第六章 细胞内功能区隔 与蛋白质分选
- 格式:pdf
- 大小:6.29 MB
- 文档页数:98


第二章1.显微分辨率(microscopic resolution)---在必然条件下利用显微镜所能看到的精细程度。
3.双向凝胶电泳(two-dimensional electrophoresis)---依照分子质量及等电点的不同将复杂的蛋白质混合物分开。
这种高分辨率的技术能够分离同一混合物中的上千种蛋白质。
2.简述冷冻蚀刻术的原理和方式。
冷冻蚀刻(freeze-etching)技术是在冷冻断裂技术的基础上进展起来的更复杂的复型技术。
若是将冷冻断裂的样品的温度略微升高,让样品中的冰在真空中升华,而在表面上浮雕出细胞膜的超微结构。
当大量的冰升华以后,对浮雕表面进行铂一碳复型,并在侵蚀性溶液中除去生物材料,复型经重蒸水多次清洗后,捞在载网上作电镜观看。
5.比较差速离心和密度梯度离心。
都是利用离心力对细胞匀浆悬浮物中的颗粒进行分离的技术。
差速离心通经常使用于分离细胞器和较大的细胞碎片,分离的对象都比介质密度大。
密度梯度离心也可分离较大颗粒和细胞器,但更多用于分离小颗粒和大分子物质。
介质形成一个密度梯度,所分离的物质密度小于介质底物的密度。
6.在进行细胞组分的分离时,实验方案设计的一样原那么是什么?依照所分离的物质具有必然的体积和密度,通过离心力场的作用加以分离,依照这两个因素可设计速度离心、等密度离心(蔗糖CsCl)。
第三章1.模板组装(template assembly)-----指由模板指导,在一系列酶的作用下,合成新的、与模板完全相同的分子。
这是细胞内一种极为重要的组装方式,DNA和RNA的分子组装就属于此类。
2.酶效应组装(enzumatic assembly)---相同的单体分子在不同的酶系作用下,生成不同的产物。
3.自体组装(self assembly)---生物大分子借助本身的力量自行装配成高级结构,现代的概念应明白得为不需要模板和酶系的催化,以别于模板组装和酶效应组装。
4.细胞社会学(cell sociology)---细胞社会学是从系统论的观点动身,研究细胞整体个细胞群体中细胞间的社会行为(包括细胞间识别、通信、集合和彼此作用等),和整体和细胞群对细胞的生长、分化和死亡等活动的调剂操纵。



细胞生物学复习资料某些亲水分子或离子在通道间的流动沟通信息。
8.细胞外被---也称为细胞被,是细胞质膜中糖蛋白、蛋白聚糖、糖脂的寡聚糖链向外伸展,交织而成的一种绒毛状结构。
9.受体酪氨酸激酶---使酪氨酸磷酸化的膜受体类。
10.表面受体---位于细胞质膜上的受体称为表面受体。
11.细胞内受体---位于胞质溶胶、核基质中的受体称为细胞内受体。
12.表皮生长因子---表皮生长因子是一种小肽,53个氨基酸残基组成,与应答细胞表面的特异受体结合,一旦结合,变促进受体二聚化并使细胞质位点磷酸化。
13.GTP结合蛋白--- 包含两大类G蛋白,一类是与7次跨膜结构域超家族受体结合的异三聚体G蛋白,参与信号转导;另一类是小的胞质G蛋白。
二、简答题1.比较黏着斑和带连接的结构组成和功能。
粘着斑连接位于上皮细胞紧密连接的下方,依借粘着蛋白与肌动蛋白相互作用,将两个细胞连起来。
根本区别是:1)带是细胞与细胞之间的粘着连接;斑是细胞与细胞外基质进行连接。
2)参与带连接的膜整合蛋白是钙粘着蛋白,而参与斑连接的是整联蛋白,带是两细胞膜上的钙粘着蛋白之间连接。
斑是整联蛋白与胞外基质中的纤连蛋白连接。
因整联蛋白是纤连蛋白的受体,所以是受体与配体的结合所介导的。
2.比较黏着斑和半桥粒。
粘着斑和半桥粒这两种细胞粘着结构在不同的基膜上形成,粘着斑在体外将细胞结合在人工基膜上,而半桥粒在体内将细胞结合在基膜上。
结构上的差异是粘着斑与细胞内肌动蛋白纤维相关联,而半桥粒与细胞内的角蛋白纤维相关联。
3.说明间隙连接的结构特点和作用。
间隙连接存在于大多数动物组织。
在连接处相邻细胞间有2~4nm的缝隙,而且连接区域比紧密连接大得多,最大直径可达μm。
在间隙与两层质膜中有大量蛋白质颗粒,是构成间隙连接的基本单位,称连接子,6个相同或相似的跨膜蛋白亚单位环绕而成,直径8nm,中心形成一个直径约的孔道。
通过向细胞内注射分子量不同的染料,证明间隙连接的通道可以允许分子量小于的分子通过。

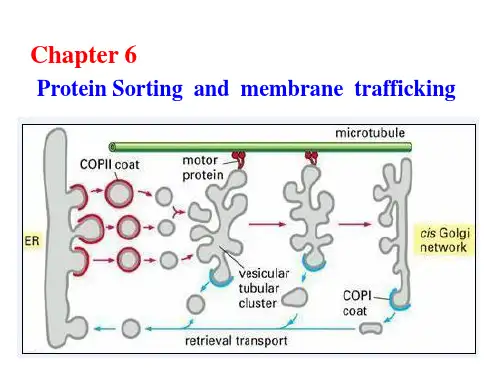

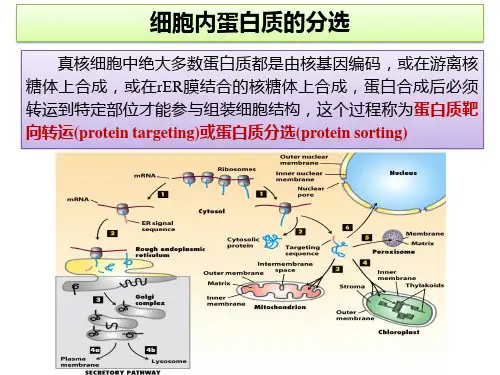
第一节蛋白质分选的基本原理第二节胞内膜泡运输第三节内质网第四节高尔基体第五节溶酶体第六节微体与原核细胞物不同的是真核细胞具有复杂的由内膜构成的功能区隔。
细胞内膜系统指在结构,功能或发生上相关的细胞内膜形成的细胞结构,包括核被膜、内质网、高尔基体及其形成的溶酶体和分泌泡等,以及其它细胞器如线粒体,质体和微体等膜包围的细胞器(膜性细胞器)。
内膜系统形成了一种胞内网络结构,其功能主要在于两个方面:其一是扩大膜的总面积,为酶提供附着的支架,如脂肪代谢、氧化磷酸化相关的酶都结合在细胞膜上。
其二是将细胞内部区分为不同的功能区域,保证各种生化反应所需的独特的环境。
本章主要介绍内质网、高尔基体、溶酶体、过氧化物酶体的功能和蛋白质分选,关于线粒体、叶绿体和细胞核的功能与蛋白质分选将分别在第七章(线粒体与叶绿体)和第十一章(细胞核与染色体)中讲解。
第一节蛋白质分选的基本原理从系统发生来看内膜系统起源于质膜的内陷和内共生(线粒体、叶绿体),从个体发生来看新细胞的内膜系统来源于原有内膜系统的分裂。
当细胞进行分裂时,不仅要进行染色体和细胞核的复制,同时各种细胞器通过吸收新合成的成分长大,然后随着细胞的分裂分配到子细胞中去。
细胞不能从无到有产生所有膜性细胞器,新的膜性细胞器来源于已存在细胞器的分裂。
如果彻底移除细胞内所有的过氧化物酶体,细胞根本不能重建新的过氧化物酶体,因为过氧化物酶体存在选择性地接受细胞质内合成的蛋白质的转位因子(translocator)。
细胞内合成的蛋白质、脂类等物质之所以能够定向的转运到特定的细胞器取决于两个方面:其一是蛋白质中包含特殊的信号序列(signal sequence or targeting sequence ),其二是细胞器上具特定的信号识别装置(分选受体,sorting receptor)。
因此内膜系统的发生具有核外遗传(epigenetic)的特性。
表6-1 一些典型的分选信号信号序列rg-Lys-Val-la-Gly-Leu-Asp-Ile-rg-Gln-Ser-Ile-Arg-Phe-Phe-Lys-Pro-Ala-Thr-Arg-Thr-Leu-Cys-Ser-Ser-Arg-Tyr-Leu-Leu-la-Met-Ala-Ser-Leu-Gln-Ser-Ser-Met-Ser-Ser-Leu-Ser-Leu-Ser-Ser-Asn-Ser-Phe-Leu-Gly-Gln-Pro-Leu-Ser-Pro-Ile-Thr-Leu-al-Ser-Leu-Leu-Leu-Val-Gly-Ile-Leu-Phe-Trp-Ala-Thr-Glu-Ala-Glu-Gln-Leu-Thr-Lys-Cys-Glu-Val-Phe-Gln-一、蛋白质分选信号细胞类至少存在两类蛋白质分选的信号:1.信号序列(signal sequence):存在于蛋白质一级结构上的线性序列,通常15-60个氨基酸残基,有些信号序列在完成蛋白质的定向转移后被信号肽酶(signal peptidase)切除.2.信号斑(signal patch):存在于完成折叠的蛋白质中,构成信号斑的信号序列之间可以不相邻,折叠在一起构成蛋白质分选的信号。