
实验六转座子引起的插入突变
一、实验目的
通过实验进一步认识转座子的遗传学效应之一:转座可引起插入突变。
二、实验原理
转座子(transposon,Tn)是存在于染色体DNA上可自主复制和移位的基本单位。最简单的转座子不含有任何宿主基因而常被称为插入序列(insertion sequence,IS),它们是细菌染色体或质粒DNA的正常组成部分。一个细菌细胞常带有少于10个IS序列。转座子常常被定位到特定的基因中,造成该基因突变。
复合式转座子(composite transposon)是一类带有某些抗药性基因(或其他宿主基因)的转座子,其两翼往往是两个相同或高度同源的IS序列,表明IS序列插入到某个功能基因两端时就可能产生复合转座子。一旦形成复合转座子,IS 序列就不能再单独移动,因为它们的功能被修饰了,只能作为复合体移动。转座时发生的插入作用有一个普遍的特征,那就是受体分子中有一段很短的(3-12bp)、被称为靶序列的DNA会被复制,使插入的转座子位于两个重复的靶序列之间。
转座作用的遗传学效应有如下几个方面,转座引起插入突变;转座产生新的基因;转座产生染色体畸变;转座引起的生物进化。
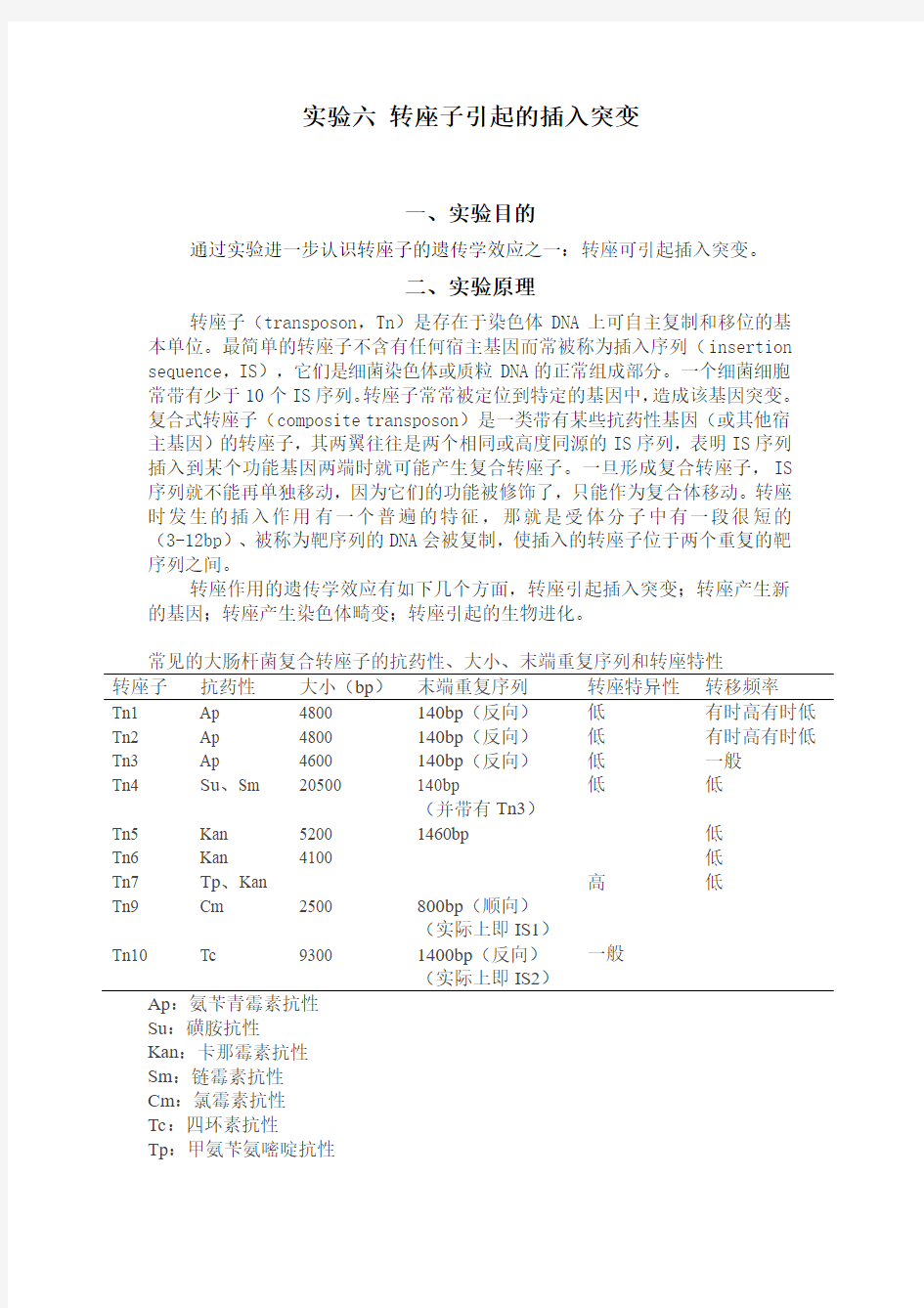
常见的大肠杆菌复合转座子的抗药性、大小、末端重复序列和转座特性
转座子抗药性大小(bp)末端重复序列转座特异性转移频率
Tn1 Ap 4800 140bp(反向)低有时高有时低Tn2 Ap 4800 140bp(反向)低有时高有时低Tn3 Ap 4600 140bp(反向)低一般
低低
Tn4 Su、Sm 20500 140bp
(并带有Tn3)
Tn5 Kan 5200 1460bp 低
Tn6 Kan 4100 低
Tn7 Tp、Kan 高低
Tn9 Cm 2500 800bp(顺向)
(实际上即IS1)
Tn10 Tc 9300 1400bp(反向)
一般
(实际上即IS2)
Ap:氨苄青霉素抗性
Su:磺胺抗性
Kan:卡那霉素抗性
Sm:链霉素抗性
Cm:氯霉素抗性
Tc:四环素抗性
Tp:甲氨苄氨嘧啶抗性
大肠杆菌复合转座子的插入突变有两个表型效应:基因突变和由转座子带来的抗药性。例如将Tn5插入到F’lac+ pro+/Str s的乳糖发酵基因中去以引起lac突变,可先将这F’因子转移到Δ(lac pro)Str r受体细胞中去,再观察是否发生了插入突变。在能排除供体的含链霉素的培养基上选择pro+受体菌落,若是抗卡那霉素的,那就是说受体细菌不但接受了F’因子,而且这F’因子带有Tn5,如果它又由lac 变为lac-,不能在以乳糖为唯一碳源的培养基上生长,那么,这一插入位置就是在乳糖发酵基因中。
为进一步提高效率,还可以采取一种措施,使在特定条件下lac+细菌不能生存而lac-细菌能够生存。已经知道gal E突变型在有乳糖和半乳糖存在的情况下,由于细胞中积累有毒的半乳糖代谢中间产物UDP-gal和gal-1-P而死亡,所以如果采用gal E突变型作为受体细菌,在含有0.02%乳糖和0.2%甘油的基本培养基上,受体细菌如果接受了一个F’lac+pro+,就不能生存。如果lac 基因由于插入了Tn5而成为lac-,那么受体细菌便能生存下来。
三、实验材料
1、菌株
受体菌:FD1006-Δ(lac proB) gal E str r
供体菌:Tn5·Gmlβλ-F’lac+pro+/Δ(proB lac)Val-
2、培养基
a、LB培养液
b、生理盐水
c、A培养基:含链霉素、葡萄糖的基本培养基
d、B培养基:含链霉素、卡那霉素、乳糖、甘油的基本培养基
2 4
KH2PO445g
(NH4)2SO410g
柠檬酸钠·2H2O 5g
去离子水定容至1L
pH7.0
3、仪器和耗材
恒温摇床、250mL三角瓶、离心管、培养皿、超净工作台
四、实验步骤
1. 第一天傍晚,分别接一环供体菌和一环受体菌于两支5mL LB培养液中,37℃
摇床上培养过夜。
2. 第二天,取两个内含40mL LB培养液的250mL三角瓶,分别加入1mL过夜培
养的供体菌及受体菌,在37℃摇床上培养2-3h。
3. 将新鲜培养的供体菌及受体菌各取0.5mL,在1.5mL离心管中混合,37℃轻
摇培养60min。吸取100μL混合液涂B板。
4. 吸取10μL混合菌液至含990μL生理盐水的离心管中混匀,依次稀释至10-2、
10-4、10-6倍。
5. 将供体菌及受体菌分别吸取100 μL涂于B培养基上作为对照。再从10-4、10-6
倍稀释液中各吸100 μL分别涂于A培养基平板上。37℃培养2d。
6. 观察、统计并计算:
1) 接受了供体F’lac+pro+的受体菌的数目:A板上生长的菌落数(个/mL)。
2) 在B板上生长的菌落数(个/mL),即Tn5插入到lac基因上菌落数。
3) 计算Tn5转座到lac上的转座频率,在B板上生长的菌落数/在A板上生长
的菌落数。
五、作业
1、列表记录观察结果。
2、记录A、B板上菌落数,计算Tn5易位到lacZ和lacY上引起插入突变的总频
率。
3、画出Tn5从染色体转移到F’lac上所引起lac-突变的示意图。
文章编号:042727104(2008)0320301205 收稿日期:2007207213 作者简介:尹 隽(1969—),女,讲师;通讯联系人钟 江,男,教授,E 2mail:jzhong@https://www.doczj.com/doc/533636661.html,. Tn5转座子插入p95基因不影响 AcMNPV 的复制 尹 隽,胡志鹏,宋大新,钟 江 (复旦大学生命科学学院微生物学与微生物工程系,上海200433) 摘 要:从Tn5转座子介导的AcMNPV 随机插入突变体库中,分离到一株复制正常的突变体AcApra41.突变定位发现Tn5转座子插入了病毒p95基因中.为了排除AcApra41中还有其他突变,利用同源重组法构建了p95基因定点插入突变的重组病毒AcGFP 2P95in.PCR 确认p95基因中插入了Tn5转座子;Westernblot 也证实A 2cApra41和AcGFP 2P95in 感染的细胞中,P95蛋白的分子量都因为插入突变而变小,由野生型的95ku 变为55ku.病毒复制动态曲线和荧光显微镜观察证实带有该插入突变的病毒能够在Sf9细胞中正常复制,并表达极晚期基因.这一结果表明完整的P95蛋白对病毒复制是非必须的. 关键词:杆状病毒;突变体;复制;基因表达 中图分类号:Q812 文献标识码:A 杆状病毒(Baculoviridae )是一类主要感染节肢动物的病毒,基因组为双链环状DNA,大小约为80~180kb [1].一方面它们作为基因表达载体已被广泛地用于生产各种蛋白质,另一方面它们也可以作为生物杀虫剂使用.在所有的杆状病毒中,苜蓿银纹夜蛾核型多角体病毒(Auto gra pha cali fornica multiplenucle 2opolyhedrovirus,AcMNPV )是被研究得最为深入的一种病毒,该种病毒的基因组大小为133894b p,包括156个潜在的阅读框[2].尽管有一些基因已经被研究得很清楚,比如病毒的结构蛋白,必需的顺式作用因子等[327],但还是有许多基因的功能尚不清楚.为了能高效地研究这些未知基因,我们用Tn5转座子介导的随机插入突变法构建了AcMNPV 突变库(李惠等[8],尹隽等,待发表). 本文研究了从突变体库中筛选到的一株p95基因带有插入突变的突变体.研究结果提示完整的P95蛋白对病毒在细胞中的复制是非必须的. 1 材料和方法 1.1 细胞、病毒和菌株 草地贪夜蛾细胞系Sf9细胞用TNM 2FH 培养基(Sigma 2Aldrich,MO,USA )补充10%的小牛血清、100U/mL 青霉素和100U/mL 链霉素培养.带有AcMNPV 基因组的质粒(bacmid )来源于Bac 2to 2Bac 系统(Invitro gen,CA,USA ),保存于大肠杆菌中,它转染Sf9细胞后可以复制产生感染性的AcMNPV [9].以在病毒多角体启动子下游插入绿色荧光蛋白基因(gfp )的bacmid (bacGFP )作为起始病毒基因组,利用体外转座系统构建了AcMNPV 随机插入突变体库(尹隽,发表中). E.coli BJ5183(RecA +)作为重组用菌 株,E.coli EC100(Epicentre,WI,USA )用于保存bacmid.用于培养含bacmid 菌株的抗生素浓度分别为:卡那霉素(Kan )50m g/mL,庆大霉素(Gen )7m g/mL,阿普拉霉素(Apra )20m g/mL. 1.2 Tn5转座子插入定位 参见李惠等[8],简单介绍如下.根据AcMNPV 基因组序列,全基因组每隔1.5kb 沿同一方向设计了88个引物(分别命名为v1~v88,具体序列略).在转座子上设计两个引物Tn 2u p (5′2GTCTCCGACCT 2 第47卷 第3期 2008年6月复旦学报(自然科学版)JournalofFudanUniversit y (NaturalScience )Vol.47No.3Jun.2008
转座子及其相关技术的研究 摘要:转座子是一类在细菌的染色体,质粒或噬菌体之间自行移动的遗传成分,是基因组中一段特异的具有转位特性的独立的DNA序列,转录组的活动对生物体基因组的转录以及演变存在着严重影响,本文就转座子的基因机理及特征、转座子沉默、转座子的标签技术以及其在植物中的运用进行阐述。 转座子是存在于DNA上可自主复制和移位的基本单位。MclCintockl’嗜次在玉米中的发现改变了人们对基因组序列稳定性的认识,打破了遗传物质在染色体上呈线性固定排列的传统理论。目前认为,多数生物体有自发突变且有重要表型效应出现的原因源于转座子的可动性,并且可以导致宿主基因组发生从点突变到染色体重排的一系列变化,转座子在进化上为建立宿主基因特性起着重要作用。 1.转座子特征与分类 基因转座时发生的插入作用中受体分子都有一段3-12bp的靶序列DNA会自我复制,使插入的转座子位于两个重复的靶序列之间。转座子可以分为两大类:以DNA-DNA方式转座的转座子和反转录转座子。第一类转座子可以通过DNA复制或直接切除两种方式获得可移片段,重新插入基因组DNA中。第二类转座子又称为返座元,在结构和复制上与反转录病毒类似,它通过转录合成mRNA,再逆转录合成新的元件整合到基因组中完成转座。 2转座子相关技术 2.1转座子分离方法 有4种方法用来分离转座子:(l)转座子诱捕法,此法适用于分离具有相当高的整合和切割频率的转座子。(2)Southern杂交法,此种方法需要有适当的探针,用于检测已知的转座子。(3)重复DNA序列鉴定法,适用于高拷贝数的无论是否有活性的转座子。(4)PcR扩增法,对己知序列的转座子可以设计引物直接PCR扩增。
实验六转座子引起的插入突变 一、实验目的 通过实验进一步认识转座子的遗传学效应之一:转座可引起插入突变。 二、实验原理 转座子(transposon,Tn)是存在于染色体DNA上可自主复制和移位的基本单位。最简单的转座子不含有任何宿主基因而常被称为插入序列(insertion sequence,IS),它们是细菌染色体或质粒DNA的正常组成部分。一个细菌细胞常带有少于10个IS序列。转座子常常被定位到特定的基因中,造成该基因突变。 复合式转座子(composite transposon)是一类带有某些抗药性基因(或其他宿主基因)的转座子,其两翼往往是两个相同或高度同源的IS序列,表明IS序列插入到某个功能基因两端时就可能产生复合转座子。一旦形成复合转座子,IS 序列就不能再单独移动,因为它们的功能被修饰了,只能作为复合体移动。转座时发生的插入作用有一个普遍的特征,那就是受体分子中有一段很短的(3-12bp)、被称为靶序列的DNA会被复制,使插入的转座子位于两个重复的靶序列之间。 转座作用的遗传学效应有如下几个方面,转座引起插入突变;转座产生新的基因;转座产生染色体畸变;转座引起的生物进化。 常见的大肠杆菌复合转座子的抗药性、大小、末端重复序列和转座特性 转座子抗药性大小(bp)末端重复序列转座特异性转移频率 Tn1 Ap 4800 140bp(反向)低有时高有时低Tn2 Ap 4800 140bp(反向)低有时高有时低Tn3 Ap 4600 140bp(反向)低一般 低低 Tn4 Su、Sm 20500 140bp (并带有Tn3) Tn5 Kan 5200 1460bp 低 Tn6 Kan 4100 低 Tn7 Tp、Kan 高低 Tn9 Cm 2500 800bp(顺向) (实际上即IS1) Tn10 Tc 9300 1400bp(反向) 一般 (实际上即IS2) Ap:氨苄青霉素抗性 Su:磺胺抗性 Kan:卡那霉素抗性 Sm:链霉素抗性 Cm:氯霉素抗性 Tc:四环素抗性 Tp:甲氨苄氨嘧啶抗性
转座子小综述 09生物技术一班汪晨皓 200915070123摘要 转座子又称跳跃因子,其实质是基因组上不必借助于同源序列就可移动 的 DNA片段,它们可以直接从基因组内的一个位点移到另一个位点。自 1951年美国McClintock在玉米中首先发现了 DNA转座子以来, 转座子已成为各种生物基因分析的有效工具之一[ 1]不仅可利用转座子诱变找到原核生物的单性生殖基因, 而且在真核生物中, 转座子的发现和运用极大地促进了果蝇遗传学的发展。人们已经应用各种方法, 在生物界各个领域证实了转座子系统的广泛存在[ 2]。利用转座子特有的转座功能, 将带有标记的转座子插入目的基因或基因组,产生了转座子标签技术、转座子定点杂交技术、转座子基因打靶技术和非病毒载体基因增补技术。人们利用这些技术, 可以确定基因组的功能、基因组间的功能差异;可以改变目的基因的活性, 获得转基因生物; 可以阻断毒力基因, 获得基因疫苗; 可以促进基因整合, 进行基因治疗等。转座子的发现改变了人们对基因组序列稳定性的认识, 打破了遗传物质在染色体上呈线性固定排列的传统理论。转座子插入新的位点后, 该位点附近的基因即受到抑制而呈现隐性的睡美人表型。一旦转座子在转座酶的作用下从这一位点上转走, 该位点的基因隐性表型又恢复为显性表型, 即睡 美人苏醒。调控转座酶和转座子活性的系统称为青蛙王子( Frog Pri nce) [ 3]。目前,认为多数生物体有自发突变且有重要表型效应出现都源于转座子的可动性, 并且可以导致宿主基因组发生从点突变到染色体重排的一系列变化。转座子在进化上为建立宿主基因特性起着重要作用。用特异的开放阅读框捕获技术, 可以使自然散在的转座酶编码基因高度表达,人为催化激活转 座子使其苏醒 , 执行插入、黏贴、切除等任务。目前已经应用于微生物、昆虫、植物、动物及人类基因组功能的研究[ 2], 例如蛙类基因组含有水手转座子超家族, 呈自然失活状态, 转座酶与转座增强子序列末端结合, 在蛋白协助下, 激活转座子, 使睡美人转座子苏醒[ 4]。 关链词 睡美人;转座子;相关技术;应用 转座子系统又称“睡美人”转座子系统,因该系统在自然状态下发生转座的能力不足, 大多数突变基因处于抑制“睡眠”状态而得名。由此发展起来的相关技术有转座子标签技术,定点杂交技术,转座子基因打靶技术, 非病毒载体基因增补技术即转座子“睡美人苏醒”技术。本文就转座子及其相关内容做一概要的总结。 转座子又称可移动基因, 跳跃基因, 是一种可在基因组内插入和切离并能改变自身位置的DNA序列。早在20 世纪50 年代, 首先由McClintock 在玉米中发现,从而改变了人们对基因组序列稳定性的认识,打破了遗传物质在染色体上呈线性固定排列的传统理论。目前生物体中所发现的10%的突变是由于它抑制其他基因的表达而形成的[ 5].转座子在进化上为建立宿主基因特性 起着重要作用。 转座子学说使McClintock荣获1983年诺贝尔生理医学奖[3].然而需要强 调的是, 并不是所有具有转座子基因的个体都可以发生转座子转座,转座能 力个体间有很大差别, 在有转座酶存在的情况下,通常情况受到一个激活因子Ac的控制。当细胞中有Ac时转座子才发生转座, 细胞中无Ac时转座子处于