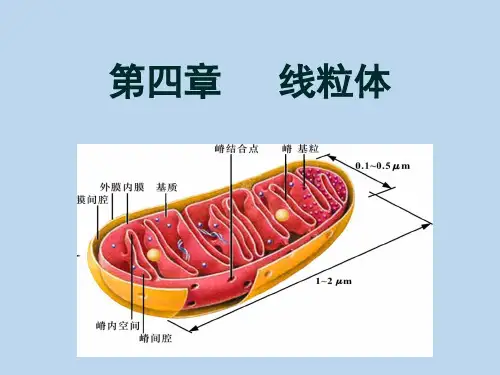
线粒体蛋白质的转运
- 格式:pdf
- 大小:1.47 MB
- 文档页数:14


细胞内蛋白质的分选和转运机制作者:陈建坤来源:《中学生物学》2018年第03期蛋白质作为生命活动的执行者和体现者,与生物的遗传、疾病等都有着重要关联。
在细胞内,有些蛋白质是先合成再进行分选转运,如线粒体、叶绿体、细胞核等结构中的蛋白质;而有些是边合成边分选转运,如分泌蛋白、膜蛋白等。
细胞根据蛋白质是否携有分选信号(信号序列)以及分选信号的性质,有选择地将蛋白质运送到细胞的不同部位。
1分选信号的种类分选信号有两类:①信号肽:蛋白质多肽链上一段连续的特定氨基酸序列,一般位于新肽链的N端,属于一级结构。
完成分选任务后常被切除。
②信号斑:位于多肽链不同部位的几个特定氨基酸序列经折叠后形成的斑块区,是一种三维结构。
完成分选任务后,仍然存在。
2原核细胞中蛋白质分选转运途径原核细胞(如细菌)没有复杂的生物膜系统。
但是为了维持生命,原核细胞需要合成一些蛋白质分泌到细胞质或者转运到细胞外发挥作用。
原核生物中蛋白质的转运分泌途径主要包括3种:①一般分泌途径,即SEC途径;②双精氨酸移位酶途径,即TAT途径;③信号颗粒识别途径,即SRP途径。
除此之外,还有V型分泌途径、TPS分泌途径和分子伴侣引导分泌途径等。
①一般分泌途径,即SEC途径:SEC分泌途径是原核生物中蛋白质主要的跨膜运输机制,主要由SEC移位酶作为介导。
SEC途径可以转运多种蛋白质,包括毒性因子、菌毛、黏附素和蛋白酶等。
SEC途径主要功能是把尚未折叠完成的蛋白质转运到质膜外,在质膜外折叠成有活性的蛋白质。
SEC途径大致可分为3个过程:信号序列的识别与定位、跨膜转运和多肽的释放。
②双精氨酸移位酶途径,即TAT途径:TAT途径识别的肽链N端信号序列通常含有两个连续的精氨酸残基。
TAT途径主要转运已经折叠完成的蛋白质,而尚未折叠完成的蛋白质通常不能通过该系统分泌,从而避免未完成折叠蛋白在胞外被降解的命运,保证了分泌产物的结构和功能的准确性。
此外,TAT途径还可以将少数蛋白质整合到质膜中。


蛋⽩质的合成、转运、修饰蛋⽩质的合成蛋⽩质的种类是由基因决定的,也就是说⼈类基因组有多少个基因,⼈体就有多少种蛋⽩质,只是蛋⽩质表达的时期和部位不同.根据⼈类基因组计划分析得知:全部⼈类基因组约有2.91Gbp,约有39000多个基因;也就是说⼈体蛋⽩质的种类有39000多种蛋⽩质⽣物合成可分为五个阶段,氨基酸的活化、多肽链合成的起始、肽链的延长、肽链的终⽌和释放、蛋⽩质合成后的加⼯修饰⼀.氨基酸的活化分散在胞液中的各种氨基酸需经特异的氨基酰-tRNA合成酶催化,ATP供能,并需Mg2+或Mn2+参与在氨基酸的羧基上进⾏活化,⽣成中间复合物()后者再与相应的tRNA作⽤,将氨基酰转移到tRNA分⼦的氨基酸臂上,即3′末端腺苷酸中核糖的3′(或2′)羟基以酯键相结合形成氨基酰-tRNA【氨基酰tRNA的⽣成】tRNA各种tRNA的⼀级结构互不相同,但它们的⼆级结构都呈三叶草形三叶草形结构的主要特征是:含有四个螺旋区、三个环和⼀个附加叉四个螺旋区构成四个臂,其中含有3′末端的螺旋区称为氨基酸臂,因为此臂的3′-末端都是C-C-A-OH序列,可与氨基酸连接三个环分别⽤Ⅰ、Ⅱ、Ⅲ表⽰环Ⅰ含有5,6⼆氢尿嘧啶,称为⼆氢尿嘧啶环(DHU环)环Ⅱ顶端含有由三个碱基组成的反密码⼦,称为反密码⼦环;反密码⼦可识别mRNA分⼦上的密码⼦,在蛋⽩质⽣物合成中起重要的翻译作⽤环Ⅲ含有胸苷(T)、假尿苷(ψ)、胞苷(C),称为假尿嘧啶环(TψC环);此环可能与结合核糖体有关tRNA在⼆级结构的基础上进⼀步折叠成为倒“L”字母形的三级结构起始因⼦原核起始因⼦只有三种(IF1、IF2、IF3)真核起始因⼦(简称为eIF)种类多且复杂,已鉴定的真核起始因⼦共有12种延长因⼦原核⽣物(简称EF)由三部分组成:EF-Tu,EF-Ts,和EF-GEF-Tu它介导氨酰-tRNA进⼊核糖体的空位EF-Ts充当EF-Tu亚基的鸟嘌呤核苷酸交换因⼦,催化EF-Tu释放GDPEF-G催化tRNA的移位和多肽延伸的每个循环后期mRNA从核糖体上掉下来真核⽣物(简称eEF)真核⽣物中分为:eEF-1和eEF-2eEF-1有两个亚基,α和βγα相当于原核⽣物中的EF-Tu亚基,它介导氨酰-tRNA进⼊核糖体的空位Βγ相当于原核⽣物中EF-Ts,核苷酸交换因⼦α,催化GDP从α上释放eEF-2相当于原核⽣物的EF-G,催化tRNA的移位和多肽延伸的每个循环后期mRNA从核糖体上掉下来终⽌因⼦(释放因⼦)原核⽣物细胞的释放因⼦(简称RF):识别终⽌密码⼦引起完整的肽链和核糖体从mRNA 上释放的蛋⽩质释放因⼦1(RF1):能识别终⽌密码⼦UAA和UAG⽽终⽌蛋⽩质合成的细菌释放因⼦释放因⼦2(RF2):能识别终⽌密码⼦UAA和UGA⽽终⽌蛋⽩质合成的细菌释放因⼦释放因⼦3(RF3):与延长因⼦EF-G有关的细菌蛋⽩质合成终⽌因⼦当它终⽌蛋⽩质合成时,它使得因⼦RF1和RF2从核糖体上释放真核⽣物细胞只有⼀种终⽌因⼦(称为eRF)能识别所有的终⽌密码⼦因为它没有与GTP结合的位点,所以它不能帮助完成合成的多肽从P位点的tRNA的释放在真核⽣物内可能还存在能与eRF合作、帮组多肽从核糖体释放的蛋⽩质核糖体的活性部位单个核糖体上存在四个活性部位,在蛋⽩质合成中各有专⼀的识别作⽤1.A部位:氨基酸部位或受位:主要在⼤亚基上,是接受氨酰基-tRNA的部位2.P部位:肽基部位或供位:主要在⼩亚基上,是释放tRNA的部位3.肽基转移酶部位(肽合成酶),简称T因⼦:位于⼤亚基上,催化氨基酸间形成肽键,使肽链延长4.GTP酶部位:即转位酶(EF-G),简称G因⼦,对GTP具有活性,催化肽键从供体部位→受体部位核糖体上还有许多与起始因⼦、延长因⼦、释放因⼦以及各种酶相结合的位点核糖体的⼤⼩是以沉降系数S来表⽰,S数值越⼤、颗粒越⼤、分⼦量越⼤原核细胞与真核细胞核糖体的⼤⼩亚基是不同的⼆.核糖体循环(肽链合成)1.肽链启动阶段在蛋⽩质⽣物合成的启动阶段,核蛋⽩体的⼤、⼩亚基,mRNA与⼀种具有启动作⽤的氨基酸tRNA共同构成启动复合体。

细胞内蛋白质的分选和运输蛋白质在细胞质基质中合成后,按其氨基酸序列中分选信号(sorting signal)的有无以及分选信号的性质被选择性地送到细胞的不同部位,这一过程称为蛋白质分选(protein sorting)和蛋白质靶向运输(protein targeting)。
另外,细胞外的蛋白质经胞吞作用进入细胞内部,也经历分选和靶向运输过程。
细胞中每一种蛋白质只有到达正确的位置才能行使其功能,如 RNA和DNA聚合酶必须送到细胞核中才能参与核酸的合成;酸性水解酶必须送到溶酶体才能进行大分子的降解作用。
因此,细胞内蛋白质的分选和运输对于维持细胞的结构与功能、完成各种细胞生命活动都是非常重要的。
细胞内蛋白质的分选信号以及运输途径和方式号肽通常引导蛋白质从细胞质基质进入内质网、线粒体和细胞核,同时也引导蛋白质从细胞核送回到细胞质基质以及从高尔基体送回到内质网;信号斑则引导一些其他分选过程,如在内质网合成的溶酶体酶蛋白上存在一种信号斑,在高尔基体的CGN中可被N-乙酰氨基葡萄糖磷酸转移酶所识别,从而使溶酶体酶蛋白上形成新的分选信号M-6-P,进一步在TGN中被M-6-P受体识别,并分选进入运输小泡最终送到溶酶体(详见第十章)。
每一种信号序列引导蛋白质到达细胞内一个特定的目的地(表10-1)。
要运送到内质网的蛋白质,在其N-末端有一段信号肽,其中间部分有5-10个疏水氨基酸。
带有这种信号肽的蛋白质,都会被运送到内质网,并进一步被运送到高尔基体,其中一部分蛋白质在C-末端还带有一个由4个氨基酸组成的信号肽,它们在高尔基体的CGN部位被识别并被送回内质网,是内质网驻留蛋白质;要运送到线粒体的蛋白质,在其N-末端带有一种信号肽,其信号序列中带阳电荷的氨基酸和疏水氨基酸呈交替排列;要运送到过氧化物酶体的蛋白质,在其C-末端有一种由三个特征性氨基酸组成的信号肽;要运送到细胞核的蛋白质,其信号肽中有一串带阳电荷的氨基酸,这一信号序列可位于蛋白质的任何部位。


细胞内蛋白质定位和转运机制细胞内蛋白质定位和转运机制是细胞内重要的生物学过程,它们维持了细胞的正常功能并参与了各种生物学活动。
本文将从细胞内蛋白质定位的基本原理、信号序列和定位机制以及蛋白质的转运机制等方面进行探讨。
一、细胞内蛋白质定位的基本原理细胞内蛋白质定位是指将蛋白质定向到细胞内特定的亚细胞结构或位置。
这一过程是通过特定的信号序列和机制实现的。
蛋白质定位的基本原理可以概括为两大类:靶向和扩散。
靶向是指蛋白质在合成过程中通过与一些特定的蛋白质或结构发生相互作用,从而被定向到细胞内的特定位置。
例如,细胞内的Golgi体是一个重要的分泌细胞器,某些蛋白质通过与Golgi体中的转运蛋白相互作用,从而被定位到Golgi体。
扩散是指蛋白质在合成过程中通过不断的扩散和分布,最终到达细胞内的特定位置。
这种定位机制主要依赖于蛋白质的物理和化学性质,以及细胞内各种蛋白质相互作用的平衡。
例如,细胞内水溶性蛋白质通过扩散和分布到达到达核内。
二、信号序列和定位机制在蛋白质的定位过程中,信号序列起到了非常重要的作用。
信号序列是蛋白质分子上的某一特定的氨基酸序列,它能够指导蛋白质被定位到特定的亚细胞结构或位置。
信号序列可以分为靶向信号序列和细胞内定位信号序列。
靶向信号序列通常位于蛋白质分子起始处,它能够与特定的蛋白质或结构发生相互作用,从而将蛋白质定向到细胞内的某个结构或位置。
细胞内定位信号序列通常位于蛋白质的内部,它能够改变蛋白质的物理和化学性质,从而影响蛋白质的定位。
蛋白质的定位机制可以分为几种类型:核定位、细胞质定位、内质网定位、线粒体定位、高尔基体定位等。
不同类型的定位机制通常与不同的信号序列和作用蛋白有关。
例如,核定位的信号序列通常富含正电荷氨基酸,而线粒体定位的信号序列则富含氨基酸序列(R-X-X-R)。
三、蛋白质的转运机制蛋白质定位到细胞内的特定位置后,往往需要通过转运机制到达目标位置。
蛋白质的转运可以分为受体介导转运和核孔复合物介导转运两种方式。


蛋白质的合成蛋白质的种类是由基因决定的,也就是说人类基因组有多少个基因,人体就有多少种蛋白质,只是蛋白质表达的时期和部位不同.根据人类基因组计划分析得知:全部人类基因组约有2.91Gbp,约有39000多个基因;也就是说人体蛋白质的种类有39000多种蛋白质生物合成可分为五个阶段,氨基酸的活化、多肽链合成的起始、肽链的延长、肽链的终止和释放、蛋白质合成后的加工修饰一.氨基酸的活化分散在胞液中的各种氨基酸需经特异的氨基酰-tRNA合成酶催化,ATP供能,并需Mg2+或Mn2+参与在氨基酸的羧基上进行活化,生成中间复合物()后者再与相应的tRNA作用,将氨基酰转移到tRNA分子的氨基酸臂上,即3′末端腺苷酸中核糖的3′(或2′)羟基以酯键相结合形成氨基酰-tRNA【氨基酰tRNA的生成】tRNA各种tRNA的一级结构互不相同,但它们的二级结构都呈三叶草形三叶草形结构的主要特征是:含有四个螺旋区、三个环和一个附加叉四个螺旋区构成四个臂,其中含有3′末端的螺旋区称为氨基酸臂,因为此臂的3′-末端都是C-C-A-OH序列,可与氨基酸连接三个环分别用Ⅰ、Ⅱ、Ⅲ表示环Ⅰ含有5,6二氢尿嘧啶,称为二氢尿嘧啶环(DHU环)环Ⅱ顶端含有由三个碱基组成的反密码子,称为反密码子环;反密码子可识别mRNA分子上的密码子,在蛋白质生物合成中起重要的翻译作用环Ⅲ含有胸苷(T)、假尿苷(ψ)、胞苷(C),称为假尿嘧啶环(TψC环);此环可能与结合核糖体有关tRNA在二级结构的基础上进一步折叠成为倒“L”字母形的三级结构起始因子原核起始因子只有三种(IF1、IF2、IF3)真核起始因子(简称为eIF)种类多且复杂,已鉴定的真核起始因子共有12种延长因子原核生物(简称EF)由三部分组成:EF-Tu,EF-Ts,和EF-GEF-Tu它介导氨酰-tRNA进入核糖体的空位EF-Ts充当EF-Tu亚基的鸟嘌呤核苷酸交换因子,催化EF-Tu释放GDPEF-G催化tRNA的移位和多肽延伸的每个循环后期mRNA从核糖体上掉下来真核生物(简称eEF)真核生物中分为:eEF-1和eEF-2eEF-1有两个亚基,α和βγα相当于原核生物中的EF-Tu亚基,它介导氨酰-tRNA进入核糖体的空位Βγ相当于原核生物中EF-Ts,核苷酸交换因子α,催化GDP从α上释放eEF-2相当于原核生物的EF-G,催化tRNA的移位和多肽延伸的每个循环后期mRNA从核糖体上掉下来终止因子(释放因子)原核生物细胞的释放因子(简称RF):识别终止密码子引起完整的肽链和核糖体从mRNA 上释放的蛋白质释放因子1(RF1):能识别终止密码子UAA和UAG而终止蛋白质合成的细菌释放因子释放因子2(RF2):能识别终止密码子UAA和UGA而终止蛋白质合成的细菌释放因子释放因子3(RF3):与延长因子EF-G有关的细菌蛋白质合成终止因子当它终止蛋白质合成时,它使得因子RF1和RF2从核糖体上释放真核生物细胞只有一种终止因子(称为eRF)能识别所有的终止密码子因为它没有与GTP结合的位点,所以它不能帮助完成合成的多肽从P位点的tRNA的释放在真核生物内可能还存在能与eRF合作、帮组多肽从核糖体释放的蛋白质核糖体的活性部位单个核糖体上存在四个活性部位,在蛋白质合成中各有专一的识别作用1.A部位:氨基酸部位或受位:主要在大亚基上,是接受氨酰基-tRNA的部位2.P部位:肽基部位或供位:主要在小亚基上,是释放tRNA的部位3.肽基转移酶部位(肽合成酶),简称T因子:位于大亚基上,催化氨基酸间形成肽键,使肽链延长4.GTP酶部位:即转位酶(EF-G),简称G因子,对GTP具有活性,催化肽键从供体部位→受体部位核糖体上还有许多与起始因子、延长因子、释放因子以及各种酶相结合的位点核糖体的大小是以沉降系数S来表示,S数值越大、颗粒越大、分子量越大原核细胞与真核细胞核糖体的大小亚基是不同的二.核糖体循环(肽链合成)1.肽链启动阶段在蛋白质生物合成的启动阶段,核蛋白体的大、小亚基,mRNA与一种具有启动作用的氨基酸tRNA共同构成启动复合体。
MTS:线粒体前提蛋白N端的基质导入序列/信号肽/导肽,富含疏水氨基酸:精氨酸(R)、赖氨酸(K)、丝氨酸(S)、苏氨酸(T)Hsp70家族的分子伴侣:与前体蛋白作用,防止前体蛋白在于外膜上的TOM复合体作用之前发生聚集或折叠线粒体膜上的主要转运蛋白复合物:外膜TOM,内膜TIMISTS:膜间腔导入序列线粒体的三种分裂方式:出芽分裂、收缩分裂、间壁分裂。
mtDNA随机分配,连续分裂的细胞中野生型mtDNA主导,分裂不旺盛的细胞突变型mtDNA占主导。
线粒体内共生说的证据:遗传系统与细菌相似(环状DNA、无组蛋白结合)、外膜上的孔蛋白、内膜上的心磷脂、蛋白质的合成方式细胞呼吸(cellular respiration)/生物氧化/细胞氧化(biological oxidation):在线粒体中,有氧气参与下,分解葡萄糖等各种小分子物质,产生二氧化碳和水,同时将分解代谢所释放的能量存储于ATP中。
细胞呼吸的过程:糖酵解、乙酰辅酶A的形成、三羧酸循环、氧化磷酸化糖酵解:胞质溶胶中,1葡萄糖+2NAD+2ADP+2Pi==2丙酮酸+2ATP+2NADH+2H+,底物水平磷酸化:高能磷酸键从底物转移到ATP中。
NADH+H+携带的质子和电子进入基质:苹果酸-天冬氨酸穿梭→NAD+甘油磷酸穿梭→FAD进入基质丙酮酸————→乙酰辅酶A三羧酸循环(TCA/Krebs循环/柠檬酸循环):线粒体基质,乙酰辅酶A+草酰乙酸==柠檬酸(3个羧基)。
循环一次,生成3个NADH+H、一个FADH2+1GTP,总共有两次循环。
膜上的呼吸链氧化磷酸化:NADH和FADH2携带的电子——————→O2+H—→水,电子传递释放能量—→ADP—→ATP呼吸链/电子传递链:电子传递体:只传电子递氢体:既传电子又传氢(NAD+、FAD)细胞色素c(膜表面)、泛醌(膜内):传递电子NADH是递氢体:1NADH+H+→3A TP(2.5)FADH2是递氢体:1NADH+H+→2A TP(1.5)一个葡萄糖彻底氧化产生的ATP数:底物水平磷酸化:4ATP氧化磷酸化:苹果酸穿梭:28ATP甘油磷脂酸穿梭:26ATP对氧化磷酸化过程中产生ATP的解释——化学渗透假说:。
蛋白质分选的共翻译转运途径方雪静俞如旺**(福建师范大学生命科学学院福建福州350117)摘要人教版普通高中生物学必修1(2019版)明确了分泌蛋白的起始合成是在游离的核糖体上,通过共翻译转运途径分泌到细胞外。
本文详细介绍蛋白质共翻译转运机制的探究史及指导共翻译转运的决定因素,描述共翻译转运的全过程和后续的膜泡运输。
关键词共翻译转运信号肽信号识别颗粒膜泡运输中国图书分类号:Q28 文献标识码:A核基因编码的蛋白质在胞质中游离的核糖体起始合成后,需要分选与转运到特定的功能位点,也称蛋白质寻耙(protein targeting)。
其分选途径大致可分为2条:①后翻译转运途径:蛋白质在胞质中完成翻译后才进行转运,线粒体、叶绿体及过氧化物酶体中的蛋白质多采用该途径进入细胞器;②共翻译转运途径:蛋白质先在游离的核糖体中合成一小段后,在信号肽的指导下边合成边转运到粗面内质网,经过加工最终分选到溶酶体、液泡 等细胞结构,有的分泌至胞外。
人教版普通高中生物学必修1(2019版)进一 步明确了分泌蛋白的起始合成是在游离的核糖体上,通过共翻译转运途径分泌到细胞外,纠正了旧 教材中的“最初是在内质网上的核糖体中由氨基酸合成肽链”。
本文将详细阐述该途径。
1共翻译转运机制的探究史20世纪60年代,乔治•帕拉德(George Palade)发现细胞质中的游离核糖体产生非分泌蛋白,而附着在内质网上的核糖体则产生分泌蛋白。
Palade的学生同特.布洛贝尔(G Q nter Blobel)认为产生该差异的原因不在于核糖体,而在于蛋白质自身[1]。
他从分泌蛋白转入内质网的过程人手,对蛋白质的转运展开研究,并抓住了2条线 索:①成熟蛋白质的氨基酸序列总比初生蛋白质更短一些;②蛋白质的翻译和转人内质网的过程似乎是同步进行的。
1975年,Blobel和戴维.萨巴蒂尼(David Saba-tini)提出“信号假说”,认为分泌蛋白的内质网靶向性是由蛋白质N端上的一个信号序列确定的。