基因连锁和交换定律
- 格式:docx
- 大小:120.20 KB
- 文档页数:5

连锁互换定律Document serial number【KKGB-LBS98YT-BS8CB-BSUT-BST108】连锁与互换定律1、连锁遗传:原来在亲本中组合在一起的两个性状在F2中有连在一起遗传的倾向,称连锁遗传。
连锁相包括互引相(AB、ab)、互斥相(Ab、aB)。
2、亲本型:与两亲本相同的性状表现型称为亲本型;不同的称为重组型。
3、完全连锁遗传:仅有亲本型,缺少重组型,eg:仅见于雄果蝇、雌家蚕。
4、不完全连锁遗传:在连锁遗传的同时发生性状的交换和重组;绝大多数生物为不完全连锁遗传。
5、利用测交法验证连锁遗传现象:特点:连锁遗传的表现为: 两个亲本型配子数是相等,> 50%; 两个重组型配子数相等,< 50%。
亲组合类型多, 重组合类型出现少6、交换值(Cv):指不完全连锁的两基因间发生交换的频率(百分率,平均次数) 。
重组值(Rf):不完全连锁的双杂合体产生的重组型配子数占总配子数的比率(百分率)。
通常又把交换值称为重组值。
但严格说,交换值不能等同于重组值,因为若两个基因座之间相距较远,其间发生偶数次多重交换时,结果不形成重组型配子,用重组值代表交换值会造成偏低的估计。
7、连锁群:不能进行自由组合的基因群(位于同一染色体上的基因群)。
特点:一种生物连锁群的数目与染色体的对数是一致的。
即有n对染色体就有n个连锁群。
8、染色体作图:把染色体的多种基因相互之间的排列顺序确定下来。
连锁遗传的特征1)摩尔根连锁互换是经典遗传学第三定律,是孟德尔自由组合定律的补充;2)发生在两对或以上基因间,且基因在染色体上线性排列;3)连锁基因发生在同一对同源染色体上;4)减数分裂偶线期,同源染色体联会,非姐妹染色单体间的互换是形成重组型的分子基础;5)两对基因座间距离越大,交换概率越大、连锁性越弱;6)完全交换即为自由组合,完全不交换即为完全连锁情形;染色体作图(基因定位)方法包括两点测交法和三点测交法计算基因间相对距离(1)非等位基因在染色体上排列的直线距离与基因间的互换率大小有关;(2)遗传学上规定,以互换率的1%作为一个遗传单位将基因定位在一条直线上。
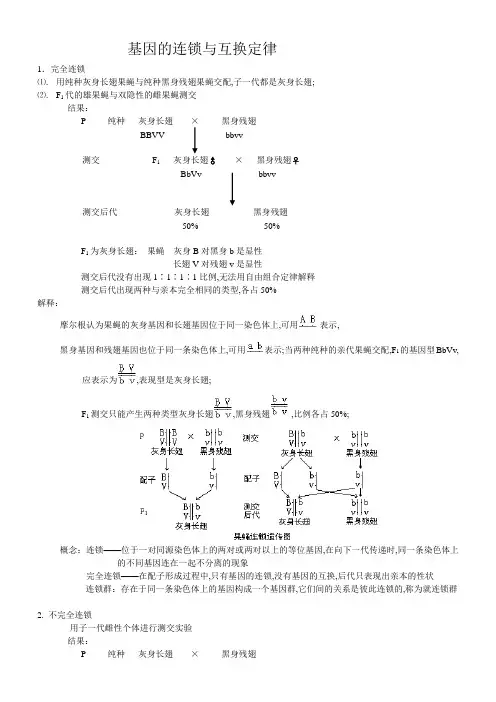
基因的连锁与互换定律1.完全连锁⑴. 用纯种灰身长翅果蝇与纯种黑身残翅果蝇交配,子一代都是灰身长翅;⑵. F1代的雄果蝇与双隐性的雌果蝇测交结果:P 纯种灰身长翅×黑身残翅BBVV bbvv测交F1灰身长翅♂×黑身残翅♀BbVv bbvv测交后代灰身长翅黑身残翅50% 50%F1为灰身长翅:果蝇灰身B对黑身b是显性长翅V对残翅v是显性测交后代没有出现1∶1∶1∶1比例,无法用自由组合定律解释测交后代出现两种与亲本完全相同的类型,各占50%解释:摩尔根认为果蝇的灰身基因和长翅基因位于同一染色体上,可用表示,黑身基因和残翅基因也位于同一条染色体上,可用表示;当两种纯种的亲代果蝇交配,F1的基因型BbVv,应表示为,表现型是灰身长翅;F1测交只能产生两种类型灰身长翅,黑身残翅,比例各占50%;概念:连锁——位于一对同源染色体上的两对或两对以上的等位基因,在向下一代传递时,同一条染色体上的不同基因连在一起不分离的现象完全连锁——在配子形成过程中,只有基因的连锁,没有基因的互换,后代只表现出亲本的性状连锁群:存在于同一条染色体上的基因构成一个基因群,它们间的关系是彼此连锁的,称为就连锁群2. 不完全连锁用子一代雌性个体进行测交实验结果:P 纯种灰身长翅×黑身残翅BBVV bbvv测交F1灰身长翅♀×黑身残翅♂BbVv bbvv测交后代灰身长翅黑身残翅42% 42%灰身残翅黑身长翅8% 8%后代出现四种性状,其中亲本类型占多数,新组合类型占少数;解释:细胞在进行减数分裂形成配子的过程中,减数分裂第一期前期,同源染色体联会,形成四分体;联会复合体中同源染色体间的非姐妹染色单体间会发生染色单体的交叉互换,在交换区段上的基因随染色体发生交换,这种交换产生新的基因组合;交叉互换后形成四种配子,其数量相同;其中有两种配子是亲本类型,两种配子是重组合类型,各占一半;但是在生殖细胞形成过程中,发生交叉互换的性母细胞并不多,所以,减数分裂产生的配子中亲本类型的配子最多,重组合类型的配子占少数;重组类型配子数与所产生的配子总数的比值称为交换率;比如:以上测交实验中,两种重组个体分别占8%,所以,两个基因间的交换率为16%;其性母细胞在进行间数分裂过程中,发生交换的性母细胞占总细胞数的比例是16%×2 = 32%交换率×2 = 发生交换的性母细胞的比例完全连锁0≤交换率≤50%自由组合1 染色体上各个基因间的交换率是不同的,这种差异在反复的试验中总是恒定的2 交换率的大小与基因在染色体上的距离有关,两基因间的距离越小,染色体交叉的机会越小,基因交换率也越小3 基因交换率反映了两基因间的距离:交换率小,距离小;交换率大,距离大4 基因图是根据基因间的交换率绘制的5 生物连锁群的数目与它的染色体的对数是一致的3.基因连锁和交换定律的实质减数分裂形成配子时,位于同一条染色体上的不同基因,常常连在一起进入配子;在减数分裂四分体时期,位于同源染色体上的等位基因有时会随着非姐妹染色单体的交换发生交换,产生基因重组;4.基因连锁和交换定律在实践中的应用1 动植物育种工作,选配优良品种大麦:抗杆锈病抗散黑穗病紧密连锁育种时选择了抗杆锈病植株等同于选择了抗散黑穗病植株不利性状与有利性状连锁:打破基因连锁,促成基因交换,重组成所需基因型2 医学实践中,推测某种遗传病在胎儿中发生的可能性甲髌综合症:人类常染色体显性遗传病,主要症状是指甲发育不良,髌骨缺少或发育不良;致病基因NP与ABO血型的基因I A、I B、i位于同一条染色体上;患病家庭中:NP基因常与I A连锁np基因常与I B、I基因连锁若已知:NP与I B间的重组率为18%,患者的后代患病的概率是:从右边的图分析可以看出,NP与I B间的重组率位18%,产生的配子中、类型配子各占41%,、配子各占9%;与其他几种配子结合后,后代中只要是A型或是AB型血含I A,一般将患甲髌综合症,不患病的可能性是18%;5.基因定位1两基因间在遗传学上的相对图距,交换率的大小反映出连锁基因之间的距离大小,通过交换率的测定,即可以确定基因在染色体上的排列次序和相对距离;遗传学上把交换率的“%”去掉,可作为两基因在遗传学上的相对图距;2基因定位的方法基因定位通常采用“三点测交”法,即用三杂合体跟三隐性个体测交,通过对测定子代表型及比例分析、计算三个连锁基因间的交换值,从而确定各个基因在同一条染色体上的次序和相对距离;值得注意的两个问题:①两边的两个基因的相对距离=两个单交换值之和+两倍的双交换值;②两边的两个基因间的交换值=两个单交换值-两倍的双交换值;另外,不必计算交换值,可直接判断三个基因的次序,其方法为:用亲组合表型的基因次序与双交换表型的基因次序进行比较,发生交换的一对等位基因应位于三对等位基因的中间;学习指导1. 完全连锁遗传后代只出现两种类型,两种类型的比例为1∶1,无重组类型出现,完全连锁现象一般出现在雄果蝇和雌家蚕中;2. 不完全连锁后代出现四种表现型,两种亲本类型占多数,两种重组类型占少数;不完全连锁在生物中普遍存在,不同的基因的连锁的程度不一样;3. 基因的连锁原因是,决定不同形状的两对或两对以上的等位基因位于一对同源染色体上,在遗传时,同一条染色体上的不同基因连在一起,不分离,表现出连锁现象;4. 基因互换的原因是:细胞减数分裂形成配子时,同源染色体上的非姐妹染色单体间发生染色体互换,导致位于其上的等位基因彼此互换;5. 减数分裂时,位于一条染色体上的多数不同基因常常进入同一配子,表现出连锁;部分等位基因会在四分体时期,随着染色体的互换而互换,产生基因重组;6. 互换引起的基因重组,产生生物变异,为生物进化提供了原材料来源;7. 互换率是F1产生的重组配子占总配子数的百分比;8. 交换的可能性与基因间的距离大小有关;交换率=0,表现为完全连锁;交换率=50%,表现为自由组合;0<交换率<50%,表现为不完全连锁;9. 交换的配子百分率是交换的性母细胞百分率的一半,因为任何一个交换的性母细胞所产生的四个配子中总有两个是亲本类型的,两个是重组类型的;10. 不同基因间的交换率不同,这种差异在反复实验中总是恒定的;11. 两个基因间的距离越近,它们之间染色体交叉的机会就越少,基因的交换率就越少,连锁就越强;反之,两个基因距离越远,交换率就越大;12. 存在于一个染色体上的基因构成一个基因群,即是一个连锁群;果蝇有四对染色体,就有四个连锁群;人类的伴性遗传1人类的红绿色盲、血友病为伴X隐性遗传病;红绿色盲、血友病的遗传与果蝇的红眼和白眼遗传方式相同;遗传系谱的主要特点为:①人群中男性患者远多于女性患者,在一些发病率低的系谱中,只有男性患者;②双亲无病,儿子可能发病,女儿则不会发病;③女性患者的儿子必患病;④系谱一般具有交叉遗传的特点;2人类的抗维生素D佝楼病为伴X显性遗传;其系谱特点为:①人群中女性患者多于男性患者,前者的病情可较轻;②患者的双亲中,必有一个该病患者;③男性患者的后代中,女儿都将患病,儿子都正常;④女性患者的后代中,子、女都各有1/2的患病风险;3人类男性外耳道多毛症是伴Y连锁遗传或限雄遗传;其特点是:患者均为男性,并且是父传子,子传孙,女性不会出现相应的遗传病症;人类染色体疾病1.染色体数目异常的疾病1常染色体数目异常——先天愚型或Down综合症软白痴患者的核型为:47,XXXY+21,也称21三体;该病大多是由卵子发生过程中21号染色体不分离,形成了多一条21染色体的异常卵细胞,受精后形成;发病率为:l/600~1/800;2性染色体数目异常的疾病①先天性学儿发育不全症Klinefelter综合病本病患者在青春期出现临床症状,睾丸小且发育不全,不能生育;其核型为47,XXY;该病大多是由卵子发生过程中X染色体不分离,形成多一条X染色体的异常卵子,受精后形成;发病率占男性的1/700~1/800;②性腺发育不全症Turner综合症本病患者只有卵巢基质而无滤泡,无生育能力;其核型为45、XO;该病大多数为精子发生过程中XY不分离,形成了性染色体异常的精子,并和卵子受精后形成;发病率占女性的1/3500;2.染色体结构异常的疾病1猫叫综合症,本病患者哭声如猫叫,智力低,肌张力也低下;患者的核型为46,XXXY,但患者的一条5号染色体的短臂缺失;214/21易位型先天愚型,患者核型为46,XXXY,-14,tt14;21;患儿核型中少一条14号染色体,多一条由14号和21号染色体形成的易位染色体;这种易位可以是新发生的结构畸变,也可以由双亲之一传来;基因表达的调控知识概要一、基因调控的重要意义任何一个细胞在一定时期内,并非全部基因都能表达,而是在一定时期内的一定条件下,只能部分特定基因在表达;所以,基因表达的调控能使生物在利用自然资源和应付生活环境方面很有灵活性,从而使生物可以更好的保存自己,繁衍种族;二、操纵子学说1.原核生物的基因调控——乳糖操纵子的关闭状态和打开状态如下图所示;乳糖操纵子的关闭状态乳糖操纵子的打开状态2.乳糖操纵子的打开状态如图所示;基因的调控系统表明,它使一个代谢系统中的酶系能够同时按所需要的数量准确地合成;乳糖操纵子只有在环境中有乳糖存在时,才开始合成这个酶系,乳糖分解完之后,由于负反馈而停止酶的合成,这就使细菌能更有效地适应环境的变化;3.真核生物基因调控的复杂性;一般地说,包括四个水平的调控;①转录前的调控a.组蛋白转位模型:DNA的一个特定位置上的一种特异性的非组蛋白磷酸化以后,磷酸基带负电荷,于是非组蛋白与带负电荷的DNA相斥,并与带正电荷的组蛋白强烈地结合在一起;组蛋白和非组蛋白复合体从DNA上脱离开来,使这部分DNA裸露出来,不再受组蛋白的抑制而开始转录;b.真核细胞基因调控系统的模型:由于真核生物代谢需很多酶,这些酶的基因可能分散在不同染色体的不同部位上;那么,达到协调机制的原理如面两图所示;真核细胭基因调控系统的模型真核细胞基因的复杂调控系统②转录水平的调控RNA聚合酶I催化rRNA的转录,RNA聚合酶Ⅱ催化mRNA的转录,RNA聚合酶Ⅲ催化tRNA和5sRNA的转录;③转录后的调控对初产生的RNA加工剪接,去掉内含子和非编码区顺序,把几个外显子连接起来,成为成熟的mRNA;④翻译水平的调控,mRNA的戴帽与核糖体小亚基识别并与之结合,是开始翻译的条件;⑤翻译后的调控翻译的最初产物是一个大的蛋白质分子;有时,必须经酶切成更小的分子才能有生物活性;如胰岛素原→胰岛素;细胞质遗传及其在育种上的应用知识概要一、细胞质遗传的特点①遗传方式是非孟德尔式的;②F1通常只表现母方的性状;③杂交的后代一般不出现一定比例的分离;二、高等植物叶绿体的遗传1.紫茉莉的绿白斑的遗传其特点为:不同枝条上的花朵相互受粉时,其后代的叶绿体种类完全决定于种子产生于哪一种枝条上,而与花粉来自哪一种枝条无关;2.玉米的埃型条斑遗传其特点是:①条斑植株作父本,正常植株作母本时,显示孟德尔式遗传;②条斑植株作母本,不论父本基因型怎样,其子代看不到典型的孟德尔式比数,显示出典型的细胞质遗传;三、真菌类线粒体的遗传核基因遗传遵循孟德尔式遗传,但杂交的后代性状跟核基因无直接关系而表现为细胞质遗传如酵母菌的“小菌落”和链孢霉的“缓慢生长”要变型等;四、细胞质遗传在育种上的应用玉米的“二区三系”制种,就是核质互作的一个例子;杂交制种过程如下:Srr ×Nrr Srr ×NRR雄性不育系↓↓保持系↓↓Srr Nrr SRr NRR雄性不育系保持系杂交种恢复系雄性不育系和保持系的繁殖制造杂交种,同时繁殖恢复系植物雄性不育的遗传:植物雄性不育并不完全是由细胞质决定的,它也有核不育型,但是难以找到保持系;细胞质和细胞核互相作用的不育,简称质-核型,表现型比较复杂,当细胞质不育基因S存在时,细胞核内必须有一对隐性基因rr,个体才表现不育;杂交或回交时,只要父本核内没有R基因,杂交后代保持雄性不育;如果细胞质基因是正常可育的N,即使核基因仍然是rr,个体仍然是正常可育的,如果核内存在显性基因R,不论细胞质基因是S 还是N,个体都表现育性正常;雄性不育系:是一种雄性退化主要是花粉退化但雌蕊正常的母水稻,由于花粉无力生活,不能自花授粉结实,只有依靠外来花粉才能受精结实;因此,借助这种母水稻作为遗传工具,通过人工辅助授粉的办法,就能大量生产杂交种子;保持系:是一种正常的水稻品种,它的特殊功能是用它的花粉授给不育系后,所产生后代,仍然是雄性不育的;因此,借助保持系,不育系就能一代一代地繁殖下去;恢复系:是一种正常的水稻品种,它的特殊功能是用它的花粉授给不育系所产生的杂交种雄性恢复正常,能自交结实,如果该杂交种有优势的话,就可用于生产;光合作用专题讲义预备知识一、光合场所:叶绿体的类囊体膜和间质中;二、光合色素:1、叶绿素3:叶绿素a、b3:1,吸收光能,少数叶绿素a能转换光能;2、类胡萝卜素1:括胡萝卜素和叶黄素2:1,吸收光能;3、藻胆素:常结合成澡胆蛋白;三、吸收光谱:红光→蓝紫光770nm→390nm,能量与波长成反比;光合机理光反应暗反应三个阶段1、原初反应2、电子传递和光合磷酸化3、碳同化能量转变光能→电能电能→活跃化学能活跃化学能→稳定化学能贮能物质量子→电子ATP、NADPH 糖类磷酸丙糖转变部位类囊体片层类囊体片层叶绿体基质一、原初反应光能的吸收、传递与转换成电能粗的波浪箭头是光能的吸收,细的波浪箭头是能量的传递,直线箭头是电子传递;空心圆圈代表聚光性叶绿素分子,有黑点圆圈代表类胡萝卜素等辅助色素;P是作用中心色素分子,D是原初电子供体,A是原初电子受体,e是电子二、电子传递和光合磷酸化附:希尔反应Z原始电子供体Q、PQ醌、质体醌Cytf、Cytb细胞色素PC质体蓝素Fd铁氧还蛋白PSII680、PSⅠ700光系统NADP+、NADPH+H+尼克酰胺腺嘌呤二核苷酸磷酸非循环式光合磷酸化三、碳的同化1、卡尔文循环基本途径、又称C 3途径①羧化阶段:1,5-二磷酸核酮糖RuBP 十CO 2→3-磷酸甘油酸②还原阶段:3-磷酸甘油酸A TP →1,3-二磷酸甘油酸NADPH →3-磷酸甘油醛 ③再生阶段:3-磷酸甘油醛→6-磷酸果糖→5-磷酸核酮糖→RuBP3RuBP +3CO 2 −−−−→−ATPNADPH 9/6 PGALd3-磷酸甘油醛+3RuBPPGALd,于叶绿体中形成淀粉,或在细胞质中转变为蔗糖; 2、C 4途径甘蔗、玉米、高梁①固定CO 2的新途径光合效率高:CO 2+磷酸烯酸式丙酮酸PEP →草酰乙酸C4 ②必须进行卡尔文循环在维管束鞘细胞中进行3、景天酸代谢CAM,景天、落地生根植物晚间固定CO 2PEP 附:光呼吸绿色植物在光照条件下吸收氧气,放出CO 2C3>C4不利积累有机物 光合作用的影响因素光照强度、CO 2浓度饱和点与补偿点、温度呼吸作用专题讲义预备知识氧化还原反应:氧化脱氢、失e 、加氧还原加氢、得e 、脱氧 呼吸机理一、有氧呼吸C 6H 12O 6+6O 2−→−酶6CO 2+6H 2O +能量 1、糖酵解细胞质一分子C 6H 12O 6分解为两分子丙酮酸,并且发生氧化脱氢和生成少量ATP;C 6H 12O 6+2NAD ++2ADP +2Pi −→−酶2CH 3COCOOH +2NADH +2H ++2ATP ① C 6H 12O 6 → G –6–P -ATP②G –6–P → 2PGALd -ATP③2PGALd → 2C 3H 4O 34ATP + 2NADH +2H +2、三羧酸循环TCA 、柠檬酸循环线粒体基质C 3H 4O 3彻底分解为CO 2和氢这个氢被传递氢的辅酶携带着,同时生成少量的A TP;→过渡反应丙酮酸的氧化脱羧CH 3COCOOH+辅酶AHSCoA →→乙酰辅酶A CH 3COSCoA-CO 2-2H NAD +→NADH +H +① 乙酰CoA +草酰乙酸4C → 柠檬酸6C+ HSCoA② 柠檬酸 → α–酮戊二酸5C -CO 2-2HNAD +→NADH +H +③ a –酮戊二酸5C → 琥珀酸4C+A TP -CO 2-2H NAD +→NADH +H +④ 琥珀酸4C → 延胡索酸4C-2H FAD →FADH 2线粒体内膜⑤ 延胡索酸4C → 苹果酸4C → 草酰乙酸4C-2H NAD +→NADH +H +附:底物水平磷酸化3、电子传递系统和氧化磷酸化氢氢离子和电子被传递给氧以生成水,并且放出大部分的能量,以生成A TP;①电子传递链是线粒体内膜上的一系列电子传递体,分子氧是最后的电子受体;②NADH+H+→3ATP;FADH2→2ATP③化学渗透学说④磷酸甘油穿梭系统36,肌肉和神经、苹果酸—草酰乙酸穿梭系统38二、无氧呼吸1、酒精发酵C6H12O6+2ADP+2Pi→2C2H5OH+2CO2+2ATP2、乳酸发酵C6H12O6+2ADP+2Pi→2C3H6O3+2ATP植物水分、矿质代谢和成花生理植物细胞的水势与渗透吸水成熟的植物细胞外为纤维素和果胶质组成的细胞壁,中央有一个大的液泡,细胞壁和液泡之间则是细胞的原生质体;从物质透过角度讲,细胞壁是一个完全的透性膜,水分和溶质都可以自由地透过;而原生质膜和液泡膜则是分别透性膜;而且原生质膜和液泡膜之间的中质也并非是任何物质都容易透过的结构;这样我们就有充分的理由将细胞的整个原生质体原生质膜、液泡膜和中质看做是一个分别透性膜;液泡中是具有一定渗透势的溶液,那么植物细胞所处的环境溶液的情况就不外乎三种;即环境溶液的水势高于细胞的水势高水势液,环境溶液的水势低于细胞的水势低水势液,环境溶液的水势与细胞的水势相等等水势液;但不论处在何种情况下,植物细胞与外界溶液之间都能够发生渗透作用,只不过在第三种情况下,由于细胞内外无水势差的存在,外观上没有水分进出细胞的现象发生;所以说一个成熟的植物细胞与外界环境溶液共同构成了一个渗透系统,能够发生渗透作用;植物细胞以渗透吸水为主,吸水的动力来自细胞内外的水势之差,那么,植物细胞的水势又该如何计算呢成熟的植物细胞中央有大的液泡,其内充满着具有一定渗透势的溶液,所以渗透势肯定是细胞水势的组成之一,它是由于液泡中溶质的存在而使细胞水势的降低值;因此又称为溶质势,用ψs 表示;由于纯水的渗透势最大,并规定为0,所以任何溶液的渗透势都比纯水要小,全为负值;当细胞处在高水势溶液中时,细胞吸水,体积扩大,由于细胞原生质体和细胞壁的伸缩性不同,前者大于后者,所以细胞的吸水肯定会使细胞的原生质体对细胞壁产生一种向外的推力,即膨压;反过来细胞壁也会对细胞原生质体、对细胞液产生一种压力,这种压力是促使细胞内的水分向外流的力量,这就等于增加了细胞的水势;这个由于压力的存在而使细胞水势的增加值就称为压力势,用ψp 表示;其方向与渗透势相反,一般情况下为正值;此外,细胞质为亲水胶体,能束缚一定量的水分,这就等于降低了细胞的水势;这种由于细胞的胶体物质衬质的亲水性而引起的水势降低值就称为细胞的衬质势,以ψm表示;所以说,植物细胞的吸水不仅决定于细胞的渗透势ψs,压力势ψp,而且也决定于细胞的衬质势ψm;一个典型的植物细胞的水势应由三部分组成,即ψw=ψs+ψp+ψm;从作用效果看,ψs和ψm是使水分由细胞外向细胞内流的力量;ψp则是使水分由胞内向外渗的力量;就是说ψs和ψm的符号与ψp的符号相反,ψs和ψm为负,而ψp为正;理论上细胞的水势ψw应由ψs、ψp和ψm三部分组成,但ψs、ψp和ψm在细胞水势中所占的比例则是随着细胞的发育时期及细胞所处的状态的改变而变化的;就ψm来讲,干燥种子和未形成液泡的细胞中,ψm是一个很大的负值;而在有液泡的细胞中,由于细胞的衬质部分已被水饱和,ψm等于零或接近于零,其绝对值很小<0.1,相对于绝对值很大的水势来讲,就十分的微不足道了;因此,在计算有液泡细胞水势的时候,ψm通常可以省掉;即有液泡细胞的水势可以用公式ψw=ψs+ψp进行表示和计算;当ψw低于外界溶液时,细胞即可吸水;在一般情况下,细胞的ψp为正值,但处于强烈蒸腾环境中的细胞的ψp为负值,而不为正值;细胞蒸腾失水,细胞体积缩小,最后可失去膨压而达到萎蔫的程度,但此时一般并不能引起质壁分离,原因是水与细胞壁的附着力很强,这样在原生质体收缩时细胞壁被向里拉,甚至发生褶皱变形;同时,细胞壁产生的反作用力使原生质体和细胞液处于张力的状态;张力相当于负的压力;在计算水势时应取负值;因此,在产生张力时,细胞的水势将变得比ψs更负;3.根系吸水及水分沿导管或管胞上升的动力植物体水分的获得主要借助于根系对土壤中水分的吸收;根系的吸水方式有两种,即主动吸水和被动吸水;主动吸水是由于根本身的生理活动而引起的水分吸收,与地上部分的活动无关,吸水动力是根压;被动吸水由蒸腾作用而引起,而与根系的活动无关,吸水的动力是蒸腾拉力;那么,根压和蒸腾拉力是怎样产生的呢关于根压产生的机制现在还不很清楚,一般是用渗透理论来解释;为更好地理解这个理论,必须首先了解植物体的结构;植物体从空间上可分为三个部分,即共质体、质外体和液泡;共质体是指植物体中所有细胞中活的部分,即是指整个植物体的原生质总体;由于各细胞原生质之间有许多胞间连丝相连,所以共质体是一个连续的系统或体系;质外体则是指细胞壁、细胞间隙和木质部导管等原生质体以外的部分;水分和溶质可以在其中自由扩散;和共质体不同,质外体是不连续的,由于内皮层凯氏带的存在,内皮层就将质外体分隔成为两个区域,其一在内皮层外,包括皮层部分的细胞壁、细胞间隙,这部分可以和土壤溶液之间保持水分和溶质的扩散平衡;另一区域在中往内,包括中柱部分的细胞壁、细胞间隙及成熟的导管;内外两部分质外体之间的水分和溶质的交流,都只有通过内皮层细胞原生质体部分来进行;液泡由于有液泡膜和原生质体隔开,所以它既不属于质外体,也不属于共质体,而且它们也不连续成整体;渗透理论认为:土壤中含有丰富的离子,在质外体的外部皮层,离子随土壤溶液进入质外体直至内皮层,溶液中的离子可被活细胞主动吸收,即由质外体进入共质体;在共质体中,这些离子可以通过胞间连丝从一个细胞运至另一个细胞,通过内皮层进入中柱的活细胞;之后细胞中的离子又被动地扩散到导管中,即由共质体进入质外体;其结果,内皮层以外的质外体离子浓度降低,水势增高;而内皮层以内的质外体离子浓度增高,水势降低;这样内外质外体之间就形成了一个水势梯度,于是水经过内皮层的渗透作用而进入中往,进入导管,使导管内产生一种静水压力即根压,水分即沿导管上升;蒸腾拉力是由于蒸腾作用而产生的;由于蒸腾,靠近气孔下腔的叶肉细胞含水量减少;水势降低,向相邻细胞吸取水分,当相邻细胞水势减低时,转向其相邻细胞吸水,如此依次传递直至向导管吸水;这就犹如造成了一种将导管中的水向上拉的力量,这种由于蒸腾作用产生的一系列水势梯度使导管中水分上升的力量就称为蒸腾拉力;主动吸水和被动吸水并存,但二者在根系吸水过程中的比重却很不相同;一般被动吸水占有很大的比重,主动吸水很少;所以蒸腾拉力是根系吸水和水分沿导管或管胞上升的主要动力;这里就产生了一个问题,蒸腾拉力将导管中的水柱向上拉,同时水柱本身的重力又将水柱向下拖,水柱的两端同时受到上拉下拖两种力量的作用,使水柱处于紧张状态,产生张力,水柱就有发生中断的趋势,而导管中水柱的连续性对根系的吸水和水分上升来讲又是必要的;那么,在这种情况下,导管中的水柱如何能保持连续而不发生中断呢众所周知,水分子与水分子之间的内聚力很大,可达-300×105Pa,同时水分子与导管或管胞内纤维素分子之间还有强的附着力,它们远远大于水柱的张力-5~-30×105Pa;所以导管或管胞中的水柱可以保持连续,保证在蒸腾作用进行时木质部中的水分能不断向上运输;这种以水分子的内聚力大于张力来解释水分上升的学说称为内聚力学说,也称为蒸腾——内聚力——张力学说,是19世纪末爱尔兰人迪克松提出的;矿质元素的吸收和运输1.植物细胞对矿质元素的吸收植物细胞吸收矿质元素的方式有三种:被动吸收、主动吸收和胞饮作用;其中被动吸收和主动吸收是植物细胞吸收矿质元素的主要方式;1被动吸收被动吸收:是指由于扩散作用或其他物理过程而进行的吸收,是不需要消耗代谢能量的,故又称非代谢性吸收;①简单扩散:扩散作用是指气体分子、溶剂分子、溶质分子从浓度高的部位向低浓度的部位均匀分布的趋向;当外界溶液的浓度大干细胞内部溶液浓度时,外界溶液中的溶质便扩散进入细胞内,当细胞内外的浓度差大时,细胞大量吸收物质,但随着浓度差变小,吸收也随之减少,直至细胞内外浓度达到平衡为止;所以浓度差是决定细胞靠扩散作用吸收物质的主要因素;②杜南平衡:杜南平衡是一种特殊的积累离子的现象;杜南平衡的结果是膜两侧某离子的浓度不相等,但也达到了平衡;植物细胞的质膜是一种半透膜;细胞内含有许多带电荷的不能扩散到细胞外的大分子化合物如蛋白质,R-,成为不扩散离子,它们可以与阳离子形成盐类如蛋白质的钾盐,KR,设其浓度为Ci,把这样的细胞放在浓度为C;的KCl溶液中,由于细胞内没有Cl-,所以Cl-沿着浓度梯度由外界溶液扩散入细胞内,同时K+也进入细胞内,以。

高中生物难点基因连锁与交换定律的应用高中生物里啊,有个让不少同学头疼的难点,那就是基因连锁与交换定律的应用。
先来说说基因连锁是咋回事儿。
咱们就拿果蝇举个例子吧。
假如有两种性状,眼睛颜色和翅膀形状。
正常情况下,基因是自由组合的。
但在某些时候呢,控制这两种性状的基因就像两个好朋友,手拉手不愿意分开,这就是基因连锁啦。
那基因交换又是咋发生的呢?还是这果蝇,在特殊情况下,这两个好朋友也会闹闹别扭,松开手,互相交换一下位置,这就产生了基因交换。
在实际应用中,这基因连锁与交换定律可有用啦!比如说在农业生产里,咱要培育优良品种。
假如想要一种同时具备抗病虫害和高产的作物,就得研究控制这两种性状的基因是不是连锁的。
要是连锁程度高,那要得到同时具备这两种优点的品种可能就不容易;要是连锁程度低,通过杂交选育,就有更大的机会得到理想的品种。
我记得之前有个学生,他在做一道关于基因连锁与交换的遗传题时,那抓耳挠腮的样子,别提多好笑了。
题目是这样的:有一种植物,红花对白花是显性,高茎对矮茎是显性,控制这两对相对性状的基因是连锁的,交换值是 20%。
现在有红花高茎和白花矮茎的亲本杂交,问F1 代自交产生的 F2 代中,红花矮茎植株所占的比例是多少。
这孩子呀,一开始完全没头绪,就知道在那瞎蒙。
我就引导他,先画出基因连锁图,再算交换值,一步一步来。
最后他总算是算出来了,那高兴的劲儿,就好像中了大奖似的。
再比如说在医学领域,基因连锁与交换定律也能帮助医生诊断一些遗传病。
像血友病,假如知道控制血友病的基因和其他基因的连锁关系,就能更准确地预测家族中患病的风险。
还有啊,在生物进化的研究中,基因连锁与交换定律也能提供一些线索。
不同物种之间,基因连锁的情况可能不同,通过比较,就能了解物种之间的亲缘关系和进化历程。
总之呢,基因连锁与交换定律虽然有点难,但只要咱们多琢磨,多做几道题,多联系实际,就一定能掌握它,让它为咱们的学习和生活服务!相信大家都能在生物的世界里畅游,攻克这个难点,取得好成绩!。




基因连锁互换定律1. 引言基因连锁互换定律是遗传学中重要的定律之一。
它描述了基因在染色体上的连锁分布以及连锁分离的频率。
本文将探讨该定律的基本概念、实验验证以及其在遗传研究中的应用。
2. 基本概念2.1 基因连锁基因连锁指的是在染色体上,位于同一条染色体上的基因之间的相对位置关系保持恒定。
这意味着这些基因在遗传上很有可能一起传递给下一代。
2.2 互换互换是指两条同源染色体之间的交叉互换事件。
在互换事件中,两条染色体上的相同位点上的DNA片段通过交换破裂重连的方式重新组合。
互换可以导致基因连锁的打破,使得原本连锁在一条染色体上的基因出现重新组合。
3. 实验验证基因连锁互换定律最早是通过托马斯·亨特·摩尔根(Thomas Hunt Morgan)的果蝇实验得出的。
通过对果蝇基因型的观察和分析,摩尔根发现某些基因总是共同遗传,这与经典遗传学中的独立性原则相违背。
3.1 果蝇实验摩尔根在实验中选择了具有明显表型差异的果蝇,例如翅膀的颜色。
他通过交配不同表型的果蝇,观察它们后代的表现,并推断基因的连锁关系。
3.2 结果与结论通过大量的实验证明,果蝇的一对基因位点可以遗传的同时,也可以发生连锁互换。
这一发现支持了基因连锁互换定律的存在。
4. 基因连锁互换定律的意义基因连锁互换定律的发现对于遗传学的发展产生了重要影响。
4.1 进化的证据基因连锁互换定律的存在证明了生物进化过程中的基因发生重组,从而导致物种多样性的增加。
4.2 遗传图谱的构建基因连锁互换定律为遗传学家构建遗传图谱提供了重要依据。
通过观察基因连锁互换事件的频率,可以推断基因之间的距离以及相对位置。
4.3 基因定位和基因治疗基因连锁互换定律也为研究基因定位和开展基因治疗提供了基础。
通过对基因连锁的研究,科学家可以确定染色体上具体基因的位置,从而更好地理解基因功能和相关疾病。
5. 总结基因连锁互换定律是遗传学的重要定律之一。
通过果蝇实验和大量实验证据,科学家证明了基因之间的连锁关系以及连锁互换的发生。


高一生物知识点:基因的连锁和交换定律高一生物为我们展示了一个丰富多彩的生物界,是一门十分有意思的学科。
高一生物的学习需要将所有知识点进行总结,方便大家集中记忆。
但是如何进行总结是摆在同学们面前的一个难题,下面小编为大家提供高一生物知识点:基因的连锁和交换定律?,供大家参考,希望对大家学习有帮助。
基因的连锁和交换定律名词:1、基因的连锁:位于同一条染色体上的不同基因在减数分裂过程形成配子时,常常连在一起不相分离,进入配子,这种现象,我们把它叫做基因的连锁。
2、基因的互换:位于同一条染色体上的不同基因,在减数分裂的四分体时期,由于同源染色体上的等位基因随着非姐妹染色体单体的交换而发生互换的现象,我们称之为基因的互换。
3、不完全连锁遗传:像这种雌果蝇的遗传,基因既有连锁,又有互换的现象。
4、完全连锁:雄果蝇的遗传只有连锁,没有互换。
语句:1、完全连锁的实例:用果蝇做杂交实验:纯种的灰身长翅与黑身残翅杂交,?F1代为灰身长翅,所以,灰身长翅为显性,黑身残翅为隐性,对?F1代中的雄性个体测交,测交后代的表现型是?1灰身长翅:1黑身残翅,与?F1代完全相同。
2、不完全连锁杂交实例:选择F1中的雌性BbVv测交:BbVv?X?bbvv→42%BbVv:42%bbvv:8%Bbvv:8%bbVv。
2、比较完全连锁与不完全连锁的异同。
(1)相同点:二组杂交的亲代与F1代情况相同。
(2)不同点:完全连锁的测交后代只有两种基因型,与亲本相同,数量比1:1。
不完全连锁的测交后代有四种基因型,其中亲本基因型(与其亲本相同的基因型)各占42%,重组基因型(与其亲本不同的基因型)各占8%。
3、(1)自由组合是分析分别位于二对同源染色体上的二对等位基因的遗传规律,A(a)与B(b)由于自由组合,产生四种数量相等的配子。
表达式为Aa Bb→1AB:1Ab:1aB:1ab。
(2)完全连锁是分析共同位于一对同源染色体上的二对等位基因的遗传规律,A(a)与B(b)由于完全连锁,所以,产生两种数量相等的配子。

摩尔根基因的连锁和交换定律基因的连锁和交换定律的实质是:在进行减数分裂形成配子时,位于同一条染色体上的不同基因,常常连在一起进入配子;在减数分裂形成四分体时,位于同源染色体上的等位基因有时会随着非姐妹染色单体的交换而发生交换,因而产生了基因的重组。
应当说明的是,基因的连锁和交换定律与基因的自由组合定律并不矛盾,它们是在不同情况下发生的遗传规律:位于非同源染色体上的两对(或多对)基因,是按照自由组合定律向后代传递的,而位于同源染色体上的两对(或多对)基因,则是按照连锁和交换定律向后代传递的。
背景孟德尔遗传的两个基本定律在得到科学界的公认以后,受到了广泛的重视,许多生物学家开始用其他的动物和植物作材料,进行杂交试验。
但是,他们在进行两对相对性状的杂交试验时发现,并不是所有的结果都符合基因的自由组合定律,于是,有人一度对孟德尔提出的遗传定律产生了怀疑。
这时,美国的遗传学家摩尔根和他的同事们用果蝇作试验材料,进行了大量的遗传学的研究工作,不仅证实了基因的分离定律和自由组合定律是正确的,而且揭示出了遗传的第三个基本定律--基因的连锁和交换定律,科学地解释了孟德尔的遗传定律所不能解释的遗传现象。
研究简述摩尔根等人用纯种灰身长翅果蝇与纯种黑身残翅果蝇交配,他们看到子一代(F1)都是灰身长翅的,由此可以推出,果蝇的灰身(B)对黑身(b)是显性;长翅(V)对残翅(v)是显性。
所以,纯种灰身长翅果蝇的基因型与纯种黑身残翅果蝇的基因型应该分别是(BBVV)和(bbvv)。
F1的基因型应该是(BbVv)。
完全连锁遗传试验摩尔根又让F1的雄果蝇(BbVv)与双隐性类型的雌果蝇(bbvv)测交,按照自由组合定律,测交后代中应该出现4种不同的类型,即灰身长翅、灰身残翅、黑身长翅、黑身残翅,并且它们之间的数量比应该为1:1:1:1。
但是,测交的结果与原来预测的完全不同,只出现两种和亲本完全相同的类型:灰身长翅(BbVv)和黑身残翅(bbvv),并且两者的数量各占50%。

高中生物基因连锁和交换定律教案旧人教必修2教学目的:知识目标知道:1.基因的连锁和交换的实验及结果。
2.基因的连锁和交换定律本质。
识记:基因连锁和交换定律在实践上的应用。
能力目标1.通过自由组合定律与连锁和交换定律的对比学习,培养学生分析归纳能力。
2.培养学生解决遗传问题的技能技巧。
情感目标1.通过摩尔根和他的同事们果蝇试验的学习,培养学生的协作精神。
2.运用辩证唯物主义观点分析和认识生物体生命活动的基本规律,逐步树立科学的世界观。
教学重点:1.对完全连锁和不完全连锁现象的解释。
2.基因的连锁和交换定律的实质。
教学难点:1.基因的连锁和交换与基因自由组合的区别。
2.对完全连锁和不完全连锁现象的解释。
3.基因的连锁和交换定律在医学实践中的应用。
教学用具:雄果蝇的连锁遗传图;雌果蝇的连锁交换遗传图;投影仪;多媒体课件。
课时安排:1课时教学过程:导入新课:前面通过学习知道一对相对性状的豌豆杂交后代会发生性状分离。
两对或多对相对性状的豌豆杂交后代会发生自由组合现象。
是否所有的两对或多对相对性状都遵循孟德尔的遗传定律。
1906年,英国生物学家贝特森(W.Bateson,1861~1926)用香豌豆作了如下试验:P 紫花长花粉×红花圆花粉F1 紫花长花粉F2 紫长69%紫圆6%红长6%红圆19%即F2表现型比不符合9:3:3:1的比例,且后代中的亲代性状表现出连在一起传递的倾向,这是孟德尔自由组合定律不能解释的连锁遗传。
教学目标达成:三、基因的连锁和交换定律(一)摩尔根简述学生阅读教材第36页,应知道美国的遗传学家摩尔根和他的同事们用果蝇作试验材料,既证明了孟德尔两大定律的正确性,而且揭示出遗传的第三个基本定律——基因的连锁和交换定律,科学地解释了孟德尔的遗传定律不能解释的遗传现象。
(二)完全连锁遗传学生阅读教材第37~38页。
提问:摩尔根采用什么材料和研究方法进行试验的?学生集体回答:果蝇,杂交法。
必修本第36课时 基因的连锁和交换定律知识精华完全连锁 现象:F 1灰身长翅♂(BbVv)×黑身残翅♂(bbvv) →1灰身长翅(BbVv):1黑身残翅(bbvv)原因: (只有亲本类型,没有重组类型) ♂)−−−→−减数分裂1:1 , F 1雄果蝇产配子时,同一条染色体上的不同基因常连在一起不分离,所以只有亲本类型的配子没有重组类型的配子。
不完全连锁现象:F 1灰身长翅雌(BbVv)×黑身残翅雄(bbvv)↓灰长(BbVv):黑残(bbvv):灰残(Bbvv):黑长(bbVv)42 : 42 : 8 : 8(亲本类型特别多,重组类型特别少)原因:B V B V b v B v b V(♀) : :F 1雌果蝇产配子时,部分初级卵母细胞的同一条染色体上不同基因因同源染色体的非姐妹染色单体之间的交叉互换,即产生亲本类型的配子又产生了重组类型的配子。
理论意义:基因的不完全连锁即互换能引起基因重组,使生物产生变异实践意义:根据育种目标选择杂交亲本时,必须考虑性状间的连锁关系题例领悟例题:基因型为的精原细胞120个,其中若有30个在形成精子过程中发生互换,在正常发育下能产生亲本基因型和重组基因型的精子数目依次是( ) A 、480、480、60、60 B 、210、210、30、30C 、90、90、30、30D 、240、240、60、60解析:120个精原细胞最多产生480个精子,所以A 、D 选项错误。
由于交叉(互换)发生在同源染色体的两条非姊妹染色单体之间,一个基因型为 B v b vB v b vb v 42 42 8B D b dB D精原细胞由于发生了互换,产生的四个精子中(BD ,bd ,Bd ,Bd),只有两个是重组基因型的精子(Bd ,Bd)。
30个精原细胞形成的120个精子中,重组基因型的精子(Bd ,bD)应各有30个。
而其余则是两亲本基因型的配子。
该精原细胞的重组率(或交换值)为:(30+30)/480 ×100%=12.5% 答案:B自我评价一、选择题1、在两对相对性状的遗传实验中,如果F 1的测交后代的表现型有两种且比为1:1,那么这个遗传的类型应属于 ( )A 、基因的分离定律B 、基因的自由组合定律C 、基因的完全连锁D 、基因的连锁和交换2、基因型为 的个体,在形成配子过程中,有20%的初级精母细胞发生了互换。
基因的连锁和交换定律
基因的连锁定律和交换定律是遗传学中重要的定律。
基因的连锁定律是指,当两种不同基因位于同一染色体上时,它们会一起遗传。
这意味着,如果一对基因位于同一染色体上,它们不会在细胞分裂过程中分离,而是一起遗传给下一代。
基因交换定律是指,当两种不同基因位于不同染色体上时,它们会在细胞分裂过程中进行交换。
这意味着,如果一对基因位于不同染色体上,它们会在细胞分裂过程中互相交换,从而使它们能够在下一代中分离遗传。
这两个定律是遗传学中重要的基础,它们有助于解释基因遗传的规律,并为研究遗传疾病和基因工程等领域奠定了基础。
基因的连锁定律是由英国遗传学家Thomas Hunt Morgan在20世纪初发现的。
他通过对果蝇(Drosophila melanogaster)的研究发现,当两种不同基因位于同一染色体上时,它们会一起遗传。
这种现象被称为基因连锁。
基因交换定律也是由Thomas Hunt Morgan发现的。
他发现,当两种不同基因位于不同染色体上时,它们会在细胞分裂过程中进行交换。
这种现象被称为基因
交换。
这两个定律是遗传学中重要的基础,它们有助于解释基因遗传的规律,并为研究遗传疾病和基因工程等领域奠定了基础。
连锁定律和交换定律解释了基因遗传的规律,这对基因工程和分子生物学的研究具有重要意义。
例如,在基因工程中,利用这两个定律可以更好地设计基因治疗策略。
在分子生物学中,这两个定律可以帮助研究者更好地理解基因组学和基因调控机制。
孟德尔连锁与互换定律的内容1.引言孟德尔连锁是指遗传学中的一个重要概念,它描述了基因在染色体上的遗传方式。
而互换定律则是遗传学中的另一个基本规律,它描述了染色体间的互换现象。
本文将详细介绍孟德尔连锁和互换定律的相关内容。
2.孟德尔连锁孟德尔连锁是基因在染色体上的遗传方式,它是指染色体上的两个基因由于位于同一染色体上而表现出的共同遗传现象。
孟德尔连锁与基因的位置有关,位于同一染色体上的基因往往会被同时传递给后代,因此它们的遗传方式呈现出一种“连锁”的规律,相互之间不容易独立分离。
2.1孟德尔连锁的原理孟德尔连锁的原理可以通过遗传交叉实验来解释。
在遗传交叉实验中,通过交叉两个不同的个体,可以观察到基因在染色体上的排列方式。
如果两个基因位于同一染色体上并且没有发生基因重组,那么它们就会同时传递给后代,表现为孟德尔连锁。
2.2孟德尔连锁的应用孟德尔连锁在遗传学研究中有着广泛的应用。
通过研究孟德尔连锁关系,可以帮助科学家确定基因的位置和功能,进而理解遗传性状的传递规律。
此外,孟德尔连锁还可用于遗传疾病的诊断和筛查,有助于提高人类健康水平。
3.互换定律互换定律是指同一染色体上的互换事件。
在互换过程中,两条同源染色体之间的交换发生,导致基因的重新组合。
这种互换现象使得位于同一染色体上的基因可以进行重新组合,破坏了孟德尔连锁的规律。
3.1互换定律的过程互换定律的过程可以通过染色体结构的变化来描述。
在互换过程中,两条同源染色体之间的非姊妹染色单体发生交换,形成新的染色体组合。
这种互换现象使得基因在染色体上重新组合,导致孟德尔连锁被打破。
3.2互换定律的意义互换定律的发现对于遗传学的研究有着重要的意义。
它揭示了基因在染色体上的重排现象,说明了孟德尔连锁并非绝对,在基因重组的情况下,基因之间的连锁关系可以被打破。
互换定律的研究帮助我们更好地理解基因的遗传规律,为遗传学研究的进一步深入提供了基础。
4.结论孟德尔连锁与互换定律是遗传学中的重要内容。