植物抗寒性产品研究
- 格式:ppt
- 大小:5.16 MB
- 文档页数:67

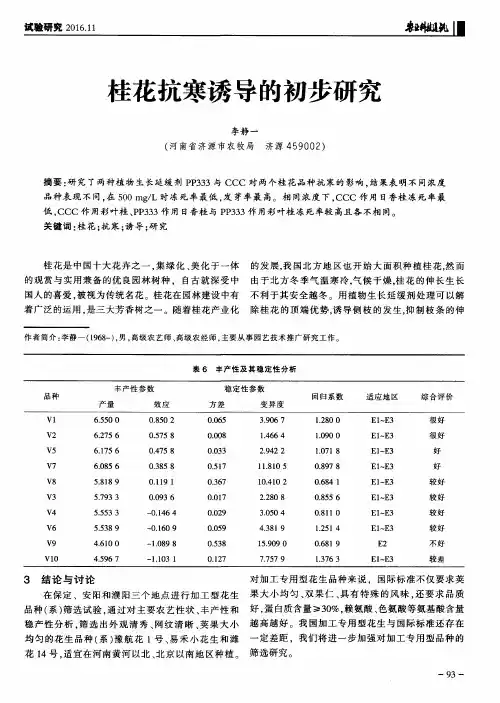
不同葡萄品种抗寒性比较研究作者:高军辉等来源:《安徽农业科学》2014年第13期摘要 [目的]鉴定新疆地区葡萄(Vitis vinifera L.)品种的抗寒力,对于品种资源的利用、品种的合理规划布局和抗寒育种及评价、推广抗寒性品种都具有重要的意义。
[方法]以3个葡萄品种(红地球、里扎马特、无核紫)为试材,研究低温处理(-23、-4 ℃)对葡萄枝条相对电导率、可溶性糖含量、束缚水/自由水的影响。
[结果]随着处理温度的降低,各品种枝条的相对电导率、可溶性糖、束缚水/自由水含量逐渐升高,但是各品种升降的速度和幅度有差异,各品种参数的升降幅度大小顺序与其抗寒性强弱顺序基本相符。
[结论]检测的3个品种中,无核紫的抗寒性最强,红地球的抗寒性最弱。
关键词葡萄品种;抗寒性;新疆地区中图分类号 S663.1 文献标识码 A 文章编号 0517-6611(2014)13-03830-02Abstract [Objective] The aim was to identify different grapevine cultivars of Xinjiang area,which had important significance for cultivars resources utilization, cultivars reasonable layout,breeding cultivars with cold resistance, and evaluation and promotion the cultivars with cold resistance. [Method] Taking three grapevine cultivars (Red Globe, Rizamat, Monukka)as materials, the content of soluble sugar, relative conductivity and irreducible water/free water of grapevine branches under low temperature(-23, -4 ℃) treatments were studied. [Result] As the temperature decreasing, the relative conductivity, soluble sugar and the ratio of irreducible water to free water of three grapevine cultivars branches increased gradually, while the change speed and range were different among cultivars, the order of cultivars parameters were consistent with the cold resistance. [Conclusion] In three grapevine cultivars, the resistance to cold of Monukka was the most, while that of Red Globe was the weakest.Key words Grapevine cultivars; Cold resistance; Xinjiang area葡萄(Vitis vinifera L.)属于葡萄科葡萄属(Vitis)[1]。

经济林中山杏抗寒性能研究【摘要】:用3种山杏的花器官为材料,进行低温胁迫,并分析3种类型的抗寒性差异。
潮定各类型的自由水含量与束缚水含量的比值,分析统计数据并进行抗寒性分析,比较各类型的抗寒性差异,总结出一定的规律,并进行分析,提出建议。
【关键词】:山杏;抗寒性能山杏抗干旱、耐瘠薄,具有优良的水土保持和防风固沙功能,其经济价值高,杏仁含多种营养成分,应用于食品、医药及工业等领域。
随着植物生理学的发展,越来越多的人从生理生化指标方面开展植物抗寒性研究,在山杏方面也有一些相关研究报道。
在低温胁迫下甜仁山杏半致死温度低于大扁杏、苦仁山杏,丙二醛含量增加速率较慢。
随干旱胁迫的加重,山杏幼根、叶片中的淀粉含量大幅度减少。
SOD、POD等酶促系统、脯氨酸含量和可溶性蛋白可作为山杏花受冻的检测指标。
1、材料和方法1.1材料调查中选取形态特征趋向于抗性强的20~30年生山杏树(设为C型)和抗性较弱的类型(设为B型)。
另外再选取两种类型的中间类型(设为A型)。
即形态特征介于上述两种类型之间的中间类型。
山杏树冠周围随机采取正常生长的一年生枝剪取10-20cm的大蕾期枝条200个。
在实验室用5%的蔗糖溶液进行培养.待盛花期进行处理。
1.2方法1.2.1组织含水量的测定取称量瓶3只(3次重复),依次编号并准确称重.将待测山杏花器官摘取50只花瓣立即装入瓶中。
盖上瓶盖并准确称重。
将称量瓶置烘箱中110℃下15min然后再于80-90℃下烘至恒重。
设称量瓶重为W1,称量瓶与花瓣的重量为W2,称量瓶与烘干花瓣的重量为W3,花器官组织含水量=(W2-W3)/(W2-W1)×100%分别求出3次重复所得到的花器官组织含水量的值.并求平均值。
按上述方法依次求出3种类型山杏花器官的组织含水量。
1.2.2自由水与束缚水值的测定取称量瓶3只(3次重复),依次编号并准确称重。
将待测山杏花器官摘取50只花瓣立即装入瓶中。
盖上瓶盖并准确称重。

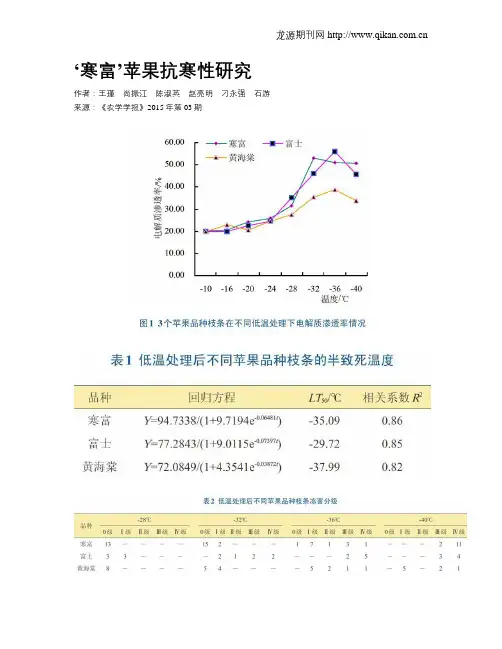
‘寒富’苹果抗寒性研究作者:王瑾尚振江陈淑英赵亮明刁永强石游来源:《农学学报》2015年第03期摘要:为研究‘寒富’苹果的抗寒能力及其在伊犁河谷适宜栽培的区域,以苹果1年生枝条作为试验材料,利用电导法对3个不同品种的苹果休眠枝条在不同低温处理下的电解质渗透率进行测定,并配合Logistic方程确定苹果枝条的低温半致死温度。
结果表明,随着处理温度的下降,苹果枝条组织的电解质渗透率变化幅度呈现“慢一快一慢”的趋势,即“S”形曲线增长。
‘寒富’苹果的半致死温度(/Ts。
)在一350C左右,对照苹果普通‘富士’的半致死温度(LT50)在-290C左右,‘黄海棠’的半致死温度(LT50)在-38℃左右。
结合冻害分级情况,‘寒富’在伊犁州的逆温带可安全越冬,部分平原区也可栽植。
关键词:‘寒富’;电解质渗透率;半致死温度;抗寒性中图分类号:S661.1文献标志码:A论文编号:2014-03 700 引言‘寒富’是沈阳农业大学于1978年用‘东光’ב富士’杂交选育的苹果优良新品种,1998年通过辽宁省品种审定委员会审定。
‘寒富’表现出抗寒、丰产、果实品质优、短枝结果性状明显的特点,2006年春,伊犁州林科院从辽宁省果树研究所引进该品种,通过区域试验,‘寒富’表现出较强的抗寒能力及结实性。
2008年秋伊犁州林科院从辽宁省果树研究所引进‘寒富’苹果接穗,2009年春在野苹果砧木上嫁接,对其田间抗寒能力进行跟踪观察。
本研究以‘寒富’苹果1年牛休眠枝条作为试验材料,选择本地的普通‘富士’和‘黄海棠’作为参照,分别研究其在不同低温处理条件下相对电导率变化规律及抗寒能力。
电解质渗透率测定作为一种植物抗寒性鉴定方法,已广泛的使用在杏、野牛欧洲李等多种植物树体抗寒测定过程中,这种方法简便、快速、灵敏。
Sukumaran等曾提出以50%电解质渗出率相对应的温度作为植株的半致死温度,但在实际应用中存在较大的误差。
因为在不同低温胁迫下,植株的电解质渗透率值与处理低温不是直线相关,而呈现出符合Logistic方程的“S”型曲线相关,求出拐点值,此时拐点的温度则是引起膜不可逆损伤的临界值,即低温半致死温度/Tso,通过半致死温度的估计值,可对植物的抗冻能力进行更深一步了解。


不同植物种抗寒性测定——过氧化氢酶(CAT)活性的测定一、实验目的:1.学习并掌握实验室间接鉴定植物抗寒性的方法和步骤。
2.测定出三种植物的抗寒性能力强弱顺序。
3.通过本实验的设计与实施,增加我们对植物抗寒性的了解,有利于我们对植物的种植与保护,也能使我们掌握基本的实验操作技术,培养探究性学习的能力,通过小组合作学习加强团队精神二、实验原理:低温胁迫下,细胞内易产生H2O2破坏膜系统的稳定,而过氧化氢酶(CAT)能把H2O2分解为H2O和O2,从而清除H2O2维护膜的稳定性。
研究表明,抗寒性强的品种有较高的过氧化氢酶活性,且随温度下降,以抗寒性强的品种下降幅度小,与抗寒性呈显著性相关。
CAT酶活性大小可用一定时间内分解的H2O2量来表示。
在反应系统中加入一定量(反应过量)的H2O2溶液,经酶促反应后,用标准高锰酸钾溶液(在酸性条件下)滴定多余的H2O2,即可求出被CAT分解的H2O2量:三、材料及仪器:1.材料及处理:九里香、小叶罗汉松、圆柏将采回的枝条剪成40cm左右的长度,用自来水冲洗数遍(洗掉泥土、灰尘、虫卵),再用蒸馏水冲洗三次,然后用吸水纸吸干水分,最后将枝条末端进行蜡封。
将每个品种蜡封后的枝条分成相等的3份,在 0℃,6℃,12℃的恒温培养箱中各放入一份培养5天,备用。
2.试剂、药品:10%H2SO4 ,0.2mol/L 磷酸缓冲液(pH=7.8),KMnO4 (AR),新煮沸冷却蒸馏水,0.1mol/L草酸,30%H2O2溶液,凡士林3.仪器:25ml容量瓶,研钵,50ml三角瓶4个,酸式滴定管,1000ml 锥形瓶,离心管,移液管(0.5ml和5ml),滴管,剪刀4.设备:高速台式离心机,恒温水浴锅,天平,玻璃棒,石英砂,恒温培养箱四、实验步骤:实验前的准备阶段,1.试剂配制10%H2SO4;0.2mol/L 磷酸缓冲液(pH=7.8);0.1mol/L 高锰酸钾标准液:称取KMnO4(AR)3.160g,用新煮沸冷却蒸馏水配制成1000ml,用0.1mol/L草酸标定;0.1mol/L H2O2:取30%H2O2溶液5.68ml,稀释至1000ml,用标准0.1mol/L KMnO4溶液在酸性条件下标定。


植物抗寒性的基础研究及其应用植物是地球上最重要的生物之一,它们可以从阳光、水和空气中获取必要的营养来支持生命的维持。
然而,在极端的气候条件下,植物可能需要预测或应对严寒的环境问题,以确保生存。
为了适应这种环境条件,许多植物已经发展出了各种抗寒性机制,从而使它们能够在寒冷的气候中生存和繁衍。
近年来,植物抗寒的基础研究已经取得了重大进展,并为生产农业作物、生态恢复和全球气候变化适应等多个领域提供了重要的指导意义。
一、植物抗寒性的基础研究进展植物在寒冷条件下的生存需要与环境进行适应和转化,从而保持各种生理过程在正常的状态下进行。
在植物抗寒的基础研究领域,主要包括以下几个方面:(一)生理过程许多植物抗寒机制的研究是建立在对冷处理后植物生理响应的分析研究基础之上的。
植物在寒冷环境中能够适应和调节的生理过程非常重要,如气体交换、光合作用、脱水和氮代谢等生理过程,研究这些过程有助于我们了解植物在不同温度下的适应机制。
(二)基因组学随着生物技术的发展,基因组学的研究方法逐渐成为植物抗寒的研究重要手段。
通过定位和克隆与植物抗寒相关的基因,可以研究植物在寒冷条件下的相关分子机制。
(三)代谢组学代谢组学是将代谢产物作为分析对象进行研究的手段。
它可以为我们提供真实的生物反应信息,帮助我们了解植物在环境变化下相应的代谢情况,以及相关代谢过程。
二、植物抗寒性的应用植物抗寒性的研究除了拓展我们对植物生命过程的认识以外,还在实践中得到了广泛的应用。
对此,我们可以从以下几个方面来进行阐述:(一)农业生产领域植物抗寒性研究对提高农作物产量、改良耐寒性作物品种等具有十分重要的意义。
研究团队可以通过转移和改变植物抗寒相关基因,开发出更加适应寒冷环境下栽培的农业作物品种;或者通过提高土壤氮含量等手段,提高植物的氮素代谢,增加其对冷度的适应能力。
(二)环境恢复领域在环境修复中,植物抗寒性也发挥出了重要作用。
例如,在恢复受水污染土地的过程中,适应寒冷环境范围内的抗寒性植物作为生长环境机制可以帮助加速该土地环境的修复速度。

库尔勒香梨是新疆的名、优、特色水果之一,近年来已相继选育出新梨1号、2号、3号、4号、6号和7号等优良品种,在国内外市场享有较高声誉,已成为重要的出口创汇品种之一。
但2003年以来的几次冻害,使库尔勒香梨安全越冬受到了严重影响。
安全越冬已成为库尔勒香梨产业可持续发展的重要制约因子。
库尔勒香梨属蔷薇科,梨亚科,梨属中的白梨系统,原产新疆库尔勒地区,商品名称有香梨、新疆香梨、中华香梨、香蜜梨等,是一古老的地方品种,已有1300多年的栽培历史。
香梨在新疆南部各地均有分布,以库尔勒地区最集中,北疆伊犁、东疆吐鲁番市也有栽培。
我国其他北方省市亦有少量试种。
库尔勒香梨植株生长势强,萌芽力中等,发枝力强,抗寒性较强。
果实大小适中,香味浓郁,皮薄肉脆,清甜爽口,细嫩多汁,在国际市场上有“中华蜜梨”“梨中珍品”“果中之王”“梨后”“梨中王子”等美誉。
1果树的抗寒性研究1.1形态解剖指标的研究果树的抗寒力受着诸多因素的影响,树种或树体部位的不同,组织细胞的生理特性和排列结构不同,其抗寒力也不尽相同,且受冻后的表现也存在差异。
一般芽眼大及生长旺盛的树种抗寒力相对较弱;果树叶片的紧密组织数与抗寒性呈正相关,且栅栏组织细胞小而细长的抗寒力较强。
1.2组织器官方面的研究果树抗寒性在枝条、花芽、形成层、韧皮部和皮孔大小等组织、器官上的表现常作为低温锻炼下的直接形态指标。
低温能使苹果枝条皮层细胞出现质壁分离现象,且抗寒性强弱顺序为:韧皮部>木质部>形成层>髓,果树各个器官之间抗寒性一般为:枝条>叶芽>花芽>蕾期>盛花期>幼果期。
1.3生理生化指标的研究1.3.1生理指标与抗寒性的研究低温条件下会引起不抗寒果树细胞理化特性发生变化。
植物受到低温影响时,胞的质膜透性会发生不同程度的增大,电解质外渗,以致电导率增大。
抗寒性强的品种细胞透性增大程度较小,且透性的变化可以逆转,易于恢复正常。
反之,不抗寒品种的细胞透性增加且不可逆转,以致造成伤害甚至死亡。

黑龙江农业科学2023(11):83-88H e i l o n g j i a n g A gr i c u l t u r a l S c i e n c e s h t t p ://h l j n y k x .h a a s e p.c n D O I :10.11942/j.i s s n 1002-2767.2023.11.0083叶丽努尔㊃哈德勒,刘璇,王玉萍,等.不同类型天山樱桃的抗寒性初步研究[J ].黑龙江农业科学,2023(11):83-88.不同类型天山樱桃的抗寒性初步研究叶丽努尔㊃哈德勒,刘 璇,王玉萍,满 好,周 龙(新疆农业大学园艺学院,新疆乌鲁木齐830052)摘要:为促进天山樱桃的进一步开发利用,以矮生和乔化两种类型天山樱桃的一年生枝条为试验材料,通过人工模拟低温胁迫处理(-20,-25,-30,-35和-40ħ),测定其相对电导率㊁可溶性糖㊁可溶性蛋白㊁脯氨酸及丙二醛含量等生理指标,以相对电导率结合L o g i s t i c 方程测算半致死温度(L T 50),进一步通过组织褐变法和恢复生长法,综合评价各参数与耐寒度间的关系,从而评价不同类型天山樱桃的抗寒性㊂结果表明,随着低温胁迫强度的增加,枝条的电导率㊁可溶性糖含量明显升高,当温度达到-40ħ时,两者的相对电导率均超过50%,矮生型天山樱桃枝条的半致死温度为-36.81ħ,乔木状天山樱桃枝条的半致死温度为-39.91ħ,可见两种类型天山樱桃的抗寒性顺序依次为乔木状天山樱桃>矮生型天山樱桃㊂关键词:天山樱桃;低温胁迫;抗寒性;生理指标收稿日期:2023-05-21基金项目:自治区高校基本科研业务费项目(X J E D U 2022-Z 004);自治区大学生创新创业项目(S 202310758063)㊂第一作者:叶丽努尔㊃哈德勒(2001-),女,本科生,从事果树抗寒生理研究㊂E -m a i l :1161589133@q q .c o m ㊂通信作者:周龙(1976-),男,博士,教授,从事果树种质资源和栽培生理方面的研究㊂E -m a i l :z h o u l o n g2004@126.c o m ㊂ 近年来,我国樱桃产业发展迅猛,其中甜樱桃因其果实外观鲜艳㊁口感较佳,而且富含维生素㊁蛋白质和钙㊁磷㊁铁等各种微量元素,因此营养价值也较高,而深受消费者的青睐[1]㊂但是由于甜樱桃抗寒性差,所以在我国的分布范围窄,集中分布在秦岭淮河以南的区域㊂为了扩大甜樱桃的栽培区,生产上多采用抗寒性强的砧木材料㊂天山樱桃作为新疆野果林特有的野生果树,具有极强的抗旱㊁抗寒以及抗病虫害能力,可作为李属及樱亚属砧木材料[2]㊂目前,对天山樱桃的研究工作仍然重点聚焦于花粉萌发特征[3]㊁遗传和表型多样性[4]㊁群落特征[5]等方面,而有关天山樱桃生态适应性的研究甚少,进一步明确天山樱桃的生态适应性,对于野生种质资源的开发利用有很高的科研价值和实践意义[6]㊂温度是天气因素中改变植物地理分布的主要原因,温度胁迫是植物栽培中常常出现的一个气候灾害,会影响到各种经济作物及其它多种植物,温度会影响植株的生长代谢,导致植株的生理指数发生改变,造成植株的生长发育受到一定的损害,严重的可致死[7]㊂相对电导率的改变能够反映植物细菌细胞壁的破坏程度,是能够反映各种树木抗逆能力的指数,同时也用作耐寒能力强弱的一个指数应用㊂根据相对电导率结合L o gi s t i c 方程推算某种植物的半致死温度,是果树抗寒性研究必不可少的方法[8]㊂李纪元等[9]发现电导法在评价黑木相思和灰木相思的抗寒性上有很好的准确性㊂何开跃等[10]通过对福建柏山低温胁迫的长期观察证实,M D A 浓度随着温度的降低而逐步增加㊂严寒静等[11]对栀子的调查研究发现,当温度逐渐下降时栀子叶内可溶性糖含量逐月增加,在温度最低的一月其可溶性糖含量是温度最高的九月的1.6倍㊂高志红等[12]对果梅的研究发现,低温胁迫下其枝条内可溶性蛋白含量和可溶性糖含量与抗寒性呈明显正相关,可用作检测种质抗寒性的重要指标㊂姜卫兵等[13]对无花果的长期研究表明,耐寒力越高的品种游离脯氨酸含量越是处于更高水平,而耐寒力较弱的种类则处在较低水平㊂S u k u m a r a n 等[14]提议以相对电导率为50%时的环境温度作为组织半致死温度,而近年来专家们普遍认为半致死温度(L T 50)可作为衡量某个物种抗寒性强弱比较精确的指数[15]㊂而耐寒能力也是植物最主要的抗性能力之一,当植物遭受逆境胁迫时,其生态改变过程是相当复杂的,且受到了许多因子的复合作用,因此仅仅依靠一两个指标很难去反映植物抗寒性的真实情况㊂不同类型天山樱桃的抗寒性具有差异,需对比分析乔木状天山樱桃和矮生型天山樱桃两种类型的抗寒性差异㊂天山樱桃为野生果树,作为不同类型天山樱桃在野外生境不同的情况下枝条发育情况㊁营养状态均不相同,如何准确进行评价是本项目研究的关键㊂本研究用多个指标对两种类型的天山樱桃的抗寒力进行研究,更具科学性和可靠性[16]㊂拟通过野外采样和试验基地采样相结合,室内人工模拟低温研究和室38黑 龙 江 农 业 科 学11期外自然越冬相结合的方式,拟合外部形态指标㊁生理指标等全面进行探讨和研究,综合予以评价㊂1 材料与方法1.1 材料在西天山野果林采集野外乔木状天山樱桃和矮生型天山樱桃的一年生休眠枝,在新疆农业大学园艺学院干旱区特色果树中心重点实验室进行材料处理㊂在实验室中,吸干表面的水分,存于4ħ冰箱备用㊂采用人工冷冻降温的方法,放置在冰箱中的温度控制范围是-20,-25,-30,-35和-40ħ,而每个温度的持续时间约为12h ㊂1.2 方法1.2.1 组织褐变观察 参照牛立新等[17]的研究结果㊂每个温度下选择3条洁净的一年生枝条,采用刀片横切成0.2c m 厚的薄片,并放置在解剖镜下观测枝条横切面的受冻变褐情况㊂采用褐变部分所占比例的方法,测定褐变面积㊂褐变面积(%)=(S 1/S 2)ˑ1001.2.2 相对电导率的测定 参照王文举等[18]的方法,将低温处理下的枝条采用刀片横切成0.2c m厚的薄片,称取0.3g ,装在45m L 的离心管中,再加入20m L 去离子水溶液,室温条件下封口浸提20h ,用D D S -307A 输出电导仪测出最初电导值C 1,用蒸馏水对照电导值C 0,将其放在煮沸浴中30m i n ,冷至常温后再测出最终的电导值C 2㊂如此反复计算3次,取平均值㊂相对电导率(%)=(C 1-C 0/C 2-C 0)ˑ1001.2.3 半致死温度的测定 根据苏李维等[19]的方法,用L o gi s t i c 方程组加以拟合环境温度(t )和相对电导率(y ),求出相对电导率的拐点温度,即为低温半致死温度(L T 50)㊂L o g i s t i c 方程:y =k /(1+a e -b t)1.2.4 生理指标的测定 参照武汉伊莱瑞特生物公司相关试剂盒使用说明测定其可溶性糖㊁可溶性蛋白㊁游离脯氨酸(P r o )㊁丙二醛(M D A )含量㊂可溶性葡萄糖含量的测定:称取0.1g 低温胁迫处理后的植物组织块,剪碎后加入乙醇并匀浆,后倒入有盖离心管中,再用80%乙醇冲洗研钵并转移到同一E P 管中,终体积定容为1.5m L ,50ħ水浴20m i n ,封口膜缠紧,且间隔2m i n 振荡混匀一次,冷却后12000r ㊃m i n -1离心10m i n,取上清液备用㊂在E P 管中依次加入0.025m L 样本㊁0.075m L 蒸馏水和0.030m L 工作液,再缓慢加入0.250m L 浓硫酸作为测定管,在E P 管中依次加入0.100m L 蒸馏水㊁0.030m L 工作液和0.025m L 浓硫酸作为空白管,混匀后,放入沸水浴中10m i n ,冷却至室温后,取200μL 转移至96孔板中,于620n m 读取吸光值㊂ΔA=A 测定-A 空白可溶性糖含量(m g ㊃g -1)=[(ΔA+0.0103)ː2.0899ˑV 1]ː(W ˑV 1ːV )ˑD =0.718ˑ(ΔA+0.0103)ːW ˑD可溶性蛋白质浓度(C pr )的测定:称取0.1g 低温胁迫处理的植物组织块,加入提取液后沐浴匀浆,12000r ㊃m i n -1离心10m i n,取上清㊂在E P 管中加入0.005m L 样本,混匀,于60ħ保温30m i n ,全部转移到96孔板,于562n m 处测定吸光值㊂ΔA=A 测定-A 空白C pr 鲜重(m g ㊃g -1)=[(ΔA+0.0046)ː0.188ˑ0.01]ː(V 1ːV ˑD )=1.06ˑ(ΔA+0.0046)ːW ˑD脯氨酸含量的测定:称取0.1g 低温胁迫处理后的植物组织块,剪碎后加入1m L 的提取液,匀浆后离心15m i n ,取上清用提取液稀释10倍,取稀释后的样本㊂取2m L 提取液,加入10m L试管中作为空白管㊂取2m L10μg㊃m L -1标准品,加入10m L 试管中作为标准管,取2m L 待测样本,加入10m L 试管中作为测定管,混匀,沸水浴30m i n ,流水冷却,加4m L 甲苯,振荡,静置10m i n,取上清液至离心管中后离心10m i n,吸取上层红色甲苯溶液,于520n m 处测定各管吸光度㊂脯氨酸含量(μg ㊃g -1)=(ΔA 1ːΔA 2)ˑc ˑV ːW ˑf丙二醛含量(M D A )的测定:称取0.1g 低温胁迫处理后的植物组织块,剪碎后加入提取液,沐浴匀浆后离心15m i n ,取上清液0.1m L ㊂取8个不同浓度的标准品各0.1m L ,分别加入对应的1.5m LE P 管中作为标准管,取0.1m L 待测样本,加入1.5m LE P 管中作为测定管,再向各管中加入0.6m L 工作液,混匀后E P 管口用保鲜膜扎紧,并扎一个小孔,95ħ以上水浴40m i n,取出后流水冷却,2000g 离心10m i n ㊂取0.25m L上清液加入酶标板,532n m ,测定O D 值㊂M D A (n m o L ㊃g -1)=(ΔA 532-b )ːa ˑf ː(m ːv )式中:b 为标曲的截距;a 为标曲的斜率;f 为样本加入检测体系之前的稀释倍数;m 为植物组织样本质量(g);v 为样本处理过程中加入试剂五抽提应用液的体积(m L )㊂1.2.5 恢复生长情况调查 取不同低温处理采集的天山樱桃枝条样品,分别放置在装有三分之二蒸馏水的150m m 的三角瓶密封后,存放于约4811期 叶丽努尔㊃哈德勒等:不同类型天山樱桃的抗寒性初步研究25ħ的室内,进行光照处理,每3d 换水1次,换水时修剪掉枝条基部便于更好地吸收水分,每天记录1次芽的萌发情况,统计萌芽率㊂1.2.6 数据分析 试验数据采用E x c e l 2019进行计算和作图,并用O r i g i n2021软件对L o gi s t i c 方程y =k /(1+a e -b t)进行拟合,若拟合度(r)显著时,再计算L T 50,从而分析判断温度和相对电导率之间的关系,最后综合比较分析两种类型天山樱桃的抗寒性㊂2 结果与分析2.1 不同类型天山樱桃的相对电导率和半致死温度由图1可知,两种类型天山樱桃枝条相对电导率随着处理温度的降低均呈现缓慢上升的趋势,且矮生型天山樱桃始终显著高于乔木状天山樱桃,并在-40ħ时达到最大值㊂其中矮生型天山樱桃相对电导率在-40~-30ħ时上升幅度最大,乔木状天山樱桃相对电导率在-40~-35ħ时上升的幅度最大㊂在相同温度下,矮生型天山樱桃的相对电导率均大于乔木状天山樱桃的相对电导率,在-35~-20ħ时,矮生型天山樱桃和乔木状天山樱桃的相对电导率均在50%以下,当温度达到-40ħ时,两者的相对电导率均超过50%㊂根据在不同温度处理下测定的两种类型天山樱桃的相对电导率求出其L o g i s t i c 方程㊁拟合度及低温半致死温度,结果如表1所示,不同温度胁迫下,乔木状的天山樱桃半致死温度为-39.91ħ,而矮生型天山樱桃休眠枝条的半致死温度为-36.81ħ,两种类型天山樱桃的方程相关系数分别为0.94269和0.95422㊂图1 低温胁迫下天山樱桃枝条相对电导率的变化表1 低温胁迫下的天山樱桃枝条半致死温度(L T 50)品种L o gi s t i c 方程拟合度L T 50/ħab矮生型天山樱桃Y =100/(1+4.44356e -0.04052t )0.95422-36.814.443560.04052乔木状天山樱桃Y =100/(1+5.22048e-0.04141t )0.94269-39.915.220480.041412.2 不同类型天山樱桃的组织褐变不同类型天山樱桃的休眠枝条对低温胁迫反应后的横切面的褐变现象如图2所示㊂枝条木质部㊁韧皮部及髓部在-30~-20ħ处理时颜色鲜绿,说明尚未经受到低温的胁迫,随着处理温度的进一步下降,两个类型天山樱桃枝条的组织褐变程度渐重,尤其是在木质部表现明显,出现褐色,处理温度-40ħ时木质部㊁韧皮部出现大面积的褐变现象,而且表现出矮生型(图2A )的褐变面积大于乔木状天山樱桃(图2B )㊂A.矮生型天山樱桃;B .乔木状天山樱桃㊂图2 低温胁迫下天山樱桃枝条的褐变情况2.3 低温处理对两种类型天山樱桃枝条生理指标的影响2.3.1 脯氨酸 由图3可知,随着温度的降低,两个类型天山樱桃枝条的脯氨酸含量都呈现先增加后减少再增加的趋势,在-20ħ和35ħ条件下,两种类型天山樱桃枝条脯氨酸含量无显著差异,-25,-30和-40ħ条件下,两种类型天山樱桃抗寒性表现出显著差异,乔木状天山樱桃枝58黑 龙 江 农 业 科 学11期条脯氨酸含量显著高于矮生型天山樱桃,在不同的低温胁迫处理下,乔木状天山樱桃枝条的脯氨酸含量均比矮生型天山樱桃枝条的脯氨酸含量高㊂温度在-40~-35ħ时,乔木状天山樱桃枝条的脯氨酸含量上升幅度最大,增加了214.92μg ㊃g -1;矮生型天山樱桃枝条的脯氨酸含量增加幅度最小,仅增加了56.94μg ㊃g -1㊂在温度下降过程中,两种类型天山樱桃间脯氨酸含量变化差异明显㊂图3 低温胁迫下天山樱桃枝条脯氨酸含量的变化2.3.2 可溶性糖 由图4可知,两种类型天山樱桃枝条的可溶性糖含量随着处理温度的下降而持续上升,两种类型天山樱桃枝条可溶性糖含量始终无显著差异㊂-20ħ时,乔木型天山樱桃枝条的可溶性糖含量较矮生型天山樱桃枝条低㊂但随着处理后温度的持续下降,两种型天山樱桃枝条可溶性糖含量迅速增加,至-40ħ时达高峰程度,且乔木状天山樱桃枝条的可溶性糖含量超过了矮生型天山樱桃㊂图4 低温胁迫下天山樱桃枝条可溶性糖含量的变化2.3.3 丙二醛 由图5可知,-35~-25ħ条件下,矮生型天山樱桃枝条丙二醛含量始终显著高于乔木状天山樱桃㊂在-40和-20ħ条件下,两种类型天山樱桃枝条丙二醛含量无显著差异,温度在-30~-20ħ时,两种类型天山樱桃枝条的丙二醛含量均上升,矮生型天山樱桃枝条的丙二醛含量上升幅度最大;温度在-35~-30ħ时,两种类型天山樱桃枝条的丙二醛含量均下降,乔木状天山樱桃枝条的丙二醛含量下降幅度最大;温度在-40~-35ħ时,矮生型天山樱桃枝条的丙二醛含量下降,乔木状天山樱桃枝条的丙二醛含量迅速上升㊂丙二醛含量最高的是矮生型天山樱桃枝条㊂图5 低温胁迫下天山樱桃枝条丙二醛含量的变化2.3.4 可溶性蛋白质 由图6可知,随着处理温度的降低,两种类型天山樱桃枝条可溶性蛋白质含量整体有上升趋势㊂-25~-20ħ和-35ħ条件下,两种类型天山樱桃枝条可溶性蛋白质含量均无显著差异,在-40和-30ħ条件下,两种类型天山樱桃枝条可溶性蛋白质含量出现显著差异,温度在-30ħ时,矮生型天山樱桃可溶性蛋白质含量达到最大值且显著高于乔木状天山樱桃㊂当温度为-25~-20ħ时,两种类型天山樱桃的可溶性蛋白质含量均迅速升高;当温度为-30~-25ħ时,矮生型天山樱桃的可溶性蛋白质含量缓慢上升,乔木状天山樱桃的可溶性蛋白质含量则缓慢下降;当温度为-35~-30ħ时,矮生型天山樱桃的可溶性蛋白质含量缓慢下降,乔木状天山樱桃的可溶性蛋白质含量又上升;当温度为-40~-35ħ时,两种类型天山樱桃的可溶性蛋白质含量匀下降㊂在整个温度下降过程中,矮生型天山樱桃的可溶性蛋白质含量均比乔木状天山樱桃的可溶性蛋白质含量高㊂图6 低温胁迫下天山樱桃枝条可溶性蛋白质含量的变化2.4 低温胁迫后两种类型天山樱桃枝条恢复生长情况由表2和图7可以看出,随着处理温度的降低,两种类型天山樱桃枝条恢复生长能力减弱,两种类型天山樱桃枝条的萌芽率均随着温度的降低而逐步下降㊂当温度降低至-30ħ时,两种类型天山樱桃枝条的萌芽率都降低在50%以内,矮生6811期 叶丽努尔㊃哈德勒等:不同类型天山樱桃的抗寒性初步研究型天山樱桃枝条萌芽率为20.05%,乔木状天山樱桃枝条萌芽率为20.12%,说明已经有部分芽体遭到破坏,乔木状天山樱桃枝条仍处于展叶期,矮生型天山樱桃枝条只处于绿尖期,当温度下降至-35ħ时,两种类型天山樱桃休眠枝条的萌芽率均只有在20%以下,矮生型天山樱桃枝条萌芽率为9.52%,乔木状天山樱桃枝条萌芽率为17.38%,但乔木状天山樱桃枝条萌芽数量仍高于矮生型天山樱桃枝条,当温度在-40ħ时两种类型天山樱桃枝条均没有萌发㊂随着处理温度的降低,乔木状天山樱桃(图7B )恢复生长的能力相对于矮生型天山樱桃(图7A )的恢复生长能力更强㊂表2 低温胁迫下天山樱桃休眠枝条的萌芽率供试枝条萌发率/%-20ħ-25ħ-30ħ-35ħ-40ħ矮生型天山樱桃55.9050.2920.059.520乔木状天山樱桃59.3851.4820.1217.38A.矮生型天山樱桃;B .乔木状天山樱桃㊂图7 低温胁迫下天山樱桃枝条恢复生长的情况3 讨论3.1 天山樱桃相对电导率和抗寒性关系逆境条件下,植株内的各种渗透调控产物大量堆积,从而赋予植株渗透调控的能力[19-20]㊂当树木枝条遭受低温胁迫时,细胞的质膜透性就会变大,进而产生电解质的外渗㊂本研究发现不同低温胁迫下,两种类型天山樱桃枝条的相对电导率随着温度的降低呈直线上升趋势,温度在-40~-35ħ时,两种类型天山樱桃枝条的相对电导率均超过了50%以上,可以进一步利用L o gi s t i c 方程计算得出两种类型天山樱桃枝条的半致死温度为-36.81和-39.91ħ㊂多数研究表明当相对电导率达到50%以上时多数植物已经遭受到不可逆的伤害,关于这一点魏鑫等[21]在越橘品种㊁马小河等[22]在酿酒葡萄的研究上也得出相同的结论㊂3.2 天山樱桃生理指标和抗寒性关系用作植物细胞内渗透调节的可溶性糖㊁可溶性蛋白㊁丙二醛和脯氨酸等在逆境情况下不断积累,有植物细胞渗透调节的功能,而这种化合物也被认为和耐寒性有关[23]㊂本研究发现两种类型天山樱桃的可溶性糖含量由于温度的不断下降整体呈现不断增加趋势㊂这一方面,在常绿阔叶的灌木富贵草[24]㊁观光木[25]的研究结果与本研究一致㊂天山樱桃本身就是极耐寒的品种,枝条内积聚了丰富的可溶性糖,可溶性糖是植株内主要的渗透保护物质,具有降低原生质冰点缓冲细胞脱水的作用,其含量与抗寒性呈正相关㊂本研究仅对两种不同类型天山樱桃进行了人工模拟低温胁迫下的抗寒性研究,为获得更加准确可靠的数据,仍需要通过连续多年的观测和田间表现进行验证试验㊂生理生化指标的测定及形态结构的观察只是对抗寒性的初步研究,未来还需要在抗寒基因的挖掘㊁功能鉴定等多方面进行深入研究㊂全面评价天山樱桃的生态适应性,为这一野生种质资源的开发利用提供理论依据㊂4 结论通过对人工低温处理条件下不同类型天山樱桃枝条组织褐变和恢复生长情况的观测,拟合L o gi s t i c 方程和脯氨酸㊁可溶性糖㊁可溶性蛋白㊁丙二醛等生理指标对抗寒性进行综合评判,乔木状的天山樱桃半致死温度为-39.91ħ,矮生型天山樱桃休眠枝条的半致死温度为-36.81ħ,且矮生型天山樱桃枝条相对电导率始终显著高于乔木状天山樱桃㊂在-40ħ条件下,矮生型天山樱桃枝条的脯氨酸含量达到271.11μg ㊃g -1,乔木状天山樱桃枝条的脯氨酸含量高达495.11μg ㊃g -1,乔木状天山樱桃枝条脯氨酸含量始终显著高于矮生型天山樱桃㊂矮生型天山樱桃枝条的可溶性糖含量为21.63m g ㊃g -1,乔木状天山樱桃枝条的可溶性糖含量为21.43m g ㊃g -1㊂矮生型天山樱桃枝条的丙二醛含量为50.12n m o L ㊃g -1,乔木状天山樱桃枝条的丙二醛含量为45.65n m o L ㊃g -1,矮生型天山樱桃枝条丙二醛含量始终显著高于乔木状天山樱桃㊂矮生型天山樱桃枝条的可溶性蛋白质含量为16.56m g ㊃g -1,乔木状天山樱桃枝条的可溶性蛋白质含量为10.75m g ㊃g -1,温度在-40ħ时,矮78黑 龙 江 农 业 科 学11期生型天山樱桃可溶性蛋白质含量达到最大值且显著高于乔木状天山樱桃㊂认为两种类型天山樱桃均有强的抗寒能力,半致死温度在-40~-35ħ之间,经综合判断乔木状天山樱桃抗寒力较强于矮生型天山樱桃㊂参考文献:[1] 张力思.甜樱桃的起源㊁分布及栽培现状[J ].北方果树,2000(4):31.[2] 彭妮,周龙,王超,等.天山樱桃种子萌发特性研究[J ].经济林研究,2015,33(3):145-149.[3] 王超,周龙,彭妮,等.天山樱桃花粉特性研究[J ].新疆农业大学学报,2013,36(6):449-453.[4] 李春侨,周龙,陆彪,等.天山樱桃种质资源表型多样性研究[J ].西北农业学报,2018,27(1):91-97.[5] 冯琳骄,褚佳瑶,高林,等.新疆伊犁和塔城地区天山樱桃群落特征与植物区系的比较[J ].经济林研究,2021,39(3):156-164.[6] 李春侨,周龙,陆彪,等.天山樱桃野生居群遗传多样性S S R分析[J ].经济林研究,2017,35(3):44-49.[7] L Y O N S JM.C h i l l i n g i n j u r yi n p l a n t s [J ].A n n u a l R e v i e wo f P l a n tP h y s i o l o g y a n d P l a n t M o l e c u l a rB i o l o g y ,1973,24:445-448.[8] 林艳,郭伟珍,徐振华,等.大叶女贞抗寒性及冬季叶片相对电导率变化研究[J ].天津农业科学,2012,18(5):145-149.[9] 李纪元,高传璧,郑学为,等.两个温带相思属树种的耐寒性评估[J ].林业科学研究,1999,12(1):87-91.[10] 何开跃,李晓储,黄利斌,等.冷冻胁迫对福建柏苗可溶性糖和丙二醛(M D A )含量的影响[J ].江苏林业科技,2007,27(6):6-8.[11] 严寒静,谈锋.栀子叶片生理特性与抗寒性的关系[J ].植物资源与环境学报,2005,14(4):21-24.[12] 高志红,章镇,韩振海.果梅种质枝条抗寒性鉴定[J ].果树学报,2005,22(6):709-711.[13] 姜卫兵,王业遴,马凯.渗透保护物质在无花果抗寒性发育中的作用[J ].园艺学报,1992,19(4):371-372.[14] S U K UMA R A N NP ,W E I S E RCJ .M e t h o d o f d e t e r m i n i n gc o l dh a rd i ne s s b y e l e c t r i c a l c o n d u c t i v i t yi n p o t a t o [J ].H o r t S c i e n c e ,1972(7):467-468.[15] 唐海霞,杨雪梅,冯立娟,等.3个石榴品种越冬抗寒性及生理差异分析[J ].园艺学报,2023,50(7):1563-1573.[16] 苏向辉.天山樱桃生物学特性初步研究及资源评价[D ].乌鲁木齐:新疆农业大学,2012.[17] 牛立新,贺普超.用改进的组织褐变法鉴定葡萄抗寒性的研究[J ].西北农业大学学报,1992,20(1):95-99.[18] 王文举,张亚红,牛锦凤,等.电导法测定鲜食葡萄的抗寒性[J ].果树学报,2007,24(1):34-37.[19] 苏李维,李胜,马绍英,等.葡萄抗寒性综合评价方法的建立[J ].草业学报,2015,24(3):70-79.[20] 玉苏甫㊃阿不力提甫,阿依古丽㊃铁木儿,帕提曼㊃阿布都热合曼,等.利用隶属函数法综合评价梨砧木抗寒性[J ].中国农业大学学报,2014,19(3):121-129.[21] 魏鑫,刘成,王兴东,等.6个高丛越橘品种低温半致死温度的测定[J ].果树学报,2013,30(5):798-802.[22] 马小河,唐晓萍,董志刚,等.6个酿酒葡萄品种抗寒性比较[J ].山西农业大学学报,2013,33(1):1-5.[23] 杨凤翔,金芳,颜霞.不同草莓品种抗寒性综合评价[J ].果树学报,2010,27(3):368-372.[24] 严海燕,董然,于向丽,等.常绿阔叶灌木富贵草越冬过程中抗寒性生理指标的动态变化[J ].北方园艺,2012(24):70-73.[25] 林宁,谢安德,王凌晖,等.低温胁迫对观光木幼苗离体叶片生理特性的影响[J ].湖北农业科学,2012,51(16):3524-3527.P r e l i m i n a r y S t u d y o nC o l dR e s i s t a n c e o fD i f f e r e n t T y p e C e r a s u s t i a n s c h a n i c a P o ja r k Y e l i n u r ㊃H a d l e r ,L I UX u a n ,W A N GY u p i n g ,M A N H a o ,Z H O UL o n g(C o l l e g e o fH o r t i c u l t u r e ,X i n j i a n g A g r i c u l t u r a lU n i v e r s i t y ,U r u m qi 830052,C h i n a )A b s t r a c t :I no r d e rt o p r o m o t et h ef u r t h e rd e v e l o p m e n ta n du t i l i z a t i o no f C e r a s u s t i a n s c h a n i c a P o ja r k ,t h e o n e -y e a r -o l db r a nc h e s o f t w o t y p e s o f C .t i a n s c h a n i c a P o j a r kw e r e u s ed a se x p e r i m e n t a lm a t e r i a l s .T h e p h y s i o l o gi c a l i n d e x e s s u c ha s r e l a t i v e c o n d u c t i v i t y ,s o l u b l e s u g a r ,s o l u b l e p r o t e i n ,p r o l i n e a n dm a l o n d i a l d e h y d e c o n t e n tw e r e m e a s u r e db y a r t i f i c i a l s i m u l a t e d l o wt e m pe r a t u r e s t r e s s t r e a t m e n t (-20,-25,-30,-35,-40ħ).T h e s e m i -l e t h a l t e m p e r a t u r e (L T 50)w a s c a l c u l a t e db y r e l a t i v e c o n d u c t i v i t y c o m b i n e dw i t hL o g i s t i c e q u a t i o n .T h e r e l a t i o n s h i p b e t w e e ne a c h p a r a m e t e ra n dc o l dr e s i s t a n c e w a sc o m p r e h e n s i v e l y e v a l u a t e db y t i s s u eb r o w n i n g m e t h o da n dr e c o v e r yg r o w t h m e t h o d ,a n de v a l u a t et h ec o l dr e s i s t a n c eo fd if f e r e n t t y p e so f C .t i a n s c h a n i c a P o j a r k .T h er e s u l t ss h o w e dt h a tw i t ht h e i n c r e a s eo f l o wt e m p e r a t u r es t r e s s i n t e n s i t y ,t h ec o n d u c t i v i t y a n d s o l u b l e s ug a r c o n t e n t o f b r a n ch e si n c r e a s e d s i g n i f i c a n t l y .W h e n t h e t e m p e r a t u r e r e a c h e d -40ħ,t h e r e l a t i v e c o n d u c t i v i t y o f b o t h e x c e e d e d 50%.T h e s e m i -l e t h a l t e m p e r a t u r e o f d w a r f C .t i a n s c h a n i c a b r a n c h e sw a s-36.81ħ,a n d t h e s e m i -l e t h a l t e m pe r a t u r e of a r b o r C .t i a n s c h a n i c a b r a n c h e sw a s-39.91ħ.I t c a nb e s e e n t h a t t h e c o l d r e s i s t a n c e o r d e r o f t h e t w o t y pe s of C .t i a n s c h a n i c a i s a r b o r C .t i a n s c h a n i c a >d w a r f C .t i a n s c h a n i c a .K e yw o r d s :C e r a s u s t i a n s c h a n i c a P o j a r k ;l o wt e m p e r a t u r e s t r e s s ;c o l d r e s i s t a n c e ;p h y s i o l o g i c i n d e x 88。
冬小麦抗寒性研究进展小麦按生长习性可分为冬小麦和春小麦两大类。
与春小麦相比,冬小麦的营养期较长,从营养期到生殖期生长缓慢。
活动结束后,它可以在春天的长阳光下绽放。
因此,全球冬小麦种植比例远高于春小麦。
冬小麦磨成的面粉味道鲜美,产量高,是其他作物特别是大豆茬的首选作物。
但严寒气候减少了适宜冬小麦种植的地理分布区域,给农业生产造成了巨大损失。
因此,探索冬小麦抗寒性的影响因素及其机制,不仅具有重要的基础理论意义,而且在解决生产中的实际问题方面具有广泛的应用价值。
1 影响冬小麦抗寒性的因素研究1.1 基因型不同冬小麦品种的抗冻性和抗寒性存在较大差异。
冬小麦通常可分为强冬、冬、半冬和弱春四种。
总的来说,各品种的耐寒性依次为:强冬性品种>冬性品种>半冬品种>弱春品种。
目前国外大多以小麦分蘖临界致死温度(LT50)作为小麦冻害生理指标,也是小麦抗冻性的标志。
冬小麦在经历了严冬后的次年能否继续存活,关键在于分蘖节是否受损。
分蘖节是一组压实在一起的节,如植株下部未伸长的节间、节和腋芽,具有再生能力。
冬天地上所有的树叶都冻死了。
只要分蘖节点没有损坏,来年春天就会从分蘖节点发出新的分蘖,继续生长。
1.2 播期、播量、播深播期对冬小麦影响很大。
冬小麦在不同生育阶段和小麦幼苗不同生长条件下的抗寒能力差异很大。
总的趋势是自接合后耐寒能力迅速减弱。
花期对低温最敏感,花期前必须进行低温处理。
早播和加速生长过程将提前结束春化阶段,生长锥伸长并开始拔节,小麦的抗寒性在拔节阶段迅速下降。
这时,遇上低温寒潮,主茎和大蘖容易冻死;冬前积温不足,出苗晚,幼苗弱,冬前绿叶面积小,地表覆盖不足,地温变化剧烈,水分易流失,光合产物少,贮糖少,耐寒性差,易受病害冻害。
小麦适时播种,可以充分利用入冬前的有效生长积温,积累较好的养分,有利于壮苗和抗寒能力的形成,有利于小麦幼苗安全越冬和稳定生长。
春天过后。
电丰等人。
对区域性冬小麦品种京411进行了5个播期,结果与上述结论一致。
‘黄花丁香’抗寒性研究中期报告
本次研究旨在探究黄花丁香植物的抗寒性能力,以期为该植物的种植和生产提供科学依据。
在此中期报告中,我们将介绍研究的进展情况和初步结果。
一、研究背景和意义
黄花丁香是一种常见的栽培观赏植物,其花期较长,花色鲜艳,适宜在冬季栽种。
然而,由于气候变化等原因,冬季的天气越来越寒冷,这给黄花丁香的生长和开花带来了很大的挑战。
因此,研究其抗寒性能力,有助于探索在寒冷气候下提高黄花丁香生存率和生产效率的途径。
二、研究方法
本次研究选择了四个地点进行实验观测,采用自然环境条件下的试验设计,分别是冷暖气泡、大棚、开阔地和林冠下。
在四个地点进行采样后,使用化学分析和基因测序等方法,对黄花丁香抗寒性相关基因和代谢物进行分析。
同时,还进行了灌溉、施肥等操作,以探索这些操作方法对抗寒性的影响。
三、初步结果
通过我们的研究发现,冷暖气泡和大棚这两个相对封闭的环境,其黄花丁香的抗寒能力显著高于开阔地和林冠下两个自然环境。
同时,我们还测得了一些抗寒性相关基因和代谢物,初步证明了黄花丁香在低温环境下具有比较强的适应能力。
另外,我们还发现灌溉和施肥对该植物的抗寒性也有一定的影响,具体影响机制还需要进一步的研究。
四、下一步工作
接下来,我们将继续进行更详细的抗寒性基因和代谢物的分析,并在黄花丁香的生长发育和寒冷适应性方面开展更深入的实验。
同时,我们还计划进一步探究灌溉和施肥等操作对黄花丁香抗寒性的影响,以期为相关业务提供更科学的建议和指导。
仁用杏抗寒性研究进展田介云;魏安智;刘玉林;王胜淇;刘永红【摘要】仁用杏是以杏仁为主要产品的杏属果树的统称,在北方普遍栽植,耐干旱、抗性强且易于管理,对于改善生态环境、防止水土流失和荒漠化均有良好效果,是理想的经济型树种.但在实际生产中,仁用杏常遭受冬季严寒和早春晚霜的危害,产量受到影响,造成仁用杏的产业化发展受到制约.仁用杏的抗寒研究主要集中于新品种选育和生理生化机制方面:‘围选1号’等抗寒新品种已应用于抗逆研究及生产中;通过抗逆相关.物质的分析也使抗寒机制研究趋于深入.综合论述了仁用杏在分子生物方面的研究及其同科植物的抗寒性等,为仁用杏的进一步抗寒研究做参考和理论依据.%Kernel apricot is a general designation of apricot producing almond.It has the advantages of being tolerant to drought,strong resistance and easy to manage,which are generally suitable to be planted in northern China.Kernel apricot has good effects on improving ecological environment,preventing soil erosion and desertification,making it an ideal non-timber tree species.However,in the actual production,kernel apricot often suffers from coldness in winter and frost in early spring,resulting in the reduction of the yield and limited development of industrialization.In this paper,the general situations and progress of cold resistance on kernel apricot study were summarized from three aspects:new variety breeding,physiological and biochemical mechanism and molecular biology.It would provide a reference and theoretical basis for further study on cold resistance of kernel apricot.【期刊名称】《西北林学院学报》【年(卷),期】2018(033)001【总页数】5页(P174-178)【关键词】仁用杏;品种选育;生理研究;抗寒基因【作者】田介云;魏安智;刘玉林;王胜淇;刘永红【作者单位】西北农林科技大学林学院,陕西杨陵712100;西北农林科技大学林学院,陕西杨陵712100;西北农林科技大学林学院,陕西杨陵712100;西北农林科技大学林学院,陕西杨陵712100;西北农林科技大学林学院,陕西杨陵712100【正文语种】中文【中图分类】S662.2仁用杏是以杏仁为主要食用、药用产品的杏属果树的统称,具有很高的开发利用价值和潜力,同时兼备适应性广、抗逆性强且栽培技术较简单等优点,可用于改善生态环境、防止水土流失和荒漠化,从而成为重要的经济林树种[1-2]。
植物抗寒性状的功能基因组学研究随着全球气候变化,冬季的寒冷已经成为了植物生长和发展的严重威胁。
为了适应寒冷环境,植物积累了多种防御和保护机制,如在冬季进入休眠状态、积蓄营养物质和分泌植物激素等。
同时,植物也通过进化和基因调控等手段,不断提高自身的抗寒能力。
近年来,随着基因组学和生物技术的发展,对植物抗寒性状的功能基因组学研究愈加深入,逐渐揭示了植物抗寒机制的分子基础,也为植物育种提供了新的思路和方法。
第一节:植物抗寒性状的功能基因组学研究现状植物抗寒性状是一种综合性状,涉及植物的多个方面,如光合作用、呼吸作用、质膜透性、细胞壁松弛、离子平衡等。
这些性状和生理反应都受多个基因和调控网络的影响。
因此,研究植物抗寒性状需要涉及到大量的基因和生物学过程,需要多个角度和层面去考虑。
当前,基因芯片、DNA测序、转录组分析、蛋白质组分析等高通量技术已经被广泛应用于植物抗寒性状的研究中。
其中,转录组和蛋白质组分析能够揭示植物在抗寒过程中特定基因和蛋白的表达变化,为深入探究植物抗寒机制提供了有力的工具和信息。
第二节:植物抗寒性状的功能基因组学研究进展随着高通量技术的应用,植物抗寒性状的功能基因组学研究已经取得了一系列重要进展。
以拟南芥为例,基于转录组和蛋白质组分析,已经发现了许多与植物抗寒性状相关的基因和蛋白。
例如,在拟南芥中,PBE1基因编码的蛋白质能够促进植物在低温环境下的生长和发育,同时也能改善植物对寒冷的适应能力;而CIPK23基因编码的蛋白质则被证明在植物抗寒过程中发挥了重要作用。
此外,功能基因组学研究也为探究植物抗寒机制的分子基础提供了新的思路。
例如,利用转录组分析,研究人员发现可溶性冷霜霉素的含量可以影响拟南芥植物对寒冷的适应能力。
在这个过程中,RNA结合蛋白CFP1能够与抗冷冻蛋白CSP41b绑定,从而促进可溶性冷霜霉素的合成,为植物在寒冷环境下存活提供了必要物质。
第三节:比较基因组学在植物抗寒性状研究中的应用比较基因组学是一种近年来发展起来的新兴领域,旨在研究不同物种之间的基因和基因组的异同情况。
河南省主栽油茶品种抗寒性研究邓全恩;丁向阳李建安【摘要】近年来河南省油茶发展迅速,但处于油茶生长的北缘地区,低温是限制河南省油茶产业发展的主要因子.为给河南省油茶良种的选育、引种及栽培区划提供理论依据,分别以'长林4号''长林40号''长林27号''XL210''大别山1号''豫油茶1号'等普通油茶品种为试材,对其叶片分别采用0、-5、-10、-15和-20℃的低温胁迫处理,选择相对电导率(REC)、丙二醛、游离脯氨酸(Pro)、可溶性蛋白等抗逆指标,以Logistic方程计算出半致死温度(LT50),并利用隶属函数法综合分析了不同普通油茶品种的抗寒能力.结果表明:随着胁迫温度的降低,电导率、丙二醛含量、游离脯氨酸含量均逐渐升高,可溶性蛋白先升高后降低;其中,电导率、丙二醛含量和油茶抗寒性均呈负相关,游离脯氨酸、可溶性蛋白和油茶抗寒性均呈相关关系;其半致死温度和隶属函数评价结果一致,6个油茶品种中,'大别山1号'的抗寒性最强,'长林4号'的抗寒性最弱,其他品种的抗寒性居中.【期刊名称】《经济林研究》【年(卷),期】2018(036)003【总页数】6页(P12-16,56)【关键词】油茶;抗寒性;生理机理;河南省【作者】邓全恩;丁向阳李建安【作者单位】中南林业科技大学经济林育种与栽培国家林业局重点实验室,湖南长沙 410004;中南林业科技大学经济林培育与保护省部共建教育部重点实验室,湖南长沙 410004;中南林业科技大学经济林培育与利用湖南省2011协同创新中心,湖南长沙 410004;河南省林业科学研究院,河南郑州 450008;中南林业科技大学经济林培育与保护省部共建教育部重点实验室,湖南长沙 410004;中南林业科技大学经济林培育与利用湖南省2011协同创新中心,湖南长沙 410004【正文语种】中文【中图分类】S794.4油茶Camellia oleifera Abel.属山茶科Theaceae山茶属Camellia L.常绿灌木或中乔木,是指山茶科山茶属植物中油脂含量较高且具有栽培经济价值的一类植物的总称[1]。