核磁共振波谱对大米脱水素K片段在模拟膜中的结构研究
- 格式:pdf
- 大小:1.31 MB
- 文档页数:6
DOI:10.11895/j.issn.0253鄄3820.171384
核磁共振波谱对大米脱水素K片段在模拟膜中的结构研究
薛蓉摇吴亦洁摇李晓晶*
(中国科学院长春应用化学研究所,长春130022)
摘摇要摇脱水素是一类植物抗逆相关蛋白,可帮助植物抵抗干旱、低温、盐碱等环境胁迫。富含赖氨酸的K片段是脱水素中的保守功能片段,在低温保护和膜保护功能中起至关重要的作用。目前,脱水素及K片段
的作用机理仍不完全清楚。本研究采用圆二色谱(CD)和核磁共振波谱(NMR)方法及分子动力学模拟计算研究了具有抗菌活性的大米脱水素K片段在模拟膜中的三维结构及其与膜的结合方式。圆二色谱研究表明,水中呈现无规卷曲构象的K片段在模拟膜中会形成琢鄄螺旋结构。核磁共振结构研究进一步证实了K片段在模拟膜中的空间结构,即中间部分形成了两亲性的琢鄄螺旋结构,其中,疏水残基位于螺旋结构的一面,亲水残基位于螺旋结构的另一面。扩散排序(DOSY)核磁共振实验表明,K片段与膜在水溶液中形成了稳定的结合体;顺磁性探针检测表明,整个K片段插入膜中,其中疏水面朝向模拟膜的疏水核,其它部分朝向模拟膜亲水表层。本研究得到的K片段在模拟膜中的精细结构为理解环境胁迫下K片段及脱水素与膜的作用机理提供了重要信息。
关键词摇脱水素;核磁共振;三维结构;膜结合;分子动力学模拟;K片段
摇2017鄄11鄄03收稿;2018鄄01鄄24接受本文系国家自然科学基金项目(No.21305134)和吉林省自然科学基金项目(No.20160101074JC)资助*E鄄mail:xjli@ciac.ac.cn1摇引言
脱水素,又称二族胚胎发育晚期丰富蛋白(LEA蛋白),是一类与植物抗逆反应相关的重要蛋白质。
这类蛋白质在植物胚胎发育晚期表达量十分丰富,在环境胁迫(如干旱、低温、盐碱等条件)下也会大量
累积[1,2]。因此,脱水素被认为是在胁迫过程中对植物起保护作用的重要物质之一,是当前抗逆研究方
面备受关注的一类蛋白质。脱水素的显著特点是拥有一个或多个富含赖氨酸的保守性K片段。截掉K片段,会使得脱水素ERD10、RcDhn5和TaDHN鄄5对脱氧脱氢酶(LDH)的低温保护活性降低[3,4]。
K片段自身也可以对LDH起到低温保护作用[5]。近期研究表明,脱水素还具有抗菌活性,K片段在其
中起关键作用,且K片段自身具有类似全长蛋白的抵抗革兰氏阳性菌活性[6]。在遭受各种胁迫的情况
下,小麦脱水素的K片段通过阻止蛋白聚集对大肠杆菌起到保护作用,而且K片段也具有抗革兰氏阴
性、阳性菌和真菌的活性[7,8]。河岸葡萄的K片段可以在受到冻融损害时阻止膜融合,并在不改变膜的
流动性和表面可及性情况下降低膜转变温度,对膜起到重要的保护作用[9]。上述研究结果表明,在脱
水素的低温保护及抗菌功能中,K片段发挥着至关重要的作用。尽管体内及体外实验证明了脱水素及K片段的重要性,但目前对其分子作用机制仍不完全清楚。功能是蛋白质结构的生化表现特征,结构是
蛋白质行驶功能的基础。因此,研究脱水素及其重要功能区域的结构特征是全面认识其作用机制的有
效途径。
脱水素具有高度灵活的结构,它包含大量亲水残基,被认为是天然无规蛋白。然而,当环境发生改
变(如缺水或是与膜表面结合)时,脱水素可以形成一些有序结构[10]。K片段被预测可以形成两亲性的
螺旋结构,因此推测其与脱水素的结构改变密切相关[10]。详细探究K片段的精细结构对于揭示
K片段,甚至于整个脱水素的作用机理是非常必要的。核磁共振波谱(NMR)技术是原位研究溶液中蛋
白质三维结构的主要分析方法,它不仅可在原子水平上提供蛋白质在生理条件下的空间结构[11~13],还
可通过观察蛋白质上个别基团的行为特点为分析其分子相互作用等研究提供重要信息。本研究以具有
抗菌活性的大米脱水素K片段[6]作为研究对象,利用圆二色谱(CD)、NMR等多种谱学方法及分子动力第46卷2018年5月摇摇摇摇摇摇摇摇摇摇摇摇分析化学(FENXIHUAXUE)摇研究报告ChineseJournalofAnalyticalChemistry摇摇摇摇摇摇摇摇摇摇摇摇第5期664~669学模拟计算等手段研究了K片段在模拟膜SDS胶束中的三维结构及其与模拟膜的相互作用方式,为进
一步了解K片段乃至脱水素抗菌及膜保护机理提供了精细的三维结构及取向信息。
2摇实验部分
2.1摇仪器与试剂J鄄810圆二色谱仪(日本Jasco公司);AVANCE核磁共振波谱仪(600MHz,Bruker公司)。合成肽
K片段(KKGFLDKIKEKLPGGHKK,纯度>95%,苏州强耀生物科技有限公司)。重水(D2O,99.8%)、氘
代十二烷基磺酸钠(SDS鄄d25,98%),均购于CambridgeIsotopeLaboratories公司。实验用水为去离子水。
2.2摇实验方法
2.2.1摇远紫外圆二色谱(Far鄄UVCD)摇样品制备:(1)水溶液中样品制备:将适量K片段直接溶于水
中,肽的浓度为30滋mol/L。(2)SDS中样品制备:分别将适量K片段和SDS溶于水中,将肽溶液移入SDS溶液,旋转混匀,制得K片段浓度30滋mol/L、SDS浓度10mmol/L的水溶液样品。
圆二色谱检测所用石英比色皿的光程为0.1cm,波长扫描范围190~260nm;带宽1.0nm;实验检
测温度为25益。每个样品扫描3次,每张CD谱图取3次扫描图的平均值。CD谱图均已扣除背底,并
以平均残基摩尔椭偏率(兹MRE)表示:
兹MRE=兹obs/10lcn(1)
其中,兹obs(mdeg)是实验所测椭偏率,l(cm)是石英比色皿的路径长度,c(mol/L)是肽的浓度,n是多
肽所含氨基酸个数。
蛋白质二级结构含量通过CDPro软件分析得到。2.2.2摇核磁共振波谱摇样品制备:称取1.7mgK片段溶于220mL水中,称取31.9mgSDS鄄d25溶于275mL水中,将K片段溶液移入SDS鄄d25溶液,在混合溶液中加入55mLD2O,涡旋混匀,制得550mL的
10%D2O鄄90%H2O的混合溶液,其中,K片段浓度为1.5mmol/L、SDS鄄d25浓度为185mmol/L。
全相关谱(Totalcorrelationspectroscopy,TOCSY)和核的欧沃豪塞增强谱(NuclearOverhausereffectspectroscopy,NOESY)检测均采用压水峰技术,混合时间分别为80和150ms,累积次数分别为64次和
80次。采样数据矩阵2048´512,延迟时间2s,采样时间0.1548s,谱宽6613.8Hz。采用标准Bruker软
件(TOPSPIN2.1版本)进行谱图处理,处理后的谱图用SPARKY软件进行分析。选用TSP鄄d4作为内
标。实验温度为298K。
扩散排序(Diffusion鄄orderedspectroscopy,DOSY)核磁共振实验温度为298K,采用含有WATERGATE压水峰技术的BPPSTE(Bipolarpulsepairstimulatedecho)脉冲序列。扩散时间为200ms,
梯度脉冲持续时间为4.5ms,扫描次数为8。2.2.3摇结构计算摇三维空间结构通过CYANA软件(1.0.6版本4)计算得到。从200个随机的初始结
构开始计算,通过几何空间的模拟退火计算寻找符合构象约束的三维结构。得到的结构按目标函数值
从小到大进行排列,然后利用程序AMBER7对目标函数值排在前20的结构进一步进行能量优化,并用PROCHECK_NMR6程序对结构进行评估。利用MOLMOL7程序进行结构可视化分析和绘图。
3摇结果与讨论
3.1摇结构分析
许多与抗寒和抗旱相关的天然无规蛋白具有一个共同特点,即蛋白序列中都包含能促使蛋白与膜
结合的结构基序。玉米脱水素ZmDHN1的删除研究表明,K片段可能负责将脱水素蛋白结合到不同膜
上[10],但具体作用残基及结合机理仍不清楚。本研究采用CD谱表征K片段在水溶液中和SDS胶束溶
液中的二级结构。如图1所示,在水溶液中,K片段只在197nm处有一个负的最低峰,表现为典型的自
由卷曲结构;在SDS胶束溶液中,K片段在206和225nm出现两个负的最低峰,在192nm处出现一个
正的最高峰,表明K片段在SDS胶束溶液中主要以琢鄄螺旋结构存在[14]。利用CDPro软件中CDSSTR程
序对肽的二级结构含量进行计算,得到琢鄄螺旋含量为33.9%,茁鄄折叠含量为21.1%,转角含量为566第5期薛蓉等:核磁共振波谱对大米脱水素K片段在模拟膜中的结构研究摇摇15.8%,自由卷曲含量为28.8%。上述结果说明SDS胶束会诱导K片段由无规结构向琢鄄螺旋结构转
变,这与一些胁迫蛋白会在膜存在时形成琢鄄螺旋结构[15~17]相类似。
进一步利用NMR方法对K片段在SDS胶束溶液中的三维结构进行研究。根据W俟thrich提出的蛋
白质NMR谱序列识别方法[18],结合TOCSY给出的各氨基酸自旋体系的信息以及NOESY给出的欧沃
豪斯效应关系对NMR谱图进行归属。NOESY谱中大部分相邻氨基酸残基间都存在NOE交叉峰,如HNi鄄HNi+1、H琢i鄄HNi+1和H茁i鄄HNi+1,部分不相邻的氨基酸间也存在NOE交叉峰,如H琢i鄄HNi+3、
H琢i鄄H茁i+3和H琢i鄄HNi+2等(部分NOESY谱图归属见图2)。其中,连续的HNi鄄HNi+1NOE连接和H琢i鄄
HNi+3、H琢i鄄H茁i+3的NOE连接是反映琢鄄螺旋结构的特征连接模式,表明K片段在SDS胶束溶液中主要
以琢鄄螺旋结构形式存在。10
4
200姿(nm)兹MRE×10-2(mdeg·L/(cm·mol))
inH2O
inSDS
190210
22023024025026068
20-2-4-6-8-10-12-14
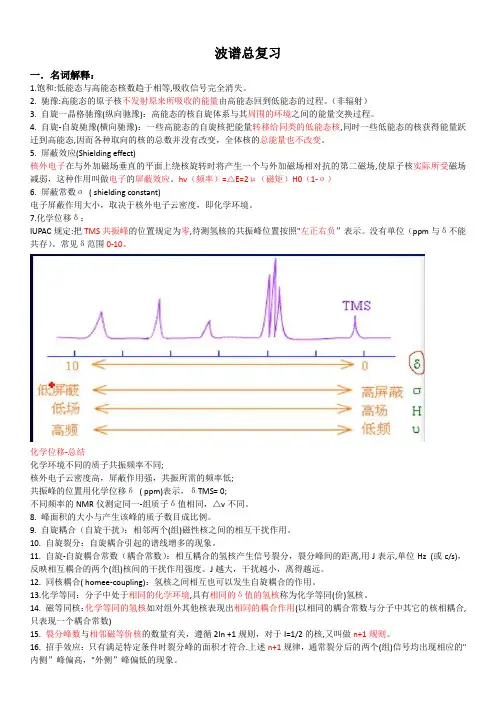
图1摇在室温条件下,30mmol/LK片段在水溶液中和10mmol/LSDS胶束溶液中的圆二色谱图Fig.1摇Circulardichroism(CD)spectraof
30mmol/LK鄄segmentinwaterand10mmol/LSDSmicellesatroomtemperature4.4啄(ppm)啄(ppm)
4.54.34.24.03.93.83.58.48.68.84.13.73.68.28.07.87.67.47.27.0
K2K2/K1G14G3H16/G15K9G3/K1G3/K2G14/P13K18D6F4G15/G14K17K11L12L12/K11L12/E10E10
K11/E10L12/K9E10/K7I8/L5K7F4/G3D6/L5G15E11/K9I8K9/I8E10/K9E10/I8
图2摇1.5mmol/LK片段在185mmol/LSDS胶束溶液中的H琢鄄HN区域局部NOESY谱图归属Fig.2摇H琢鄄HNregionofNOESYspectrumfor1.5mmol/LK鄄segmentin185mmol/Lsodiumdodecylsulfate(SDS)micelles
摇摇根据NOESY谱输出的NOE连接图(图3A)可见,琢鄄螺旋结构的特征连接模式主要存在于G3~L12
区域,表明此区域形成了琢鄄螺旋结构。进一步通过比较SDS胶束溶液中K片段各个残基的H琢化学位