线粒体的形态结构.
- 格式:ppt
- 大小:1.42 MB
- 文档页数:31


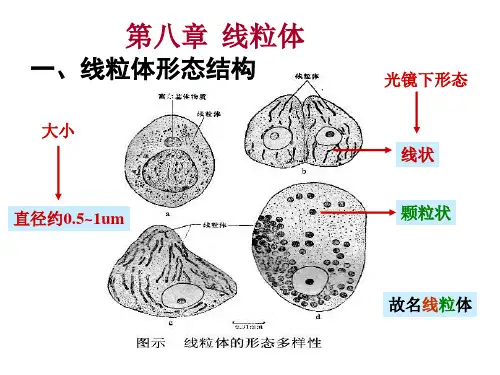
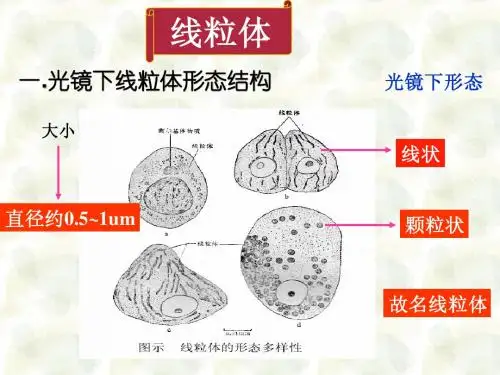
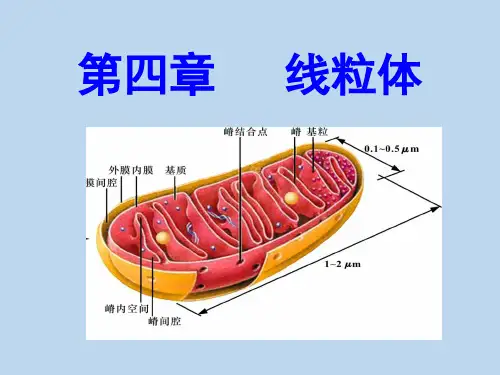
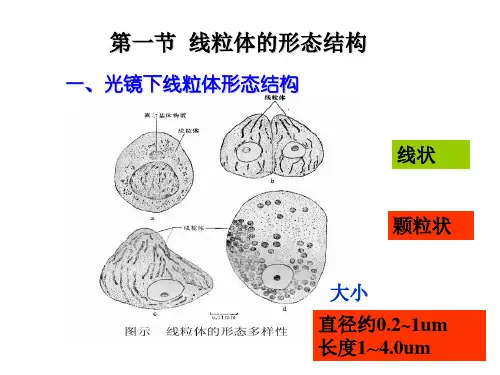
线粒体形态结构:椭圆形少数圆形宽0.8~1μm,长1~2μm(特殊7~40μm)两层单位膜外膜、内膜、外室、内室组成内膜向线粒体腔形成褶叠,称Mit嵴(嵴间平行,与纵轴垂直)嵴增加内膜面积(外膜的5倍,细胞膜结构总面积的1/3)基粒(F1颗粒、F1因子、内膜亚单位)外膜、内膜、外室、内室组成线粒体化学组成蛋白质、脂、水(脂:磷脂等膜脂;水:代谢产生)蛋白质最重要成份可溶性:基质酶、膜外周蛋白不溶性:内外膜整合蛋白(受体、酶等)蛋白质在线粒体各部分分布不同外膜两类:►(1)氧化还原酶:单胺氧化酶、NADH-细胞色素C还原酶等(单胺氧化酶是外膜标志酶)►(2)脂类代谢酶:酰基辅酶A合成酶、脂肪酸延伸酶等此外:孔蛋白,2~3nm通道,外膜具有通透性,小分子物质自由进出外室中酶种类少,主要为激酶,如腺苷酸激酶(标志酶)、二磷酸激酶等内膜上酶种类多、结构复杂、许多酶以复合物的形式出现内膜上的酶,四类:(1)移位酶:特异性载体,运输磷酸、Ca2+、核苷酸、α-酮戊二酸、谷氨酸、鸟氨酸等代谢产物(2)生物合成酶类:参与Mit DNA、RNA、蛋白质合成、脂肪酸、血红素合成(3)呼吸链酶:构成内膜的主要成份a.复合物I:NADH脱氢酶b.复合物II: 琥珀酸脱氢酶c.复合物III:CoQ-细胞色素C还原酶d.复合物IV:细胞色素氧化酶(4)能量转换酶——基粒(ATP酶复合体)♦基粒:ATP合成酶(F1-F0偶联因子,F1-F0 ATP酶)三部分:头部:球状,直径9nm,容易分离,水溶性,称F1因子,由α3β3εδγ 9亚基组成►分离的F1因子能催化:ADP+Pi → ATP也能催化:ATP → ADP+Pi►但Mit内,合成反应,存在一种热稳定蛋白(MW1万),可与F1结合,抑制ATP水解,促进合成►称F1抑制蛋白柄部:►由一种蛋白质组成►高4.5nm,容易分离►作用:使F1因子对寡霉素敏感►F1因子合成ATP的活性能被寡霉素抑制,但寡霉素不直接作用于F1因子,通过柄部蛋白传递►称寡霉素敏感性转授蛋白(oligomycin-sensitivity confering protein,OSCP)膜部:►嵌合在膜内疏水蛋白复合体,称F0因子►至少由3~4条肽链组成►其中一种与寡霉素结合,通过OSCP对F1因子起作用►与质子H+传递有关►当去除F1 和OSCP,内膜对H+的通透性↑,结合上去,可堵住H+泄漏,推测有H+通道线粒体ATP的合成►ATP合成是线粒体最重要功能►ATP来源于糖、氨基酸、脂肪酸的氧化►三类物质存在共同的氧化途径:三羧酸循环ATP合成几种假说:►化学偶联假说►构象偶联假说►化学渗透假说(1978年诺贝尔奖)又称电化学偶联假说基本观点:呼吸链在电子传递中,起着质子泵的作用从NADH脱氢酶开始,每传递一对电子(2e),将线粒体基质的3对质子(6H+)泵到外室(从琥珀酸脱氢酶开始?)化学渗透假说►随着电子传递不断进行,内膜内外形成[H+]浓度差和电位差►电化学梯度(势能)►外室[H+]有返回基质的趋势►每一对质子,驱动合成一个ATP实验证明:微电极,内膜两侧存在电位和pH梯度缬氨酶素,内膜通透性↑,[H+]梯度消失,电子传递进行,但ATP 合成停止►ATP合成酶工作机制►分子“马达”►旋转催化机制►γ亚基像车轮的“轴”,在质子流推动下,转动►带动与“轴”相连的3个β亚基转动(逆时针)►导致β亚基构象交替变化(三种构象:L 结合ADP+Pi;T 合成ATP;O 释放ATP,并周而复始)►1997年,诺贝尔化学奖线粒体的增殖和起源进化(1)增殖分割或分裂产生(2)起源进化内共生假说细菌被原始真核细胞吞噬,演变成线粒体(叶绿体蓝藻)非共生假说共生假说证据:Mit内膜与细菌质膜相似,表现在:►都有呼吸链酶►细菌质膜可以向细胞质内延伸出类似于线粒体嵴的结构(中体)Mit外膜与真核细胞质膜相似(内吞时质膜包围)►Mit DNA与细菌DNA相似:双链环状,复制方式和过程,分子量,裸露►核糖体相似:沉降系数70S左右,小亚基16S rRNA核苷酸序列相似,蛋白质合成过程相似,各种抗生素抑制抗生素抑制细菌生长机理:抑制70S核糖体(不抑制80S核糖体)。