外源基因的表达检测及遗传特性
- 格式:ppt
- 大小:252.50 KB
- 文档页数:53

外源基因的定位和转录调控分子机制随着基因工程技术的不断发展,外源基因的引入和转录调控机制也逐渐成为生物学研究的热点问题之一。
在这个过程中,外源基因的定位和转录调控分子机制就变得至关重要。
本文将从以下几个方面探讨这个问题。
一、外源基因定位的技术手段外源基因的定位是基因工程研究的重要前提。
常见的技术手段有:1. PCR扩增:PCR是一种常用的外源基因定位技术,通过PCR反应,可以从待测样品中扩增出有代表性的DNA序列。
2. Southern印迹:Southern印迹是一种高灵敏度的外源基因定位技术,通过DNA电泳分离和传递等步骤,可以将待测DNA分子固定在膜上,进而对其进行探针杂交和检测。
3. FISH法:FISH法是通过荧光标记外源DNA序列,能够直接标记分子的位置和状态。
该技术具有定位精度高、灵敏度高等优点。
以上三种技术手段可以相互补充,从不同角度确定外源基因的位置和数量。
二、转录调控的机制外源基因的转录调控是基因工程研究的另一个重要方面。
它涉及到各种不同的转录因子、蛋白质和RNA等分子机制。
1.启动子启动子是转录因子、RNA聚合酶、蛋白质等分子参与的转录调控区域。
以轉錄因子增加启动子活性、RNA聚合酶聚到启动子等机制,促进外源基因的转录。
2.核糖体终止与脱落外源基因转录结束后,RNA聚合酶被肌肉卸载或者核糖体终止转录,RNA分子从RNA聚合酶上脱落,达到终止转录的目的。
3. RNA后转录调控RNA嵌合物和mRNA后调节会负责操纵RNA分子的最终结构和功能。
包括RNA剪接、RNA剪枝、RNA修饰等步骤,以改变RNA分子的结构和稳定性,进而影响外源基因的表达。
三、转录因子对外源基因表达的调节转录因子是外源基因转录调控的重要机制之一。
它会绑定到外源基因的启动子区域,增强或减少其转录活性,进而影响外源基因的表达。
目前,已经发现了多种调节类转录因子,可以在外源基因表达时起到重要作用。
其中,常见的转录因子包括激素受体、EGF受体、PPAR受体等。

《转基因阿尔巴斯白绒山羊外源基因沉默相关研究》篇一一、引言随着现代生物技术的迅猛发展,转基因动物作为基因编辑的优良载体,已经逐渐在畜牧养殖、生物医药、科研等多个领域发挥重要作用。
其中,转基因阿尔巴斯白绒山羊以其优秀的繁殖性能和优良的绒毛品质受到了广泛关注。
然而,在转基因过程中,外源基因的稳定表达及其沉默机制却成为了研究的关键问题。
本篇研究论文将深入探讨转基因阿尔巴斯白绒山羊中外源基因沉默的相关问题及其原因,旨在为今后的基因编辑及遗传育种工作提供理论基础和参考依据。
二、外源基因在阿尔巴斯白绒山羊中的表达转基因阿尔巴斯白绒山羊的培育过程中,外源基因的导入是关键步骤之一。
这些外源基因通常被整合到山羊的基因组中,并期望能够稳定表达以实现预期的生物学效应。
然而,在实际操作中,外源基因的表达往往受到多种因素的影响,包括基因本身的特性、整合位点的选择、宿主细胞的反应等。
三、外源基因沉默现象及其机制外源基因沉默是指转基因在宿主细胞中无法正常表达或表达水平显著降低的现象。
这一现象在转基因阿尔巴斯白绒山羊中同样存在,并可能由多种机制引起。
其中,最主要的机制包括甲基化修饰、非编码RNA的调控、宿主免疫系统的识别与清除等。
这些机制可能单独或共同作用,导致外源基因的表达沉默。
四、研究方法与实验设计为了研究转基因阿尔巴斯白绒山羊中外源基因沉默的原因及机制,我们设计了一系列实验。
首先,我们利用分子生物学技术手段,检测外源基因在山羊不同组织中的表达情况。
其次,通过分析基因组的甲基化水平,探讨甲基化修饰对外源基因表达的影响。
此外,我们还利用生物信息学方法,分析非编码RNA的分布及与外源基因的相互作用。
最后,通过建立免疫学模型,研究宿主免疫系统对转基因的识别与清除机制。
五、实验结果与分析经过一系列实验,我们得出了以下结论:1. 外源基因在阿尔巴斯白绒山羊的不同组织中存在表达差异,这可能与基因本身的特性及整合位点的选择有关。
2. 甲基化修饰是导致外源基因沉默的重要机制之一,且在不同组织中存在差异。



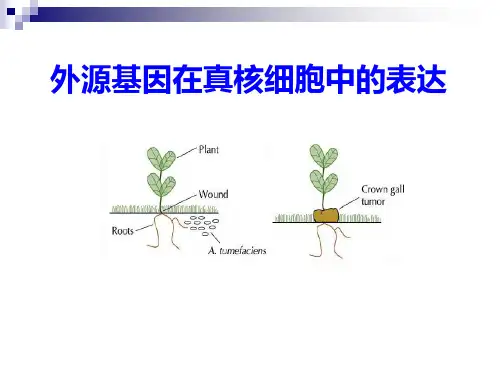
实验九外源基因在大肠杆菌中的诱导表达和降解物阻遏作用【实验目的】1.了解外源基因在原核细胞中表达的基础理论。
2.掌握乳糖操纵子的调节机制和操作方法。
【实验原理】1.外源基因在原核细胞中的表达蛋白质通常是研究的最终目标,因此蛋白质的表达在基因工程中占有非常重要的地位。
常用的表达系统有原核细胞和真核细胞。
原核细胞表达系统主要使用大肠杆菌,真核细胞表达系统主要有酵母细胞、哺乳动物细胞和昆虫细胞。
这些表达系统各有优缺点,应根据实验目的和实验室条件加以选择。
本实验主要介绍以大肠杆菌为代表的原核细胞表达系统。
(1)大肠杆菌表达系统的特点:生物学特性和遗传背景清楚,易于操作;已开发较多的克隆载体可供选择;容易获得大量的外源蛋白(外源蛋白可占细菌总蛋白50%左右)。
(2)蛋白质在原核细胞中的表达特点:原核细胞有其固有的RNA聚合酶,识别原核基因的启动子。
因此,在用原核细胞表达目的基因(无论是真核基因还是原核基因)时,一般应使用原核启动子。
原核基因的mRNA含有SD序列,启动蛋白质的合成。
而在真核基因上则缺乏该序列。
因此,一些商品化原核表达载体上设计有SD序列,以方便真核基因的表达。
原核细胞没有mRNA转录后加工的能力。
因此,在原核细胞中表达真核基因时,应使用cDNA 为目的基因。
原核细胞缺乏真核细胞对蛋白质进行翻译后加工的能力。
如表达产物的功能和蛋白质的糖基化、高级结构的正确折叠有关,必须慎重使用原核表达系统。
外源基因在大肠杆菌中高效表达时,表达产物往往在胞浆聚集,形成均一密度的包涵体。
包涵体的形成有利于保护表达产物不被胞内的蛋白酶降解,而且可以通过包涵体和胞内其他蛋白质密度不同来纯化包涵体蛋白。
但包涵体蛋白不具有该蛋白的所有生物学活性,往往需要通过变性复性的方法恢复活性,有时只能回复部分活性。
(3)蛋白质在原核细胞表达的调控启动子是转录水平调控的主要因素。
根据启动子起始mRNA合成效率的不同,可分为强、弱启动子,但是启动子的强弱是相对于不同基因而言的。


外源基因在植物种子中的表达与遗传稳定性研究基因编辑技术的兴起引发了对外源基因在植物种子中表达与遗传稳定性的研究。
外源基因可通过基因编辑技术、转基因等方式引入植物中,从而改变植物的性状。
然而,外源基因的遗传稳定性对植物的遗传改良和技术应用具有重要意义。
本文将从表达稳定性、遗传转移趋势两个方面综述外源基因在植物种子中的表达与遗传稳定性研究。
一、表达稳定性表达稳定性是指外源基因在植物种子中的表达程度是否稳定不变。
以水稻为例,Leone等人通过RT-qPCR测定外源基因Bar在各代间的表达水平差异。
结果表明,通过离子束转化的Bar基因在各个世代中表达相对稳定,但存在一定的浮动范围。
另一方面,通过农杆菌介导的转化方式表达的外源基因在各代间存在显著的表达变异。
这说明不同的转化方式对外源基因的稳定性有不同的影响。
表达稳定性还受到植物材料、生长条件、检测方法等多方面因素的影响。
例如,不同部位的种子含有不同程度的外源基因表达差异;温度、湿度等环境条件也会影响外源基因的表达。
此外,基因的检测方法和转录后调控机制也会对表达稳定性产生影响。
二、遗传转移趋势遗传转移趋势是指外源基因在不同世代间的遗传转移概率。
外源基因引入植物后,如果能够成功地遗传给后代,就可以达到遗传改良的目的。
研究表明,外源基因的遗传稳定性与植物的自然杂交、转基因品种与自然种群接触等因素密切相关。
特别是在转基因品种与自然种群接触时,外源基因的遗传转移率会更高。
然而,没有进一步的分析与控制,外源基因的转移概率可能过高,导致环境风险、生态安全等问题。
因此,控制外源基因在植物种子中的转移趋势是一个关键的问题。
有研究表明,结构稳定的转基因植物对外源基因的遗传稳定性更高。
建立基于DNA重组和基因编辑技术的高效分子标记技术,有助于对外源基因在植物中的遗传转移进行监控和控制。
结论外源基因在植物种子中的表达稳定性和遗传转移趋势是遗传改良和技术应用中的重要问题。
表达稳定性受到多种因素的影响,在转化方式、材料、环境、检测等方面的调控有助于提高表达稳定性。


遗传学实验引言遗传学是研究遗传原理和规律的科学,通过实验可以帮助我们更好地理解和应用遗传学的知识。
本文将介绍几个常见的遗传学实验,并详细讨论实验的步骤和结果。
实验一:显性遗传实验实验目的通过观察后代表现形状确定亲代基因表达方式。
实验步骤1.选取一对昆虫作为实验对象,确保它们具有不同的表现形状。
例如,可以选择黑色翅膀的昆虫A和白色翅膀的昆虫B。
2.让昆虫A和昆虫B进行交配。
3.观察并记录交配后代的表现形状。
实验结果根据观察结果,如果后代中出现了黑色翅膀的昆虫,说明黑色翅膀是昆虫A的显性基因;如果后代全是白色翅膀的昆虫,说明黑色翅膀是昆虫B的隐性基因。
实验二:基因突变实验实验目的检测和观察基因突变对个体表现的影响。
实验步骤1.选择一种含有某个基因的生物作为实验对象。
2.通过诱变剂处理生物体,诱发基因突变。
3.观察和记录突变个体与正常个体的差异。
实验结果根据观察结果,突变个体与正常个体在某些性状上会有明显的差异。
这些差异可以帮助我们了解基因的功能和作用。
实验三:基因型分析实验实验目的通过遗传标记和DNA分析来判断个体的基因型。
实验步骤1.提取个体的DNA样本。
2.选择适当的遗传标记进行PCR扩增。
3.将扩增产物进行电泳分析,观察带型。
4.与已知基因型的样本进行比对,判断个体的基因型。
实验结果通过电泳分析,我们可以得到个体的基因型。
这对于遗传研究和疾病诊断非常重要。
实验四:基因转导实验实验目的通过将外源基因导入细胞中,研究基因的功能和调控机制。
实验步骤1.选择目标细胞,如细菌或植物细胞。
2.构建外源基因的载体。
3.将载体导入目标细胞。
4.观察和记录导入细胞中外源基因的表达情况。
实验结果通过观察外源基因在目标细胞中的表达情况,我们可以了解基因的调控机制,并进一步应用于基因工程和农业生产。
结论遗传学实验是研究遗传学的重要手段,通过实验可以帮助我们更好地理解遗传原理和基因的功能。
本文介绍了显性遗传实验、基因突变实验、基因型分析实验和基因转导实验的步骤和结果。
外源基因在转基因生物中的遗传稳定性研究随着现代生物技术的飞速发展,转基因技术已经被广泛应用于食品、化妆品、医药以及农业等领域中。
转基因生物是指利用DNA重组技术将外源基因导入宿主细胞而形成的新生物。
外源基因的导入使得转基因生物具有了新的性状和特征,从而拓展了其应用领域。
然而,在实际应用中,外源基因在转基因生物中的遗传稳定性问题一直是一个备受关注和争议的话题。
外源基因在转基因生物中的遗传稳定性问题主要包括两个方面:一是外源基因在转基因生物中的表达稳定性问题,二是外源基因在转基因生物世代遗传稳定性问题。
首先,外源基因在转基因生物中的表达稳定性问题是指外源基因在转基因生物中是否能够持续地表达出其应该具有的功能。
外源基因的表达稳定性是转基因生物具有新特征的重要前提。
一旦外源基因的表达失效,转基因生物的特征也会受到影响。
因此,转基因生物外源基因的表达稳定性是一个十分关键的问题。
遗传稳定性是表达稳定性的一个重要方面。
遗传稳定性是指转基因生物在繁殖过程中是否能够保持其遗传信息的稳定性。
如果外源基因在转基因生物的繁殖过程中出现了不确定性的变异,那么将会影响其下一代的特征和应用效果。
因此,外源基因在转基因生物中的遗传稳定性问题对于转基因生物的应用安全和长期应用效果具有重要影响。
其次,外源基因在转基因生物世代遗传稳定性问题是指在转基因生物的繁殖和后代中,外源基因是否能够保持稳定。
这是由于在转基因生物的繁殖和后代中,外源基因可能会发生随机变异和重组,从而影响其稳定性。
如果外源基因在转基因生物的各个世代中出现了不确定性的遗传行为,那么将会影响其下一代的特征和应用效果。
为了探究外源基因在转基因生物中的遗传稳定性问题,许多科学家进行了一系列的研究。
有研究表明,在转基因植物的繁殖过程中,外源基因可能会出现随机变异和重组,从而影响其遗传稳定性。
这些突变可能会增加或降低外源基因的表达,或者引起新的表达或功能。
而有些研究,则提出了一些关于提高转基因生物遗传稳定性的方法。
外源DNA在细胞中的转移和表达随着科技的不断发展,我们对基因组的研究越来越深入。
在研究过程中,我们发现外源DNA的转移和表达对于生物的遗传特征和功能有着深远的影响。
本文将探讨外源DNA在细胞中的转移和表达的相关问题。
一、外源DNA的转移外源DNA指的是与受体生物个体基因组不同来源的DNA。
它主要通过三种方式转移:共轭转移、转化和病毒感染。
1.共轭转移共轭转移是指某些细菌通过细胞接触传递质粒来进行外源DNA的转移。
质粒是一种环形DNA分子,存在于细胞质中。
质粒常常拥有一个带有选择性抗性基因的繁殖元件,其可以使细胞对抗多种抗生素的攻击。
共轭转移是一种有目的的方式,它使得受体个体获得了不同的基因信息,从而增强了生命体对环境的适应能力。
2.转化转化是指细菌在自然环境中吸收外源DNA的过程。
在自然环境中,有许多细菌死亡,并释放出它们的DNA。
这些DNA被其他细菌吸收并整合到自己的基因组中。
这个过程被称为“自然转化”。
3.病毒感染有些病毒会把它们的DNA复制到细胞中。
这些病毒被称为“转座子”。
转座子能够整合到基因组的一个特定位置上,并且能够激活或关闭其他基因。
这种方式是一种被动的转移,但是病毒感染也可以改变个体的生物遗传特征。
二、外源DNA的表达外源DNA被转移到受体个体后,在细胞中,需要进一步经过转录和翻译等过程才能表现出来。
外源DNA在受体个体中的表达三种方式:转录、翻译和蛋白质修饰。
1.转录转录是指DNA信息通过RNA分子传递到蛋白质的生物合成过程。
外源DNA在受体个体中的转录,需要使用特殊的酶来“阅读”DNA序列。
这些酶负责转录DNA,把它们转化成类似于RNA的分子。
这种方式被称为“反转录”。
反转录发生在病毒、反转录病原体和外来基因的转染过程中。
2.翻译翻译是指RNA指导的蛋白质骨架的合成过程。
外源DNA被转录为mRNA后,mRNA要到核外才能被翻译成蛋白质。
在核外,mRNA被翻译成蛋白质,然后该蛋白质在细胞中被修改,最终形成目标蛋白质。