呼吸的神经调节和化学调节
- 格式:docx
- 大小:301.02 KB
- 文档页数:9

第四节呼吸运动的调节要求:1、呼吸中枢及呼吸节律的形成。
2、外周和中枢化学感受器。
二氧化碳、H+和低氧对呼吸的调节。
呼吸运动是一种节律性的活动,其深度和频率随体内、外环境条件的改变而改变例如劳动或运动时,代谢增强,呼吸加深加快,肺通气量增大,摄取更多的O2,排出更多的CO2,以与代谢水平相适应。
呼吸为什么能有节律地进行?呼吸的浓度和频率又如何能随内、外环境条件而改变?这些总是是本节的中心。
一、呼吸中枢与呼吸节律的形成呼吸中枢是指中枢神经系统内产生和调节呼吸运动的神经细胞群。
多年来,对于这些细胞群在中枢神经系统内的分布和呼吸节律产生和调节中的作用,曾用多种技术方法进行研究。
如早期的较为粗糙的切除、横断、破坏、电刺激等方法,和后来发展起来的较为精细的微小电毁损、微小电刺激、可逆性冷冻或化学阻滞、选择性化学刺激或毁损、细胞外和细胞内微电极记录、逆行刺激(电刺激轴突,激起冲动逆行传导至胞体,在胞体记录)、神经元间电活动的相关分析以及组织化学等方法。
有管些方法对动物呼吸中枢做了大量的实验性研究,获得了许多宝贵的资料,形成了一些假说或看法。
(一)呼吸中枢呼吸中枢分布在大脑皮层、间脑、脑桥、延髓和脊髓等部位。
脑的各级部位在呼吸节律产生和调节中所起作用不同。
正常呼吸运动是在各级呼吸中枢的相互配合下进行的。
1.脊髓脊髓中支配呼吸肌的运动神经元位于第3-5颈段(支配膈肌)和胸段(支配肌间肌和腹肌等)前角。
很早就知道在延髓和脊髓间横断脊髓,呼吸就停止。
所以,可以认为节律性呼吸运动不是在脊髓产生的。
脊髓只是联系上(高)位脑和呼吸肌的中继站和整合某些呼吸反射的初级中枢。
2.下(低)位脑干下(低)位脑干指脑桥和延髓。
横切脑干的实验表明,呼吸节律产生于下位脑干,呼吸运动的变化因脑干横断的平面高低而异(图5-17)。
图5-17 脑干呼吸有关核团(左)和在不同平面横切脑干后呼吸的变化(右)示意图DRG:背侧呼吸组 VRH:腹侧呼吸组 NPBM:臂旁内侧核A、B、C、D为不同平面横切在动物中脑和脑桥之间进行横切(图5-17,A平面),呼吸无明显变化。


生理学研究人体各系统的生理功能及其调节生理学是研究生命体系结构、生物化学及各生理系统功能以及其调节的学科。
人体的各器官系统密不可分,通过相互作用保持了人体稳定的内外环境。
下文将从呼吸系统、心血管系统、消化系统及神经系统四个方面,阐述它们的生理功能及其调节。
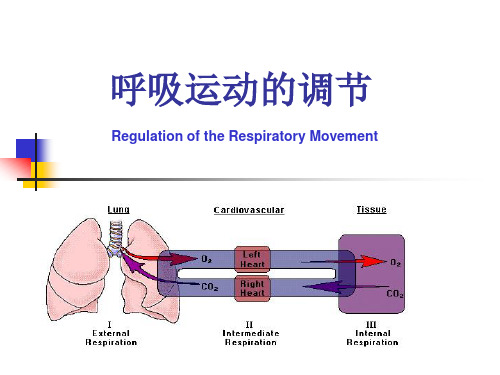
1. 呼吸系统呼吸系统是人体重要的生理系统之一,其主要的功能是为人体提供充足的氧气,通过排出二氧化碳来维持内环境。
呼吸过程由外呼吸和内呼吸两部分组成。
其调节主要体现在中枢神经系统和外周化学感受器。
外呼吸指的是空气在外部环境和肺部之间的交换,其调节主要通过神经系统。
中枢神经系统可以通过运动神经控制肋骨外展收缩,胸膜膨胀和收缩,使肺的容积发生变化而引起气流进出肺部。
此外,中枢神经系统还可以通过调节心率、血压等,进一步调控呼吸。
内呼吸就是指在组织细胞内发生的氧与二氧化碳的交换,其调节主要通过化学感受器。
当人体二氧化碳浓度升高或氧浓度降低时,感受器就会发送信号给中枢神经系统,促使呼吸频率和深度发生改变。
2. 心血管系统心血管系统包括心脏和血管两部分,其主要的功能是将氧和养分输送到组织细胞中。
心血管系统的调节主要体现在心肌细胞自身的特性及神经系统和荷尔蒙的影响。
心肌细胞自身的特性主要指自律性和兴奋性。
心脏可以自主地产生兴奋冲动,并通过导电组织传导到各部分,激发心肌细胞收缩。
此外,心肌细胞还具备内激素的调节能力,当心肌细胞伸展或缩短时,会产生肽类抑制因子和去甲肾上腺素等,这些物质可以调节心脏的兴奋性和收缩力。
神经系统和荷尔蒙是调节心血管系统的重要影响因素。
神经系统通过交感神经和副交感神经的作用影响心率和心肌收缩力。
而荷尔蒙主要是通过甲状腺素、肾上腺素和去肾上腺素等影响心肌细胞的收缩。
3. 消化系统消化系统是将食物转化为可被吸收利用的营养物质的重要系统。
消化过程主要由口腔、食管、胃及各种腺体组成,其调节主要涉及神经、激素及局部因素。
神经系统通过交感神经和副交感神经的作用来调节胃的运动。

讲稿:呼吸运动的调节【目的要求】1.观察各种理化因素对呼吸运动的影响。
2.分析各因素的作用途径,了解呼吸运动的调节机制。
【课堂提问及解答】1.调节呼吸运动的中枢?2.呼吸为什么有节律?3.调节呼吸运动的环节?答1:呼吸中枢是指(分布在大脑皮层、间脑、脑桥、延髓、脊髓等部位)产生和调节呼吸运动的神经细胞群。
正常呼吸运动是在各呼吸中枢的相互配合下进行的。
答2:呼吸节律形成的机制;基本呼吸节律形成的学说(1)起步细胞学说(2)N元网络学说等。
答3:呼吸运动的反射性调节包括(1)肺牵张反射(2)化学感受性反射调节(3)呼吸肌本体感受性反射(4)其他反射。
【实验原理】1.CO2↑→(+)中枢化学感受器、(+)外周化学感受器→(+)呼吸中枢→呼吸加深加快。
2.H+↑→(+)外周化学感受器、(+)中枢化学感受器→(+)呼吸中枢→呼吸加深加快。
3.O2↓→(+)外周化学感受器→(+)呼吸中枢→呼吸加深加快。
【重点难点】:呼吸运动调节的反射弧【观察指标】呼吸频率、幅度、PaO2、PaCO2、pH【方法与步骤】1.兔常规操作。
行气管插管和颈总动脉插管。
我们这里与以往不同的是,气管插管的一端通气口要与呼吸传感器相连,然后进入生物信号采集处理器,记录呼吸波。
颈总动脉插管插好以后,取血作血气分析,以作我们后面实验的对照。
我们取血的时候要注意抗凝和隔绝空气。
所以我们在取血前,要用肝素将注射器血管管壁湿润,取血的时候,将前面流出的几滴血弃去,取血后,立即将密封盖盖好,用手指弹一弹注射器血管壁,使血液与肝素混合,防止凝血,取完血后,大家还要记注,要用肝素将插管内的血液全部推回动脉。
2.增大无效腔。
等大家记录一段稳定的呼吸波后,并且已经取血做了血气分析,我们就可以做无效腔增大对呼吸的影响。
我们的器械盘里准备了一根长的橡胶管,将这根橡胶管连接在气管插管的(侧管)另一个通气口上,记录呼吸波形。
5分钟后从动脉插管处取血作血气分析。
然后,观察家兔的呼吸,等到它的呼吸恢复到正常以后,才可做下一步的实验,这个大概需要5-10分钟。

化学感受器是指其适宜刺激是化学物质的感受器。
参与呼吸调节的化学感受器因其所在部位的不同,分为外周化学感受器和中枢化学感受器。
1.外周化学感
受器颈动脉体和主动脉体是调节呼吸和循环的重要外周化学感受器。
在动脉血PO2降低、PCO2或H+浓度升高时受到刺激,冲动经寞神经和迷走神经传入延髓,反射性地引起呼吸加深加快和血液循环的变化。
其中,颈动脉体主要调节呼吸,而主动脉体在循环调节方面较为重要。
2.中枢化学感受器
延髓有一个不同于呼吸中枢但可影响呼吸的化学感受器,称为中枢化学感受器,以别于外周化学感受器。
中枢化学感受器位于延髓腹外侧浅表部位,左右对称,可以分为头、中、尾三个区。
中枢化学感受器的生理刺激是脑脊液和局部细胞外的H+.其有效刺激不是CO2本身,而是CO2所引起的[H+]的增加。
在体内,血液中的CO2能迅速通过血脑屏障,使化学感受器周围液体中的[H+]升高,从而刺激中枢化学感受器,再引起呼吸中枢的兴奋。
可是,脑脊液中碳酸酐酶含量很少,CO2与水的水合反应很慢,所以对CO2的反应有一定的时间延迟。
血液中的H+不易通过血脑屏障,故血液pH的变动对中枢化学感受器的直接作用不大,也较缓慢。
中枢化学感受器与外周化学感受器不同,它不感受缺O2的刺激,但对CO2的敏感性比外周的高,反应潜伏期较长。
中枢化学感受器的作用可能是调节脑脊液的[H+],使中枢神经系统有一稳定的pH环境,而外周化学感受器的作用主要是在机体低O2时,维持对呼吸的驱动。
呼吸的神经调节和化学调节人体的呼吸运动与心脏活动有相似之处,都是有节奏的、日夜不停的活动。
但这两种活动的起因却有很大的不同。
心肌有自动节律性,而产生呼吸运动的肌 肉都是骨胳肌,受躯体神经支配,没有神经的兴奋,呼吸肌(隔肌、肋间肌)都 不会自动收缩。
有节律的自动的呼吸活动起源于支配呼吸肌的运动神经元的有节 律的发放。
这种发放完全依靠来自脑的神经冲动。
如果在隔神经起点以上切断脊 髓,呼吸活动便会停止。
调节呼吸的神经机制有两类,一类是随意控制,另一类 是自动控制。
随意控制系统位于大脑皮层,它通过皮层脊髓束将冲动传送到呼吸 运动神经元。
大脑皮层可以有意识地控制呼吸活动的规式,例如有意的过度通气 或呼吸暂停(屏息)。
人们的语言活动必须有意地调整、改变呼吸规式才能发出 各种不同的声音。
刺激大脑皮层的某些区域可以引起呼吸活动的变比,还可以在 反射性呼吸活动的基础上建立条件反射。
这些都说明神经系统的高级中枢、大脑 皮层对呼吸活动进行着调节与控制。
自动控制系统位于脑桥和延髓。
1812年C. J. J. Legallois 发现,切除大脑、高位脑干的一部分以及小脑, 呼吸的节律性仍然存在。
他首先提出呼吸中枢的概念,并认为维持节律性呼吸的 中枢在延髓的中心。
以后一百多年来,生理学家用切除法、刺激法以及电描记法 等研究呼吸中枢的应置。
本世纪20年代初期,T. Lumsden 用精细的脑干切断法 得到了关于呼吸中枢的一些重要事实材料。
他在脑桥的上缘切断与中脑的联系如 图切面1,不影响麻醉动物的呼吸。
切断两侧迷走神经则引起呼吸变慢加深。
在脑桥的 上三分之一正在小脑脚之上切断与低位脑干的联系,而迷走神经保持完整,如图 切面2引起呼吸稍稍变慢变深。
如果再切断两侧迷走神经,则呼吸停止于吸气状 态,有时又被很短的呼气所打断。
他在脑桥的下缘切断脑干,完全排除脑桥的影吸中胸口者揩迷走神经完整 迷走神 经切断AM M 横叨痼干及切断迷走神经后的呼吸规式 飞.上丘1匚下丘CF.小脑脚 延髓中枢响,则有规律的呼吸节律消失,代之以喘息如图切面3。
在延髓下缘横切,引起 呼吸完全停止(见图)。
者揩横叨痼干及切断迷走神经后的呼吸规式 飞.上丘1匚下丘CF.小脑脚从根据这些事实当时得出下列结论:1 .在延髓有呼吸中枢,能发动及维持吸气、呼气。
2 .在脑桥的中部和下部有长吸中枢,如果不受控制、可以产生吸气痉挛或 长吸呼吸。
3 .在脑桥的上部有呼吸调整中枢,与迷走神经传入冲动一起周期性地抑制 长吸中枢的活动。
这种在延髓、脑桥存在三级呼吸中枢的理论曾经长期得到公认,后来经过进 一步的研究,提出了修正和补充。
有实验证明,在损毁呼吸调整中枢和切断迷走神经后恢复清醒的猫仍能保持 正常的平静呼吸,并不出现长吸呼吸,而且在长吸中枢并未发现神经元密集的核 团。
因此现在认为长吸呼吸是麻醉等因素造成的非正常状态,并不存在“长吸中 枢”。
延髓呼吸中枢许多实验表明,延髓是最基本的呼吸中枢。
切除延髓的一部分可以使呼吸停 止,刺激延髓的某些部分可以引起呼气或吸气活动。
在呼气或吸气时,可以在延 髓记录到电活动。
其中有些神经元在吸气时发放神经冲动,而在呼气时停止发放, 这些神经元叫做吸气神经元,另一些数目较少的神经元呼气时发放神经冲动,而 在吸气时停止发放,这些神经元叫做呼气神经元,呼吸的节律性是这两种神经元 交互作用的结果。
在延髓中呼吸神经元基本上集中在背腹两组的4个核团内。
一 组叫做背呼吸组,大部分是吸气神经元,集中在孤束核。
这些神经元接受第IX 、 [SCVSCJ 工 c 迷走神 经完整 I 由枢 延髓中枢中 施腼桥延A J ±X对脑神经的传入纤维。
这些神经传送来自肺、咽、喉部和外周化学感受器的感觉冲动。
背呼吸组的吸气神经元的轴突投射到对侧脊髓,通过膈运动神经元支配膈。
这些神经元也有轴突投射到腹呼吸组。
另一组是腹呼吸组,包括疑核和后疑核(NRA)的神经元,其中既有吸气神经元,也有呼气神经元。
疑核接受第IX、X对脑神经的传入投射,其轴突也通过这两对神经支配同侧咽部呼吸辅助肌。
后疑核前部神经元主要是吸气神经元,大部分轴突交叉下行到对侧脊髓支配外肋间肌运动神经元,小部分支配同侧膈运动神经元,驱动外肋间肌和膈肌。
后疑核尾部神经元主要是呼气神经元,其轴突也交叉下行到对侧脊髓的内肋间肌运动神经元和其它呼气肌运动神经元(见图)。
现有的资料说明延髓内并不存在过去认为的单一的对称的吸气中枢和呼气中枢,而是有若干个解剖上和机能上都各有特点的呼吸神经元群。
呼吸调整中枢位于脑桥中部的呼吸调整中枢包括臂旁内侧核(NPBM)和Kolliker-Fuse 核。
电刺激这个区域引起吸气停止,说明此区可以调整延髓呼吸中枢的活动。
吸气时延髓吸气神经元发出冲动上行到呼吸调整中枢使之产生冲动下行至延髓抑制吸气神经元,使吸气向呼气转换。
呼吸中枢的节律性活动是怎样产生的,目前仍是一个尚未阐明的问题。
有两种可能:一是由起搏细胞引起的自动节律性活动,但在脊椎动物中目前尚无法证明;二是由局部神经元回路的正负反馈所控制,目前有几种假说,尚待深入研究。
黑-伯二氏反射黑-伯二氏反射又叫肺牵张反射,是由E. Hering和J. Brener在1866年发现的。
这个反射有两个组成部分:1 .向肺内吹气使肺膨胀,引起膈肌收缩的抑制,呼吸停止在呼气的位置。
2 .从肺内抽气使肺萎缩,引起膈肌的强烈收缩,出现深吸气(见图)。
在肺扩张时刺激了气管平滑肌中的牵张感受器,冲动由迷走神经传入延髓, 抑制吸气神经元,切断吸气,引起被动呼气。
呼气,特别是深呼气时肺萎缩,对 牵张感受器的刺激减弱,传入冲动减少,解除了对吸气机制的抑制,吸气神经元 兴奋,引起吸气。
这个反射可能提供了呼吸自找调节的机制。
切除迷走神经后呼 吸变得缓慢而深沉,就是由于失去了这类牵张反射的负反馈机制所引起的。
在不同的动物中肺牵张反射的作用不同,现在知道兔的最强,人的最弱。
人 在正常平静呼吸时肺牵张反射不起作用,但婴儿出生的最初几天存在这一反射。
呼吸的化学调节20世纪初,Haldanc 学派在呼吸的化学调节方面做了许多工作,发现呼吸气 体成分的变化与血液成分的改变对呼吸有重要的影响,特别是二氧化碳的作用尤 为显著。
让受试者对着气囊反复呼吸一定量的空气,其中二氧化碳逐渐增加,氧逐渐 减少。
当二氧化碳增加到3%,氧减少到17%时,呼吸明显增强;当二氧化碳增 到5.6%,氧减少到14.8%时,呼吸非常显著地增强,甚至到达不可忍受的程度。
如果用钠石灰把呼吸排出的二氧化碳吸收,二氧化碳为零,只有氧的减少。
在这 种情况下,氧要减少到10%以下才能引起呼吸明显的变化。
如果呼吸气体中氧 的成分高于大气中氧的成分,二氧化碳增多时也会出现显著的呼吸增强(见图)。
上色:肺吹胀和雯缩对呼吸的影响11点线表示气音内的压力, 实线表示膈的运动口 下图:肺充气时,迷走神经单根纤维 的动作电位。
用10口室升空气在人处吹胀猫肺,从E 点开始保 持恒定的肺容积,在C 点突爆放气口作位动电 流省压 气气内7.3.7.2. 7, 1 7.0 6.9 6.8 6.7清醒健康A人呼空气中二氧化碳浓度,逐渐升高时的反应以及氧浓度逐渐降债时的反应从这些实验可以看出、呼吸的增强几乎完全是由于二氧化碳的增加。
相反,单纯的缺氧并不引起呼吸困难的先兆,但可能由于缺氧而突然引起昏迷。
肺泡气中二氧化碳分区是很稳定的,如稍有增加,呼吸就会显著加强。
对于健康的人,肺泡气中二氧化碳分区每改变l毫米汞柱就会引起通气量平均2.5升每分钟的改变。
如肺泡气内二氧化碳分压增加约5毫米汞柱,则呼吸活动增加100%。
肺泡气内二氧化碳分压低于正常水平会引起呼吸暂停。
二氧化碳是怎样影响呼吸活动的呢?1927年Heymans父子用含有丰富二氧化碳的血液灌注狗的孤离的肺循环,没有发现呼吸的变化。
但灌注孤离的左心室和主动脉弓,或灌注孤离的颈动脉体,呼吸就会增强。
在颈动脉体和主动脉体中有化学感受器,能感受血液中氧、二氧化碳和氢离子浓度的变化,反射性地引起呼吸活动的改变。
不过颈动脉体和主动脉体中的化学感受器只对含大量二氧化碳的血液发生反应,对小得多的血液二氧化碳含量的生理性变化不起反应,不会由于肺泡气中二氧化碳分压仅仅升高5毫米汞柱就引起通气量增加一倍的反应。
既然血液中二氧化碳含量的生理性变化不能刺激外周化学感受器,那必然是刺激了中枢神经系统的某些区域。
1960年Mitchell等人确定猫、狗的这种中枢化学感受器位于延髓腹表面,靠近第皿一XI对脑神经发出处(见图)。
在这个区域,局部施加氢离子、二氧化碳、ACh或烟碱可在几秒内刺激呼吸活动,而局部冷却或麻醉很快就会使呼吸下降。
由于血液与脑细胞之间存在血脑屏障,脂溶性物质,如呼吸气体、麻醉剂可以自由通过而离子不易通过。
因此,在正常情况下,中枢化学感受器细胞周围的细胞外液的酸度变化只能由流经脑的血液中二氧化碳分压改变所引起。
血液中二氧化碳分压升高,二氧化碳穿过血脑屏障进入中枢化学感受器细胞,在细胞内水合成碳酸,再离解成氢离子和碳酸氢根,使细胞内氢离子浓度升高,刺激中枢化学感受器细胞(见图)。
中枢化学感受器细胞受®血液和脑脊液成分的双重歌向中枢化学感受器将传入冲动输送到呼吸中枢,引起呼吸活动增加。
二氧化碳是中枢化学感受器正常的兴奋剂,血液中二氧化碳分压过低就不能通过中枢比学感受器兴奋呼吸中枢,导致呼吸暂停,然而待久吸入高浓度的二氧化碳(3%)也会引起呼吸中枢麻痹。
氢离子对呼吸的调节也有作用。
从静脉快速注射酸性溶液引起过度通气,而注射氢氧化钠溶液则引起通气降低。
通气对氢离子浓度的变化不如对二氧化碳分压的变化敏感。
pH值要降低0.05-0.1才能引起通气明显的反应(见图一20)。
对孤离的颈动脉体灌注酸性溶液,反射性地增加呼吸,但pH值减低0.1才有效。
切断动物外周化学感受器的神经支配后注射酸性溶液仍然引起通气增加,这说明中枢的某些部位可能对氢离子敏感。
由于血液中氢离子浓度的变化不容易影响中枢化学感受器周围的脑脊液,氢离子可能是直接刺激呼吸中枢而引起相应的变化。
缺氧对中枢化学感受器没有影响,但用低氧血灌注孤离的颈动脉体导致通气增加。
Biscoe等曾仔细地研究了颈动脉体单根传入神经纤维在不同的动脉血氧分压时冲动发放的情况(见图)。
a I fi I ii200 400 800猫化学感受器传入神经单木脚维的发放频率(W /秒)与动脉血串张力,聿米汞柱)的关系E表示同i束中两根纤维;支与C都是单根纤舞,动脉血氧分压口当动脉血氧分压由80千帕(600毫米汞柱)降到13.33千帕(100毫米汞柱,海平面的正常值)时,冲动发放频率才稍有增加;而动脉血氧分压从13.33千帕(100毫米汞柱)继续降到4千帕(30毫米汞往)则冲动发放频率急剧上升。