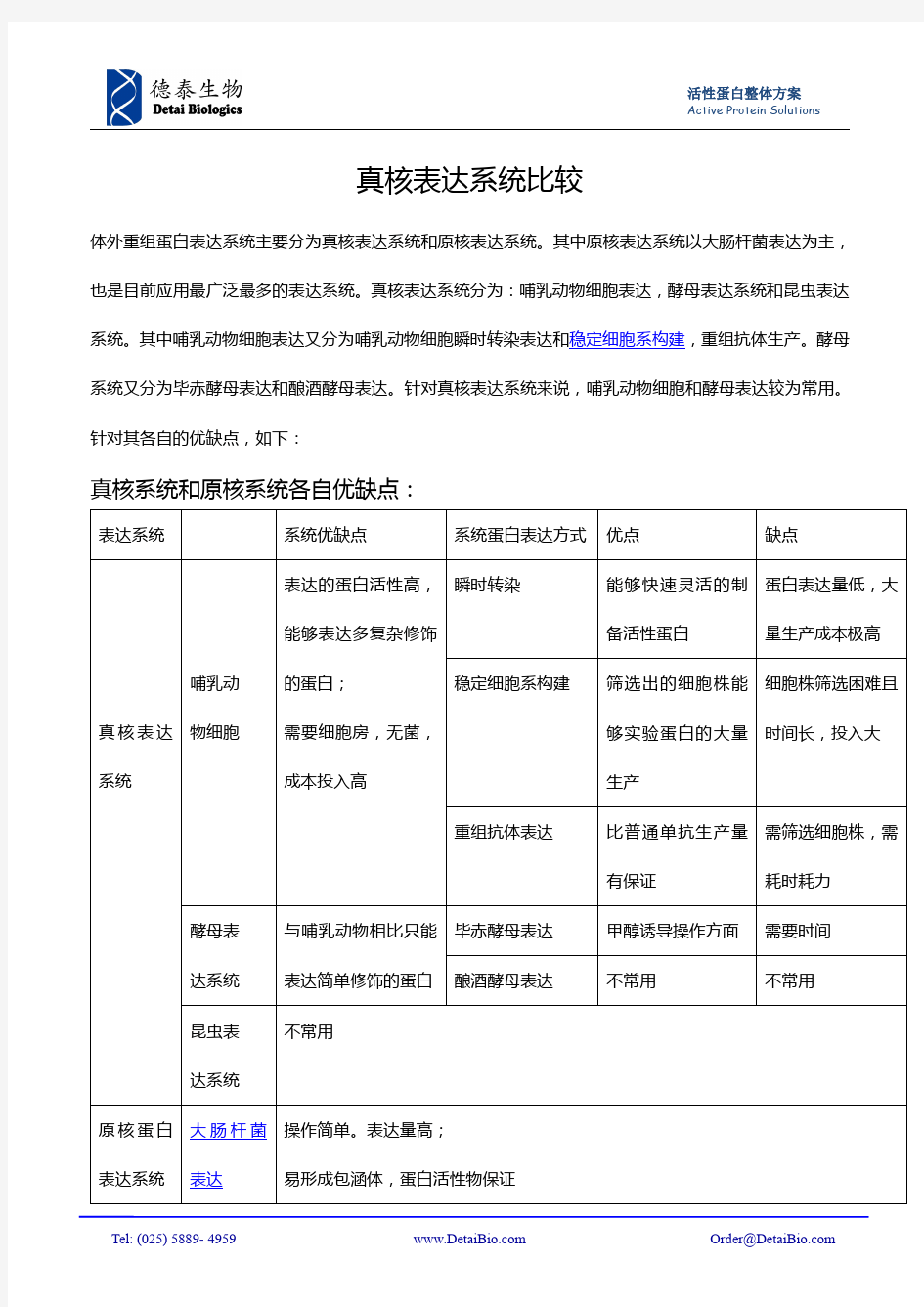
真核细胞表达系统的类型与常用真核细胞表达载体 标签:真核细胞酵母表达系统细胞表达载体真核表达系统昆虫表达系统动物表达系统 摘要: 原核表达系统是常被用来研究基因功能的成熟系统,由于原核表达系统具有包涵体蛋白不易纯化、蛋白修饰不完整等缺陷,人们也开始利用真核细胞表达系统来研究基因。 原核表达系统是常被用来研究基因功能的成熟系统,由于原核表达系统具有包涵体蛋白不易纯化、蛋白修饰不完整等缺陷,人们也开始利用真核细胞表达系统来研究基因。 自上世纪70年代基因工程技术诞生以来,基因表达技术已渗透到生命科学研究的各个领域。并随着人类基因组计划实施的进行,在技术方法上得到了很大发展,时至今日已取得令人瞩目的成就。随着人类基因组计划的完成,越来越多的基因被发现,其中多数基因功能不明。利用表达系统在哺乳动物细胞内表达目的基因是研究基因功能及其相互作用的重要手段。 在各种表达系统中,最早被采用进行研究的是原核表达系统,这也是目前掌握最为成熟的表达系统。该项技术的主要方法是将已克隆入目的基因DNA段的载体(一般为质粒)转化细菌(通常选用的是大肠杆菌),通过iptg诱导并最终纯化获得所需的目的蛋白。其优点在于能够在较短时间内获得基因表达产物,而且所需的成本相对比较低廉。但与此同时原核表达系统还存在许多难以克服的缺点:如通常使用的表达系统无法对表达时间及表达水平进行调控,有些基因的持续表达可能会对宿主细胞产生毒害作用,过量表达可能导致非生理反应,目的蛋白常以包涵体形式表达,导致产物纯化困难;而且原核表达系统翻译后加工修饰体系不完善,表达产物的生物活性较低。 为克服上述不足,许多学者将原核基因调控系统引入真核基因调控领域,其优点是: ①根据原核生物蛋白与靶DNA间作用的高度特异性设计,而靶DNA与真核基因调控序列基本无同源性,故不存在基因的非特异性激活或抑制; ②能诱导基因高效表达,可达105倍,为其他系统所不及; ③能严格调控基因表达,即不仅可控制基因表达的“开关”,还可人为地调控基因表达量。 因此,利用真核表达系统来表达目的蛋白越来越受到重视。目前,基因工程研究中常用的真核表达系统有酵母表达系统、昆虫细胞表达系统和哺乳动物细胞表达系统。 1.酵母表达系统 最早应用于基因工程的酵母是酿酒酵母,后来人们又相继开发了裂殖酵母、克鲁维酸酵母、甲醇酵母等,其中,甲醇酵母表达系统是目前应用最广泛的酵母表达系统。目前甲醇酵母主要有H Polymorpha,Candida Bodini,Pichia Pastris3种。以Pichia Pastoris应用最多。
pET 原核表达 pET 载体中,目标基因克隆到T7 噬菌体强转录和翻译信号控制之下,并通过在宿主细胞提供T7 RNA 聚合酶来诱导表达。Novagen 的pET 系统不断扩大,提供了用于表达的新技术和选择,目前共包括36 种载体类型、15 种不同宿主菌和设计用于有效检测和纯化目标蛋白的许多其它相关产品。 优点 ·是原核蛋白表达引用最多的系统 ·在任何大肠杆菌表达系统中,基础表达水平最低 ·真正的调节表达水平的“变阻器”控制 ·提供各种不同融合标签和表达系统配置 ·可溶性蛋白生产、二硫键形成、蛋白外运和多肽生产等专用载体和宿主菌 ·许多载体以LIC 载体试剂盒提供,用于迅速定向克隆PCR 产物 ·许多宿主菌株以感受态细胞形式提供,可立即用于转化 阳性pFORCE TM 克隆系统具有高效克隆PCR 产物、阳性选择重组体和高水平表达目标蛋白等特点。 pET 系统概述 pET 系统是在大肠杆菌中克隆和表达重组蛋白的最强大系统。根据最初由Studier 等开发的T7 启动子驱动系统,Novagen 的pET 系统已用于表达成千上万种不同蛋白。 控制基础表达水平 pET 系统提供6 种载体- 宿主菌组合,能够调节基础表达水平以优化目标基因的表达。没有单一策略或条件适用于所有目标蛋白,所以进行优化选择是必要的。 宿主菌株 质粒在非表达宿主菌中构建完成后,通常转化到一个带有T7 RNA 聚合酶基因的宿主菌(λDE3 溶原菌)中表达目标蛋白。在λDE3 溶原菌中,T7 RNA 聚合酶基因由lacUV5 启动子控制。未诱导时便有一定程度转录,因此适合于表达其产物对宿主细胞生长无毒害作用的一些基因。而宿主菌带有pLysS 和pLyE 时调控会更严紧。pLys 质粒编码T7 溶菌酶,它是T7 RNA 聚合酶的天然抑制物,因此可降低其在未诱导细胞中转录目标基因的能力。pLysS 宿主菌产生低量T7 溶菌酶,而pLysE 宿主菌产生更多酶,因此是最严紧控制的λDE3 溶原菌。 有11 种不同DE3 溶原化宿主菌。使用最广泛的为BL21 及其衍生菌株,它的优点在于缺失lon 和ompT 蛋白酶。B834 菌株为甲硫氨酸营养缺陷型,因此可用35 S- 甲硫氨酸和硒代甲硫氨酸对目标蛋白进行高特异活性标记。BLR 为recA - 衍生菌株,改善了质粒单体产量,有助于稳定含有重复序列的目标质粒。两个硫氧还蛋白还原酶( trxB ) 突变菌株(AD494,BL21 trxB ) ,有利于大肠杆菌胞浆中二硫键形成。Origami TM 和OrigamiB 菌株为trxB/gor 双突变,这两个酶是主要还原途径的关键酶。Origami 和OrigamiB 宿主菌的主要优点是能形成正确折迭的含有二硫键的蛋白。新的Rosetta TM 菌株补充了四种大肠杆菌稀有密码子的tRNA ,改善了由于密码子使用频率不同而引起的一些真核蛋白低表达。其它菌株背景包括K-12 菌株HMS174 和NovaBlue ,象BLR 一样为recA - 。这些菌株可稳定表达其产物可能导致DE3 噬菌体丢失的某些目标基因。由于存在F 附加体编码
原核、真核表达载体构建 真核表达载体和原核表达载体的区别:主要是因为原核和真核表达系统所需的表达元件不同。 比如说启动子,终止子在两种表达系统中是不一样的。 带有真核表达元件的是真核载体,能在真核生物内表达; 带有原核表达元件的是原核载体,能在原核生物内表达。两者都具有的为穿梭载体。 ㈠原核表达载体指:能携带插入的外源核酸序列进入原核细胞中进行复制的载体。 原核表达载体调控原件 1.启动子 启动子是DNA链上一段能与RNA聚合酶结合并起始RNA合成的序列,它是基因表达不可缺少的重要调控序列。没有启动子,基因就不能转录。由于细菌RNA聚合酶不能识别真核基因的启动子,因此原核表达载体所用的启动子必须是原核启动子。原核启动子是由两段彼此分开且又高度保守的核苷酸序列组成,对mRNA的合成极为重要。在转录起始点上游5~10 bp处,有一段由6~8个碱基组成,富含A和T的区域,称为Pribnow 盒,又名TATA 盒或-10区。来源不同的启动子,Pribnow 盒的碱基顺序稍有变化。在距转录起始位点上游35 bp 处,有一段由10 bp组成的区域,称为-35区。转录时大肠杆菌RNA聚合酶识别并结合启动子。-35区与RNA聚合酶s亚基结合,-10区与RNA聚合酶的核心酶结合,在转录起始位点附近DNA被解旋形成单链,RNA聚合酶使第一和第二核苷酸形成磷酸二酯键,以后在RNA聚合酶作用下向前推进,形成新生的RNA 链。原核表达系统中通常使用的可调控的启动子有Lac(乳糖启动子)、Trp(色氨酸启动子)、Tac(乳糖和色氨酸的杂合启动子) 、lPL (l噬菌体的左向启动子)、T7噬菌体启动子等。 2. SD序列 1974年Shine和Dalgarno首先发现,在mRNA上有核糖体的结合位点,它们是起始密码子AUG和一段位于AUG上游3~10 bp处的由3~9 bp组成的序列。这段序列富含嘌呤核苷酸,刚好与16S rRNA 3¢末端的富含嘧啶的序列互补,是核糖体RNA的识别与结合位点。以后将此序列命名为Shine-Dalgarno序列,简称SD序列。它与起始密码子AUG之间的距离是影响mRNA转录、翻译成蛋白的重要因素之一,某些蛋白质与SD序列结合也会影响mRNA与核糖体的结合,从而影响蛋白质的翻译。另外,真核基因的第二个密码子必须紧接在ATG 之后,才能产生一个完整的蛋白质。 3.终止子 在一个基因的3¢末端或是一个操纵子的3'末端往往有特定的核苷酸序列,且具有终止转录功能,这一序列称之为转录终止子,简称终止子(terminator)。转录终止过程包括:RNA聚合酶停在DNA模板上不再前进,RNA的延伸也停止在终止信号上,完成转录的RNA从RNA聚合酶上释放出来。对RNA聚合酶起
标签:真核细胞酵母表达系统细胞表达载体真核表达系统昆虫表达系统动物表达系统 摘要 : 原核表达系统是常被用来研究基因功能的成熟系统,由于原核表达系统具有包涵体蛋白不易纯化、蛋白修饰不完整等缺陷,人们也开始利用真核细胞表达系统来研究基因。 原核表达系统是常被用来研究基因功能的成熟系统,由于原核表达系统具有包涵体蛋白不易纯化、蛋白修饰不完整等缺陷,人们也开始利用真核细胞表达系统来研究基因。 自上世纪70年代基因工程技术诞生以来,基因表达技术已渗透到生命科学研究的各个领域。并随着人类基因组计划实施的进行,在技术方法上得到了很大发展,时至今日已取得令人瞩目的成就。随着人类基因组计划的完成,越来越多的基因被发现,其中多数基因功能不明。利用表达系统在哺乳动物细胞内表达目的基因是研究基因功能及其相互作用的重要手段。 在各种表达系统中,最早被采用进行研究的是原核表达系统,这也是目前掌握最为成熟的表达系统。该项技术的主要方法是将已克隆入目的基因DNA段的载体(一般为质粒)转化细菌(通常选用的是大肠杆菌),通过iptg诱导并最终纯化获得所需的目的蛋白。其优点在于能够在较短时间内获得基因表达产物,而且所需的成本相对比较低廉。但与此同时原核表达系统还存在许多难以克服的缺点:如通常使用的表达系统无法对表达时间及表达水平进行调控,有些基因的持续表达可能会对宿主细胞产生毒害作用,过量表达可能导致非生理反应,目的蛋白常以包涵体形式表达,导致产物纯化困难;而且原核表达系统翻译后加工修饰体系不完善,表达产物的生物活性较低。 为克服上述不足,许多学者将原核基因调控系统引入真核基因调控领域,其优点是: ①根据原核生物蛋白与靶DNA间作用的高度特异性设计,而靶DNA与真核基因调控序列基本无同源性,故不存在基因的非特异性激活或抑制; ②能诱导基因高效表达,可达105倍,为其他系统所不及; ③能严格调控基因表达,即不仅可控制基因表达的“开关”,还可人为地调控基因表达量。 因此,利用真核表达系统来表达目的蛋白越来越受到重视。目前,基因工程研究中常用的真核表达系统有酵母表达系统、昆虫细胞表达系统和哺乳动物细胞表达系统。 1.酵母表达系统 最早应用于基因工程的酵母是酿酒酵母,后来人们又相继开发了裂殖酵母、克鲁维酸酵母、甲醇酵母等,其中,甲醇酵母表达系统是目前应用最广泛的酵母表达系统。目前甲醇酵母主要有H Polymorpha,Candida Bodini,Pichia Pastris3种。以Pichia Pastoris 应用最多。
真核细胞常见表达载体 1、pCMVp-NEO-BAN载体 特点: 该真核细胞表达载体分子量为6600碱基对,主要由CMVp启动子、兔β-球蛋白基因内含子、聚腺嘌呤、氨青霉素抗性基因和抗neo基因以及pBR322骨架构成,在大多数真核细胞内都能高水平稳定地表达外源目的基因。更重要的是,由于该真核细胞表达载体中抗neo基因存在,转染细胞后,用G418筛选,可建立稳定的、高表达目的基因的细胞株。 插入外源基因的克隆位点包括Sal1、BamH1和EcoR1位点。注意在此载体中有二个EcoR1位点存在。 2、pEGFP, 增强型绦色荧光蛋白表达载体(Enhanced Fluorecent Protein Vector) 特点: pEGFP表达载体中含有绿色荧光蛋白,在PCMV启动子驱动下,在真核细胞中高水平表达。载体骨架中的SV40 origin使该载体在任何表达SV40 T 抗原的真核细胞内进行复制。Neo抗性盒由SV40早期启动子、Tn5的neomycin/kanamycin抗性基因以及HSV-TK基因的聚腺嘌呤信号组成,能应用G418筛选稳定转染的真核细胞株。此外,载体中的pUC origin 能保证该载体在大肠杆菌中的复制,而位于此表达盒上游的细菌启动子能驱动kanamycin抗性基因在大肠杆菌中的表达。 用途: 该表达载体EGFP上游有Nde1、Eco47111和Age1克隆位点,将外源基因扦入这些位点,将合成外源基因和EGFP的融合基因。借此可确定外源基因在细胞内的表达和/或组织中的定位。 亦可用于检测克隆的启动子活性(取代CMV启动子,Acet1-Nhe1)。 3、pEGFT-Actin, 增强型绿色荧光蛋白/人肌动蛋白表达载体 特点: pEGFP-Actin表达载体中含有绿色荧光蛋白和人胞浆β-肌动蛋白基因,在PCMV启动子驱动下,在真核细胞中高水平表达。载体骨架中的SV40 origin使该载体在任何表达SV40 T 抗原的真核细胞内进行复制。Neo抗性盒由SV40早期启动子、Tn5的neomycin/kanamycin抗性基因以及HSV-TK基因的聚腺嘌呤信号组成,能应用G418筛选稳定转染的真核细胞株。此外,载体中的pUC origin 能保证该载体在大肠杆菌中的复制,而位于此表达盒上游的细菌启动子能驱动kanamycin抗性基因在大肠杆菌中的表达。 用途: pEGFP-Actin载体在真核细胞表达EGFP-Actin融合蛋白,该蛋白能整合到胞内正在生的肌动蛋白,因而在活细胞和固定细胞中观察到细胞内含肌动蛋白的亚细胞结构。 4、pSV2表达载体 特点:该表达质粒是以病责SV40启动子驱动在真核细胞目的基因进行表达的,克隆位点为Hind111。SV40启动子具有组织/细胞的选择特异性。此载体不含neo基因,故不能用来筛选、建立稳定的表达细胞株。5、CMV4 表达载体 特点:该真核细胞表达载体由CMV启动子驱动,多克隆区域酶切位点选择性较多。含有氨苄青霉素抗性基因和生长基因片段以及SV40复制原点和fl单链复制原点。但值得注意的是,该表达载体不含有neo基因,转染細胞后不能用G418筛选稳定的表达细胞株。 其他常用克隆Vector: pBluscript II KS DNA 15 ug pUC18 DNA 25 ug pUC19 DNA 25 ug
Chi l 原核表达基本试验步骤 将克隆化基因插入合适载体后导入大肠杆菌用于表达大量蛋白质的方法一般称为原核表达。这种方法在蛋白纯化、定位及功能分析等方面都有应用。大肠杆菌用于表达重组蛋白有以下特点:易于生长和控制;用于细菌培养的材料不及哺乳动物细胞系统的材料昂贵;有各种各样的大肠杆菌菌株及与之匹配的具各种特性的质粒可供选择。但是,在大肠杆菌中表达的蛋白由于缺少修饰和糖基化、磷酸化等翻译后加工,常形成包涵体而影响表达蛋白的生物学活性及构象。 表达载体在基因工程中具有十分重要的作用,原核表达载体通常为质粒,典型的表达载体应具有以下几种元件: (1)选择标志的编码序列; (2)可控转录的启动子; (3)转录调控序列(转录终止子,核糖体结合位点); (4)一个多限制酶切位点接头; (5)宿主体内自主复制的序列。 原核表达一般程序如下:获得目的基因-准备表达载体-将目的基因插入表达载体中(测序验证)-转化表达宿主菌-诱导靶蛋白的表达-表达蛋白的分析-扩增、纯化、进一步检测,其中包括: 一、试剂准备 (1)LB培养基。 (2)1M IPTG(异丙基硫代-β-D-半乳糖苷):2.38g IPTG溶于10ml ddH2O
中,0.22μm滤膜抽滤,-20℃保存。 CCY的IPTG是1M的,用时进行1000倍稀释。 二、操作步骤 (一)获得目的基因 1、通过PCR方法:以含目的基因的克隆质粒为模板,按基因序列设计一对引物(在上游和下游引物分别引入不同的酶切位点),PCR循环获得所需基因片段。 2、通过RT-PCR方法:用TRIzol法从细胞或组织中提取总RNA,以mRNA 为模板,逆转录形成cDNA第一链,以逆转录产物为模板进行PCR循环获得产物。 (二)构建重组表达载体 1、载体酶切:将表达质粒用限制性内切酶(同引物的酶切位点)进行双酶切,酶切产物行琼脂糖电泳后,用胶回收Kit或冻融法回收载体大片段。 2、PCR产物双酶切后回收,在T4DNA连接酶作用下连接入载体。我们用Soultion I连接。 (三)获得含重组表达质粒的表达菌种 1、将连接产物转化大肠杆菌BL21,根据重组载体的标志(抗Amp或蓝白斑)作筛选,挑取单斑,碱裂解法小量抽提质粒,双酶切初步鉴定。 2、测序验证目的基因的插入方向及阅读框架均正确,进入下步操作。否则应筛选更多克隆,重复亚克隆或亚克隆至不同酶切位点。 3、以此重组质粒DNA转化表达宿主菌的感受态细胞。
原核生物和真核生物基因表达调控特点的比较1.相同点:转录起始是基因表达调控的关键环节2.不同点:A.原核基因的表达调控主要包括转录和翻译水平 真核基因的表达调 控主要包括染色质活化、转录、转录后加工、翻译、翻译后加工多个层次B.原核基因表达调控主要为负调控,真核主要为正调控C.原核转录不需要转录因子,RNA聚合酶直接结合启 动子,由sita因子决定基因表的的特异性 真核基因转录起始需要基础特异两类转录因子 依赖DNA-蛋白质、蛋白质-蛋白质相互作用 调控转录激活D.原核基因表达调控主要采用操纵子模型 转录出多顺反子RNA 实现协调调节 真核基因转录产物为单顺反子RNA 功能相关蛋白的协调表达机制更为复杂。真核生物基因表达调控的环节主要在转录水平 其次是翻译水平。原核生物基因以操纵子的形式存在。转录水平调控涉及到启动子、sita因子 与RNA聚合酶结合 、阻遏蛋白 负调控 、正调控蛋白、倒位蛋白、RNA聚合酶抑制物、衰减子等。翻译水平的调控涉及SD序列、mRNA的稳定性 不稳定(5’端和3’端的发夹结构可保护不被酶水解mRNA的5’端与核糖体结合 可明显提高稳定性)、翻译产物及小分子RNA的调控作用。真核生物基因表达的调控环节较多 在DNA水平上可以通过染色体 丢失、基因扩增、基因重排、DNA甲基化、染色体结构改变影响基因表达。在转录水平主要通过反式作用因子调控转录因子与TATA盒的结合、RNA聚合酶与转录因子-DNA复合物的结合及转录起始复合物的形成。在转录后水平主要通过RNA修饰、剪接及mRNA运输的控制来影响基因表达。在翻译水平有影响起始翻译的阻遏蛋白、5’AUG、5’端非编码区长度、mRNA 的稳定性调节及小分子RNA。真核基因调控中最重要的环节是基因转录 真核生物基因表达需要转录因子、启动子、沉默子和增强子。葡萄糖存在 乳糖不存在 此时无诱导剂
原核表达步骤
Chi l 原核表达基本试验步骤 将克隆化基因插入合适载体后导入大肠杆菌用于表达大量蛋白质的方法一般称为原核表达。这种方法在蛋白纯化、定位及功能分析等方面都有应用。大肠杆菌用于表达重组蛋白有以下特点:易于生长和控制;用于细菌培养的材料不及哺乳动物细胞系统的材料昂贵;有各种各样的大肠杆菌菌株及与之匹配的具各种特性的质粒可供选择。但是,在大肠杆菌中表达的蛋白由于缺少修饰和糖基化、磷酸化等翻译后加工,常形成包涵体而影响表达蛋白的生物学活性及构象。 表达载体在基因工程中具有十分重要的作用,原核表达载体通常为质粒,典型的表达载体应具有以下几种元件: (1)选择标志的编码序列; (2)可控转录的启动子; (3)转录调控序列(转录终止子,核糖体结合位点); (4)一个多限制酶切位点接头; (5)宿主体内自主复制的序列。 原核表达一般程序如下:获得目的基因-准备表达载体-将目的基因插入表达载体中(测序验证)-转化表达宿主菌-诱导靶蛋白的表达-表达蛋白的分析-扩增、纯化、进一步检测,其中包括: 一、试剂准备 (1)LB培养基。 (2)1M IPTG(异丙基硫代-β-D-半乳糖苷):2.38g IPTG溶于10ml ddH2O
中,0.22μm滤膜抽滤,-20℃保存。 CCY的IPTG是1M的,用时进行1000倍稀释。 二、操作步骤 (一)获得目的基因 1、通过PCR方法:以含目的基因的克隆质粒为模板,按基因序列设计一对引物(在上游和下游引物分别引入不同的酶切位点),PCR循环获得所需基因片段。 2、通过RT-PCR方法:用TRIzol法从细胞或组织中提取总RNA,以mRNA 为模板,逆转录形成cDNA第一链,以逆转录产物为模板进行PCR循环获得产物。 (二)构建重组表达载体 1、载体酶切:将表达质粒用限制性内切酶(同引物的酶切位点)进行双酶切,酶切产物行琼脂糖电泳后,用胶回收Kit或冻融法回收载体大片段。 2、PCR产物双酶切后回收,在T4DNA连接酶作用下连接入载体。我们用Soultion I连接。 (三)获得含重组表达质粒的表达菌种 1、将连接产物转化大肠杆菌BL21,根据重组载体的标志(抗Amp或蓝白斑)作筛选,挑取单斑,碱裂解法小量抽提质粒,双酶切初步鉴定。 2、测序验证目的基因的插入方向及阅读框架均正确,进入下步操作。否则应筛选更多克隆,重复亚克隆或亚克隆至不同酶切位点。 3、以此重组质粒DNA转化表达宿主菌的感受态细胞。
[Merck推荐]原核表达秘笈之宿主菌株选择指南 在原核蛋白表达过程中,选择构建一个合适原核表达体系需要综合考虑3大因素:表达载体、宿主菌株、表达诱导条件,以获得最满意的表达效果。 事实上,在平时的实验中,最容易被忽视的就是宿主菌的选择——多数人会直接选择自己实验室曾经用过的表达菌株,或者是载体配套的菌株,而不去追究原因——即使表达结果不佳,大多在表达条件和载体上找原因,也不会考究菌株的选择是否适合。 作为原核表达的宿主,对外源基因的表达会产生一定的影响,是勿庸置疑的。每一个宿主细胞都像一个微观的小工厂,按照细胞固有的程序完成“你给它们安排的生产任务”——因为很难亲眼观察微观世界中表达是如何进行的,当出现问题时,我们需要经验判断问题所在。宿主细胞对原核表达可能会产生哪些影响呢? 知其然还要知其所以然。比如,菌株内源的蛋白酶过多,可能会造成外源表达产物的不稳定,所以一些蛋白酶缺陷型菌株往往成为理想的起始表达菌株。堪称经典的BL21系列就是lon和ompT蛋白酶缺陷型,也是我们非常熟悉的表达菌株。大名鼎鼎的BL21(DE3)融源菌则是添加T7聚合酶基因,为T7表达系统而设计。 真核细胞偏爱的密码子和原核系统有不同,因此,在用原核系统表达真核基因的时候,真核基因中的一些密码子对于原核细胞来说可能是稀有密码子,从而导致表达效率和表达水平很低。改造基因是比较麻烦的做法,Rosetta 2系列就是更好的选择——这种携带pRARE2质粒的BL21衍生菌,补充大肠杆菌缺乏的七种(AUA, AGG, AGA, CUA, CCC, GGA 及CGG)稀有密码子对应的 tRNA,提高外源基因、尤其是真核基因在原核系统中的表达水平。(已经携带有氯霉素抗性质粒) 当要表达的蛋白质需要形成二硫键以形成正确的折叠时,可以选择K–12衍生菌Origami 2系列,thioredoxin reductase (trxB) 和glutathione
原核表达和真核表达 原核表达的步骤和主要试剂 1、获得目的基因; 2、构建重组表达载体; 3、获得含重组表达质粒的表达菌种; 4、诱导表达; 5、包涵体的分离。 获得目的基因 PCR法钓基因 引物(捷瑞),普通taq酶(Regene,Fermentas)/Pfu 酶(Fermentas)/Hotstart Taq酶 (Fermentas),dNTP(Regene,Fermentas) RT-PCR法获取基因 Trizol(Invitrogen,捷瑞),DEPC(捷瑞或其他进分), Rnase抑制剂(MBI),M-MLV/AMV(Fermentas)或 M-MLV/AMV一步法RT-PCR试剂盒(捷瑞、Qiagen、 Axygen、Omega) 构建重组表达载体 内切酶(Fermentas),载体(捷瑞,Fermentas),琼脂糖(西班牙),T4连接酶(Fermentas,promega),测序 获得含重组表达质粒的表达菌种 菌株(捷瑞),质粒小抽试剂盒(捷瑞、Qiagen、Axygen),感受态细胞制备试剂盒(捷瑞),抗生素(捷瑞,Amresco,sigma),测序(伟沃) 诱导表达 IPTG(捷瑞,Amresco,sigma),胰蛋白胨(Oxiod),酵母粉(Oxiod),琼脂(进分,国产) 包涵体的分离 Triton X -100(捷瑞,进分),PMSF(sigma),DTT (Amresco,进分),尿素(Amresco,进分,sigma),还原型谷胱甘肽(Amresco,进分,sigma) 真核表达 蛋白质在真核表达的过程中,可以通过真核细胞的转录加工系统获得具有一定构象和生物活性的蛋白质。真核表达现在主要有毕赤酵母表达体系、杆状病毒昆虫细胞表达体系、哺乳动物细胞表达体系。 各种表达系统各有其优缺点,酵母和昆虫细胞表达系统蛋白表达水平高,生产成本低,但它们的加工修饰体系与哺乳动物细胞不完全相同;哺乳动物细胞产生的蛋白质更接近于天然蛋白质,但其表达量低、操作繁琐。各种表达系统由于翻译后的加工不完全相同,因而产生的重组蛋白的生物学活性和免疫原性有时会有差别。 毕赤酵母表达体系步骤及试剂 1、目的基因的获取; 2、构建表达质粒载体; 3、表达质粒的转化; 4、转化子的鉴定; 5、表达产物的鉴定。 目的基因的获取 基因组DNA提取 酚/氯仿法 蛋白酶K(Merk和Amresco),饱和酚(捷瑞), 氯仿﹕异戊醇(24﹕1)(捷瑞) 试剂盒法 血液/动物/植物/微生物基因组DNA提取试剂盒 (捷瑞、Qiagen、Axygen、Omega) RNA的提取 Trizol法 Trizol(Invitrogen,捷瑞),DEPC(捷瑞或其 他进分),Rnase抑制剂(MBI)
蛋白质的表达、分离、纯化实验 标签: 蛋白质 表达 分离 纯化 蛋白质表达、分离、纯化可以:(1)探索和研究基因的功能以及基因表达调控的机理;(2)供作结构与功能的研究;(3)作为催化剂、营养剂等。 详细 实验方法 原核表达法 实验方法原理 携带有目标蛋白基因的质粒在大肠杆菌BL21中,在 37℃,IPTG 诱导下,超量表达携带有6个连续组氨酸残基的重组氯霉素酰基转移酶蛋白,该蛋白可用一种通过共价偶连的次氨基三乙酸(NTA )使镍离子(Ni 2+)固相化的层析介质加以提纯,实为金属熬合亲和层析(MCAC )。蛋白质的纯化程度可通过聚丙烯酰胺凝胶电泳进行分析。 实验材料 大肠杆菌BL21 试剂、试剂盒 LB 液体培养基氨苄青霉素Washing BufferElution BufferIPTG 蒸馏水胰蛋白胨酵母粉氯化钠 仪器、耗材 摇床离心机层析柱离心管移液枪枪头盒烧杯玻璃棒 实验步骤 一、试剂准备 1. LB 液体培养基:Trytone 10 g , yeast extract 5 g ,NaCl 10 g ,用蒸馏水配至1000 mL 。 2. 氨苄青霉素:100 mg/mL 。 3. 上样缓冲液:100 mM NaH 2PO 4,10 mM Tris ,8M Urea ,10 mM 2-ME , pH8.0。 4. Washing Buffer :100 mM NaH 2PO 4,10 mM Tris ,8 M Urea ,pH6.3。 5. Elution Buffer :100 mM NaH 2PO 4,10 mMTris ,8M Urea , 500 mM
真核基因和原核基因表达调控的异同? 真核基因表达调控的基本原理与原核基因相同,主要表现在: 1、与原核基因的调控一样,真核基因表达调控也以转录水平调控为最重要; 2、在结构基因均有调控序列,并依靠特异蛋白因子与这些调控序列的结合与否调控基因的表达。 3、都要经历转录、翻译的过程。 4、表达过程都有复杂性,多环节 不同 1、真核基因表达调控过程更复杂。 2、在染色质结构上。原核细胞的DNA是裸露的,而真核细胞DNA包装在染色体中。DNA与组蛋白组成核小体形成为染色体基本单位。在原核细胞中染色质结构对基因的表达没有明显的调控作用,而在真核细胞中染色质的变化调控基因表达,并且基因分布在不同的染色体上,存在染色体间基因的调控问题; 3、真核生物中编码蛋白质的基因通常是断裂基因,含有有非编码序列即内含子,因而转录产生的mRNA前体必须剪切加工才能成为有功能的成熟的mRNA,而不同拼接方式的可产生不同的mRNA。而原核生物的基因由于不含有外显子和内含子,因此,转录产生的信使RNA不需要剪切、拼接等加工过程。 4、在原核基因转录的调控中,既有正调控,也有负调控,二者同等重要,而真核细胞中虽然也有正调控成分和负调控成分,但目前已知的主要是正调控,且一个真核基因通常都有多个调控序列,必须有多个激活物同时特异地结合上去才能调节基因的转录; 5、原核基因的转录和翻译通常是相互偶联的,而真核基因的转录与翻译在时空上是分开的,从而使真核基因的表达有多种调控机制。 6、真核生物细胞中存在mRNA的稳定性调控
7、真核生物大都为多细胞生物,基因的表达随细胞内外环境条件的改变和时间程序在不同的表达水平上进行着精确调控,而原核生物主要受环境因素和营养状况影响基因调控。 8、真核生物由三种RNA聚合酶分别负责三种RNA的转录,而原核生物只有一种。
4000字。 技术方法类(包括分析方法类)释文提纲: (1)定义及概述(不设标题) (2)技术或方法原理(及发展历史) (3)应用及注意事项 意见:基本符合百科全书的要求。 真核细胞表达系统(Eukaryotic expression system)运用基因工程技术手段,在体外将外源基因分子插入病毒、质粒等载体分子,形成新的遗传物质组合,并导入真核细胞中实现外源基因蛋白表达的技术系统。按照宿主细胞类型,真核细胞表达系统包括酵母表达系统、丝状真菌表达系统、昆虫细胞表达系统和哺乳动物细胞表达系统。真核细胞表达系统是重组蛋白表达的有利工具,在基础科学及医药领域发挥了重要作用。 技术或方法原理(及发展历史) 酵母表达系统酵母是低等的单细胞真核模式生物,已完成基因组测序,遗传背景清晰,具有生长繁殖快,成本低,易于实现高密度培养和大规模发酵的优点。目前发展成熟的酵母表达系统主要包括酿酒酵母表达系统和毕赤酵母表达系统。酿酒酵母在食品工业领域的使用已有数千年历史,被美国FDA确认为安全性生物。1981年,Hitzeman等将成功将人干扰素基因导入酿酒酵母细胞,揭开了酿酒酵母表达系统的研究和应用历史,迄今已有多种外源蛋白在酿酒酵母中获得表达。然而酿酒酵母表达系统存在缺乏强启动子,质粒易丢失,分泌效率低,且富含高甘露糖型超糖基化等不足,导致表达产物存在过度糖基化的局限性,而逐渐被巴斯德毕赤酵母等甲醇营养型酵母表达系统所取代。巴斯德毕赤酵母来源于野生型石油酵母NRRL-Y11430,为子囊菌类单倍体酵母,富含甲醇代谢必须的醇氧化酶、过氧化氢酶、二羟丙酮合成酶的过氧化物酶系,能在以甲醇为唯一碳源的简单培养基上快速生长,属于甲醇营养型酵母的一种。基因工程常用的毕赤酵母菌株主要包括GS115(Mut+His—)、X-33(Mut+His+)、KM71(Muts His—)、SMD1168(his4 pep4)等。目前毕赤酵母表达系统的启动子有醇氧化酶AOX1启动子(Ellis et al., 1985)及三磷酸甘油醛脱氢酶GAP启动子,外源基因通过同源重组整合到酵母染色体基因组上,外源蛋白表达水平受甲醇或葡萄糖的严格调控。表达产物有胞内表达和胞外分泌两种方式,胞内表达适合于通常在胞浆中表达或不含二硫键的非糖基化蛋白,胞内表达载体有pPIC3、pPSC3k、pHIL-D1、pAO815 等[5]。分泌型表达载体在酵母α因子信号肽的引导下,将成熟的蛋白质分泌到细胞外,分泌型表达载体有pPIC9K、pHIL-S1p、YAM7SP6、pGAPZa、pPICZα等。毕赤酵母表达系统具有操作简便、遗传稳定、表达量高、对目的蛋白质进行糖基化、二硫键形成等翻译后修饰功能,使表达的重组蛋白具有同天然蛋白相似的生物学功能,在非糖活性蛋白生产上具有显著优势。 昆虫细胞表达系统 昆虫细胞具有与高等真核生物细胞相似的翻译后修饰、加工及转移蛋白质的能力。昆虫细胞表达系统主要利昆虫细胞、昆虫整体和杆状病毒结构基因中多角体蛋白的强启动子构建的表达载体,可使很多真核目的基因得到有效甚至高水平的表达。它具有真核表达系统的翻译后加工功能,如二硫键的形成、糖基化及磷酸化等, 使重组蛋白在结构和功能上更接近天然蛋白; 其最高表达量可达500mg/L;可表达非常大的外源性基因(- 200kD); 具有在同一个感染昆虫细胞内同时表达多个外源基因的能力; 对脊椎动物安全。由于病毒多角体蛋白在病毒总