平邑甜茶叶片光合速率及叶绿素荧光参数对氯化镉处理的响应
- 格式:pdf
- 大小:899.25 KB
- 文档页数:8
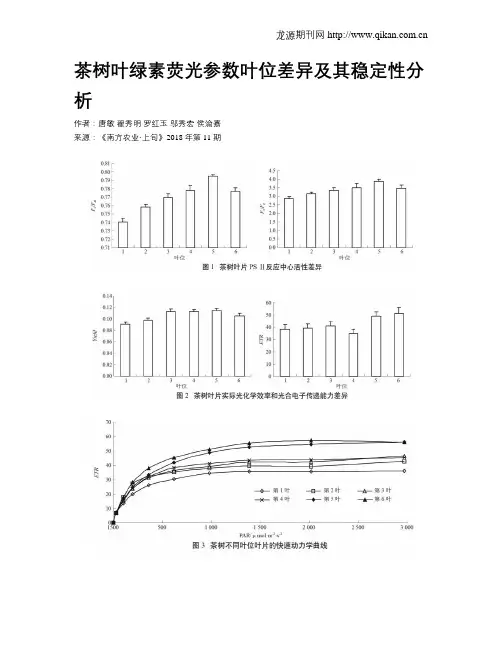
茶树叶绿素荧光参数叶位差异及其稳定性分析作者:唐敏翟秀明罗红玉邬秀宏侯渝嘉来源:《南方农业·上旬》2018年第11期摘; ;要; ;2017年,以福鼎大白茶为试验对象,比较了不同叶位茶树叶片叶绿素荧光参数的差异及稳定性。
结果表明:茶树新梢上部叶片光启动速率快,对强光相对敏感,荧光参数稳定性较差。
随叶位下移,叶片逐渐成熟,到第5叶时,其PSⅡ反应中心最大光能转换效率、潜在活性、实际光化学效率和电子传递速率均处于较高水平,且所测数据变异系数相对较小,表现出较强的稳定性。
建议在进行茶树叶绿素荧光参数测定时,选择芽下第5叶或邻近的功能叶为试验对象以提高数据稳定性,从而更准确地反应茶树光合生理特征。
关键词; ;茶树;叶片;叶绿素;荧光参数;叶位差异;稳定性中圖分类号:Q945.11; ; 文献标志码:A; ; DOI:10.19415/ki.1673-890x.2018.31.003叶绿素荧光分析技术作为植物胁迫生理研究中的一种无损探针,能快速准确地揭示植物对光能的吸收、传递、耗散、分配等特征。
基于荧光数据的分析,能有效反应不同植物或不同品种在抗性、光合潜能、区域适应性等方面的差异[1-4]。
然而,叶绿素荧光参数的差异不仅存在于同一植物的不同品种之间,也存在于同一品种的不同叶位之间[5-7];即使用同一品种的同一叶位叶片进行荧光参数测定,也会因叶片成熟度差异,生理活动的活跃程度差异、对环境的适应能力差异等原因,导致重复测定时数据波动性较大,从而影响试验结果的准确性[8]。
笔者以福鼎大白茶为研究对象,拟通过对不同叶位主要荧光参数的差异及数据稳定性进行分析,以增强试验对象选择时的科学性,从而提高试验数据的稳定性和试验结果的可重复性。
1 材料与方法1.1 试验地点试验地为重庆市农业科学院茶叶研究所种植资源圃,位于重庆市永川区茶山竹海街道。
1.2 试验材料供试品种为三年生福鼎大白茶,按双行双株侧窝种植,大行距150 cm,小行距40 cm,穴距33 cm,常规管理。

西北植物学报,2020,40(9):1518-1526A c t aB o t .B o r e a l .-O c c i d e n t .S i n.d o i :10.7606/j .i s s n .1000-4025.2020.09.1518 h t t p ://x b z w x b .a l l jo u r n a l .n e t 收稿日期:2020-06-07;修改稿收到日期:2020-09-05基金项目:山西省重点研发计划(201703D 221028-1);山西省1331成果转化培育项目(2020C G 027)作者简介:刘媛媛(1994-),女,在读硕士研究生,主要从事园艺作物遗传育种研究㊂E -m a i l :2516806672@q q.c o m *通信作者:段国锋,副教授,主要从事园艺作物遗传育种研究㊂E -m a i l :d a n n e r @s x a u .e d u .c n超重力胁迫下 平邑甜茶 幼苗叶片生理指标及其转录组分析刘媛媛,申成丞,吕红柳,吴 非,段国锋*(山西农业大学园艺学院,山西太谷030801)摘 要:该试验以 平邑甜茶 60d 龄幼苗为材料,设置T 1(5000r /m i n ˑ3h )㊁T 2(5000r /m i n ˑ6h )㊁T 3(10000r /m i n ˑ3h )和T 4(10000r /m i n ˑ6h )超重力处理,以未进行超重力处理为对照(C K ),考察超重力处理下幼苗叶片叶绿素㊁可溶性蛋白㊁丙二醛㊁脯氨酸含量和膜透性等与逆境相关的生理指标的变化,并对其进行转录组分析,初步探究 平邑甜茶 在超重力处理下的转录组响应机制㊂结果表明:(1)超重力处理造成 平邑甜茶 幼苗叶片由舒展状生长状态向不同萎缩生长状态转变,其中T 4处理萎缩生长状态明显㊂(2)超重力处理下, 平邑甜茶 幼苗叶片叶绿素和丙二醛含量在T 3和T 4处理下显著高于CK ,T 1和T 2处理下与C K 无显著差异;叶片脯氨酸含量在各超重力处理下均显著高于C K ;膜透性在T 4处理下显著高于CK ,T 1㊁T 2㊁T 3处理与C K 无显著差异;可溶性蛋白含量在各超重力处理下均不同程度降低,但均与C K 差异不显著㊂(3)样品测序结果显示,大多数转录本的序列长度在1000n t 以上,有37725个,占全部转录本79.15%,且C K ㊁T 2㊁T 4样品以及相对应的新基因中均以FP KM 值小于5.0的转录本数为主,其转录本数依次分别有21412㊁20162㊁22368个和1352㊁1411㊁1406个,占比分别为45.78%㊁43.10%㊁47.82%和81.45%㊁85.00%㊁84.70%㊂(4)G O 功能注释与富集分析显示,随着超重力处理强度的加大,差异表达基因在G O 富集分类中存在明显不同的途径,说明 平邑甜茶 对超重力的响应是复杂的,但主要集中在转录因子活性㊁蛋白质去磷酸化过程方面㊂(5)差异转录因子表达分析显示,随超重力处理强度的增加表达上调幅度比较大的基因家族有b Z I P ㊁W R K Y 和E R F ,其中b Z I P 和W R K Y 转录因子家族中的基因表达量均随超重力处理强度的增加呈上升趋势,但E R F 家族中的MD 01G 1177100㊁MD 08G 1096000㊁MD 13G 1130700基因表达量随超重力处理强度的增加有下降的趋势㊂研究表明,超重力处理对 平邑甜茶 幼苗造成了不同程度的伤害,且叶片总叶绿素㊁丙二醛㊁脯氨酸含量以及膜透性均受到显著影响,故幼苗叶片中叶绿素㊁可溶性蛋白㊁丙二醛及脯氨酸含量以及细胞膜膜透性均可作为评价超重力胁迫的抗性指标;研究推测超重力胁迫对 平邑甜茶 转录因子的影响主要集中在W R K Y ㊁b Z I P 和E R F 转录因子家族㊂该研究结果为候选抗性基因的挖掘和筛选奠定了理论基础㊂关键词:平邑甜茶;超重力;生理指标;转录组中图分类号:Q 945.78;Q 789文献标志码:AP h y s i o l o g i c a l I n d e x e s a n d T r a n s c r i p t o m i c A n a l y s i s o f M a l u s h u p e n e n s i s S e e d l i n g L e a v e s u n d e r H y p e r -g r a v i t y St r e s s L I U Y u a n y u a n ,S H E N C h e n g c h e n g ,L ÜH o n g l i u ,WU F e i ,D U A N G u o f e n g*(C o l l e ge of H o r t i c u l t u r e ,S h a n x i Ag r i c u l t u r a l U n i v e r s i t y ,T a i g u ,Sh a n xi 030801,C h i n a )A b s t r a c t :I n t h i s s t u d y ,w e u s e d 60-d a y -o l d s e e d l i n g s o f M a l u s h u p e n e n s i s a s m a t e r i a l s ,a n d s e t s T 1(5000r /m i n ˑ3h ),T 2(5000r /m i n ˑ6h ),T 3(10000r /m i n ˑ3h )a n d T 4(10000r /m i n ˑ6h )h y p e r -g r a v i t y s t r e s s ,w i t h n o h y p e r -g r a v i t y s t r e s s a s t h e c o n t r o l (C K ).W e i n v e s t i g a t e d t h e c h l o r o p h yl l ,s o l u b l e p r o -t e i n,m a l o n d i a l d e h y d e,p r o l i n e c o n t e n t s a n d m e m b r a n e p e r m e a b i l i t y o f t h e s e e d l i n g l e a v e s u n d e r t h e h y-p e r-g r a v i t y s t r e s s a n d t r a n s c r i p t o m e a n a l y s i s o n t h e m,t o p r e l i m i n a r y e x p l o r e t h e t r a n s c r i p t o m e r e s p o n s e m e c h a n i s m o f M.h u p e n e n s i s u n d e r h y p e r-g r a v i t y s t r e s s.T h e r e s u l t s s h o w e d t h a t:(1)t h e h y p e r-g r a v i t y s t r e s s c a u s e d t h e l e a v e s o f t h e M.h u p e n e n s i s s e e d l i n g s t o c h a n g e f r o m a s t r e t c h e d g r o w t h s t a t e t o a d i f f e r-e n t a t r o p h y g r o w t h s t a t e,a n d t h e T4s h o w e d a s i g n i f i c a n t a t r o p h y g r o w t h s t a t e.(2)U n d e r h y p e r-g r a v i t y s t r e s s,t h e c h l o r o p h y l l a n d m a l o n d i a l d e h y d e c o n t e n t s o f t h e s e e d l i n g l e a v e s o f M.h u p e n e n s i s u n d e r T3a n d T4w e r e s i g n i f i c a n t l y h i g h e r t h a n t h a t o f C K,a n d t h e r e w a s n o s i g n i f i c a n t d i f f e r e n c e b e t w e e n C K,T1a n d T2t r e a t m e n t s.T h e p r o l i n e c o n t e n t o f l e a v e s u n d e r a l l h y p e r-g r a v i t y s t r e s s e s w e r e s i g n i f i c a n t l y h i g h e r t h a n t h a t o f C K.T h e m e m b r a n e p e r m e a b i l i t y u n d e r T4w a s s i g n i f i c a n t l y h i g h e r t h a n t h a t o f C K,a n d t h e r e w a s n o s i g n i f i c a n t d i f f e r e n c e b e t w e e n T1,T2,T3a n d C K.T h e s o l u b l e p r o t e i n c o n t e n t i s r e d u c e d t o d i f f e r e n t d e g r e e s u n d e r e a c h h y p e r-g r a v i t y s t r e s s,b u t i t i s n o t s i g n i f i c a n t l y d i f f e r e n t f r o m C K.(3)T h e s a m p l e s e-q u e n c i n g r e s u l t s s h o w s,t h e s e q u e n c e l e n g t h o f m o s t t r a n s c r i p t s i s m o r e t h a n1000n t,t o t a l o f37725a c-c o u n t i n g f o r79.15%o f a l l t r a n s c r i p t s,a nd C K,T2,T4s a m p le s a n d t h e c o r r e s p o n d i n g n e w g e n e s a r ed o m i n a te d b y t r a n s c r i p t s w i t h F P KM v a l u e<5.0,t h e n u m b e r of t r a n s c r i p t s w e r e21412,20162,22368 a n d1352,1411a n d1406r e s p e c t i v e l y,a c c o u n t i ng f o r45.78%,43.10%,47.82%a n d81.45%,85.00% a n d84.70%,r e s p e c t i v e l y.(4)G O f u n c t i o n a n n o t a t i o n a n d e n r i ch m e n t a n a l y si s s h o w t h a t t h e r e a r e o b v i-o u s l y d i f f e r e n t w a y s o f d i f f e r e n t i a l l y e x p r e s s e d g e n e s i n G O e n r i c h m e n t c l a s s i f i c a t i o n,a s t h e i n t e n s i t y o f h y p e r-g r a v i t y s t r e s s i n c r e a s e s.I t i m p l i e s t h a t t h e r e s p o n s e o f M.h u p e n e n s i s t o h y p e r-g r a v i t y i s c o m p l e x, b u t i t m a i n l y f o c u s e s o n t r a n s c r i p t i o n f a c t o r a c t i v i t y a n d p r o t e i n d e p h o s p h o r y l a t i o n.(5)D i f f e r e n t i a l t r a n-s c r i p t i o n f a c t o r e x p r e s s i o n a n a l y s i s s h o w s t h e b Z I P f a m i l y,t h e WR K Y f a m i l y a n d t h e E R F f a m i l y h a v e a r e l a t i v e l y l a r g e u p-r e g u l a t i o n w i t h t h e i n c r e a s e o f t h e i n t e n s i t y o f t h e h y p e r-g r a v i t y s t r e s s.A m o n g t h e m, t h e g e n e e x p r e s s i o n l e v e l s o f t h e b Z I P a n d WR K Y t r a n s c r i p t i o n f a c t o r f a m i l i e s a l l s h o w a n u p-r e g u l a t i o n t r e n d w i t h t h e i n c r e a s e o f t h e i n t e n s i t y o f t h e h y p e r-g r a v i t y s t r e s s.H o w e v e r,t h e e x p r e s s i o n o f MD01G1177100,MD08G1096000,a n d MD13G1130700i n t h e E R F f a m i l y h a s a d o w n-r e g u l a t i o n t r e n d w i t h t h e i n c r e a s e o f t h e i n t e n s i t y o f h y p e r-g r a v i t y s t r e s s.T h e r e s e a r c h s h o w e d h y p e r-g r a v i t y s t r e s s c a u s e d v a r y i n g d e g r e e s o f d a m a g e t o t h e s e e d l i n g s o f M.h u p e n e n s i s,a n d t h e t o t a l c h l o r o p h y l l,m a l o n d i a l d e h y d e, p r o l i n e c o n t e n t s a n d m e m b r a n e p e r m e a b i l i t y o f t h e l e a v e s w e r e a l l s i g n i f i c a n t l y a f f e c t e d.T h e r e f o r e,t h e c o n t e n t s o f c h l o r o p h y l l,s o l u b l e p r o t e i n,m a l o n d i a l d e h y d e a n d p r o l i n e i n t h e l e a v e s o f s e e d l i n g s,a n d t h e m e m b r a n e p e r m e a b i l i t y c a n b e u s e d a s i n d i c a t o r s o f r e s i s t a n c e t o h y p e r-g r a v i t y s t r e s s.T h e r e s e a r c h s p e c u-l a t e d t h a t t h e e f f e c t s o f h y p e r-g r a v i t y s t r e s s o n t h e t r a n s c r i p t i o n f a c t o r s o f M.h u p e n e n s i s a r e m a i n l y c o n-c e n t r a t e d i n t h e f a m i l y o f WR K Y,b Z I P a n d E R F t r a n s c r i p t i o n f a c t o r s.T h e r e s u l t s o f t h i s s t u d y l a y a t h e o r e t i c a l f o u n d a t i o n f o r m i n i n g a n d s c r e e n i n g c a n d i d a t e r e s i s t a n c e g e n e s.K e y w o r d s:M a l u s h u p e n e n s i s h y p e r-g r a v i t y;p h y s i o l o g i c a l i n d e x;t r a n s c r i p t o m e植物在感受到外界环境的变化后能迅速调整自身来适应新环境[1]㊂影响植物生长发育的环境因子有温度[2-4]㊁光照[5-6]㊁水分[7-8]㊁气体[9]及重力[10-11]等,其中,重力作为较为稳定的因素在植物生长发育过程㊁代谢以及形态建成等方面都有十分重要的作用[12]㊂当前,对植物逆境研究多集中在温度㊁水分㊁元素等因素方面上,而往往忽视对重力的研究,人们早就意识到重力作用可以引导植物器官进行不同方向生长的能力[13],如根的正向重力性和茎的负向重力性[14]㊂航天事业的蓬勃发展,兴起了人们对于微重力条件下植物的感受及适应状态的研究,而超重力的研究更多是在离心机的基础上实现的㊂随着分子生物学的研究发展,R N A测序已成为鉴定候选基因和基因表达模式最常用㊁最有效的方法[15]㊂转录组测序可以在单核苷酸水平上检测植物在某一特定状态下的整体转录活动[16],解释特定的生物学过程和在逆境过程下植物作用的分子机理[17],以及基因调控过程和涉及到的植物生长发育过程的复杂调控机制[18-19]㊂平邑甜茶 (M a l u s h u p e n e n s i s R h e d.)又名湖北海棠,为苹果属(M a l u s)蔷薇科(R o s a c e a e)植物,是优良苹果砧木资源之一[20]㊂重力对植物生物过程和生物生化功能有重要影响[21],已有研究证明超重力会影响种子和根系活力,但利用超重力处理幼苗产生变化的研究还鲜有报道,以及利用超重力处理幼苗来实现植物 交叉适应 并应用于增强植物对逆境抵抗性是否可行还缺少探究㊂本试验以 平邑甜茶 为材料,对其60d龄幼苗进行超重力处理,测91519期刘媛媛,等:超重力胁迫下 平邑甜茶 幼苗叶片生理指标及其转录组分析定其叶绿素㊁丙二醛㊁脯氨酸㊁可溶性蛋白含量和膜透性生理指标,并利用转录组学分析超重力处理下平邑甜茶 的转录组变化,以期对 平邑甜茶 超重力响应机制进行初步探索,为植物的 交叉适应 栽培模式及抗性育种机理提供理论支持㊂1 材料和方法1.1 试验设计将层积露白至出芽的 平邑甜茶 种子播种于50m L 离心管(经改造后底部可透气透水)中,在人工气候箱(温度27ħ,相对湿度60%,光照35000L x ,光暗比16h ʒ8h )中培养至60d 龄幼苗后进行T 1(5000r /m i n ˑ3h )㊁T 2(5000r /m i n ˑ6h )㊁T 3(10000r /m i n ˑ3h )和T 4(10000r /m i n ˑ6h )超重力处理,以未进行超重力处理为对照(C K ),每个处理3个重复,每个重复12管 平邑甜茶 60d 龄幼苗㊂1.2 指标测定将各处理(C K ㊁T 1㊁T 2㊁T 3㊁T 4)幼苗整株迅速脱离离心管清洗干净,摘取茎段所有叶片放入液氮速冻用于后续试验㊂叶绿素(采用鲜样,未放入液氮冷冻)含量测定选用丙酮提取法,膜透性(采用鲜样,未放入液氮冷冻)测定选用电导仪法,丙二醛(M D A )测定选用硫代巴比妥酸(T B A )法,脯氨酸含量测定选用酸性茚三酮法,可溶性蛋白测定选用考马斯亮蓝G -250法,以上测定均采用高俊凤[22]的方法;转录组测定部分由百迈客(北京)完成:采用植物R N A 提取试剂盒(天根生化科技(北京)有限公司)提取总R N A ,检测合格的R N A 样品用于c D N A 文库构建,确保文库质量(有效浓度>2n m o l /L )后由I l l u -m i n a H i S e q 2500平台进行测序㊂使用C u f f l i n k s 软件对样品测序后所得的m a p pe d r e a d s 进行去除接头序列㊁重复冗余序列后拼接,最后使用B L A S T 对D E G s 进行分类和基因功能注释,通过与数据库(G O ㊁N R ㊁C O G ㊁K E G G ㊁K O G 等)信息进行比对获得测序的转录组基因的注释信息㊂1.3 数据处理采用E x c e l 2013和SA S 8.0完成数据方差分析及D u n c a n 多重比较㊂2 结果与分析2.1 超重力处理对平邑甜茶 幼苗叶片抗逆性相关生理指标的影响不同超重力处理对平邑甜茶 幼苗造成不同程度的伤害(图1),超重力处理造成 平邑甜茶 幼苗叶片由舒展状生长状态向不同萎缩生长状态转变,其中T 4处理萎缩生长状态作为明显,同时叶片中叶绿素㊁可溶性蛋白㊁丙二醛及脯氨酸含量以及细胞膜膜透性等均可以作为评价抗性的参考指标㊂图2显示,在不同超重力处理条件下, 平邑甜茶 幼苗叶片中可溶性蛋白呈波动变化的趋势,与对照无显著差异;而叶片总叶绿素㊁丙二醛㊁脯氨酸含量以及膜透性均受到显著影响,且随着超重力处理的增强变化趋势相同,它们均在T 1和T 2处理时与对照(C K )无显著差异,在T 3㊁T 4时多显著高于C K ㊂其中,叶片总叶绿素含量㊁丙二醛含量和膜透性在T 3处理下分别是CK 的2.68㊁5.07和1.24倍,在T 4处理下分别是CK 组的3.36㊁8.10和1.64倍(P <0.05)㊂综合上述结果,后续试验选取C K ㊁T 2和T 4这3个处理进行平邑甜茶 幼苗叶片转录组测序分析㊂2.2 超重力处理对平邑甜茶 幼苗转录组的影响2.2.1 测序结果可靠性分析 首先,对C K ㊁T 2和T 4处理3次重复9个样品RN A 测序的原始数据进行质量控制,即通过除去含有接头的R e a d s 和低质量的R e a d s (N 的比例大于10%㊁同时质量值Q ɤ10的碱基数占50%以上的R e a d s)的方式进行过滤,共得到68.82G b C l e a n D a t a ,其中样品C l e a n D a t a 均达到6.04G b ,各样品Q 30碱基百分比(碱基识别精C K.对照;T 1.5000r /m i n ˑ3h ;T 2.5000r /m i n ˑ6h ;T 3.10000r /m i n ˑ3h ;T 4.10000r /m i n ˑ6h ;下同图1 超重力处理下 平邑甜茶 幼苗形态的变化C K.C o n t r o l ;T 1.5000r /m i n ˑ3h ;T 2.5000r /m i n ˑ6h ;T 3.10000r /m i n ˑ3h ;T 4.10000r /m i n ˑ6h .T h e s a m e a s b e l o w F i g 1 T h e m o r p h o l o g y o f t h e s e e d l i n g s o f M a l u s h u pe n e n s i s u n d e r h y p e r -g r a v i t y t r e a t m e n t 0251西 北 植 物 学 报 40卷度)均不小于92.92%㊂然后,分别对各样品的C l e a n R e a d s 与G D D H 13_v 1.1_G D R 参考基因组进行序列比对,比对率从86.16%到86.97%不等㊂由各样品比对到参考基因组上的结果(表1)可知,匹配读段均不低于86.16%,比对到参考基因组单一匹配读段不低于81.87%,比对到多处位置的多匹配读段平均为2.77%㊂比对率是转录组数据利用率的最直接体现,根据本试验比对效率可知,本次测序质量较高,可用于后续生物信息学分析㊂进一步方差分析结果(表1)显示,G C 含量㊁Q 30百分比和总读数以及样品与参考基因组的比对率在各样品之间均无显著性差异,而单一匹配读数和多匹配读数在C K 组与T 4组之间均存在显著性差异,说明样品间重复性较高,但在C K 组和T 4组之间差异性表达基因存在显著性差异㊂2.2.2 转录本长度分布和表达量情况 通过对样品测序可得,C K ㊁T 2㊁T 4样品中转录本序列长度小于300n t 的转录本有1082个,300~500n t之间有不同小写字母表示处理间在0.05水平存在显著性差异,下同图2 超重力处理下 平邑甜茶 生理指标的变化D i f f e r e n t n o r m a l l e t t e r s i n d i c a t e t h a t t h e r e i s a s i gn i f i c a n t d i f f e r e n c e b e t w e e n t h e t r e a t m e n t s a t 0.05l e v e l .T h e s a m e a s b e l o w F i g .2 T h e p h y s i o l o g i c a l i n d e x e s o f M .h u pe n e n s i s u n d e r h y p e r -g r a v i t y t r e a t m e n t 表1 转录组测序数据质量情况T a b l e 1 T r a n s c r i p t o m e s e q u e n c i n g d a t a q u a l i t y o f R N A -s e q样品S a m p l e G C 含量G C c o n t e n t/%Q 30比例Q 30r a t i o/%过滤后序列C l e a n r e a d s /b p样品与参考基因组的比对率M a p pe d r a t e /%比对序列M a p pe d r e a d s 唯一比对序列U n i q m a p pe d r e a d s 多位置比对序列M u l t i p l e m a p re a d s C K 47.87a 92.99a 44432578a 86.97a 84.20a2.77b T 147.62a 92.92a 42389486a 86.32a83.04a b 3.29a bT 447.66a92.99a44583823a86.16a81.87b4.29aA.依次表示全部转录本长度;B .全部转录本表达量;C .新转录本表达量分布图3 转录本序列长度及表达量分布A.T h e l e n g t h o f a l l t r a n s c r i p t s ;B .T h e e x p r e s s i o n o f a l l t r a n s c r i p t s ;C .T h e d i s t r i b u t i o n o f e x p r e s s i o n o f n e w t r a n s c r i pt s F i g .3 S e q u e n c e l e n g t h a n d e x p r e s s i o n d i s t r i b u t i o n o f t r a n s c r i pt s 12519期 刘媛媛,等:超重力胁迫下 平邑甜茶 幼苗叶片生理指标及其转录组分析A ㊁B ㊁C 依次是T 2与C K ㊁T 4与C K ㊁T 2与T 4GO 注释差异表达基因数目所占百分比及全部基因数目图4 超重力处理下G O 功能注释差异表达基因分布A ,B a n dC a r e p e r c e n t a g e o f t h e n u m b e r o f d i f f e r e n t i a l l y e x p r e s s e d g e n e s a n d t h e n u m b e r o f a l l ge n e s of C K v s T 2,C K v s T 4,T 2v s T 4,re s p e c t i v e l y F i g .4 G Of u n c t i o n a n n o t a t i o n o f d i f f e r e n t i a l l y e x p r e s s e dg e n e s d i s t r i b u t i o n u n d e rh y p e r -g r a vi t y st r e ss A ㊁B ㊁C 依次是T 2与C K ㊁T 4与C K ㊁T 2与T 4的差异表达基因G O 富集图5 超重力处理差异表达基因G O 富集分析A ,B a n dC a r e G O e n r i c h m e n t o f d i f f e r e n t i a l e x p r e s s e d g e n e s o f C K v s T 1,C K v s T 4,T 1v s T 4,re s p e c t i v e l y F i g .5 T h e e n r i c h e d G O of d i f f e r e n t i a l e x p r e s s i o ng e n e s u n d e rh y p e r -g r a vi t y st r e ss 数字1㊁2㊁3代表各样本重复对应图6 不同超重力处理下差异转录因子表达热图N u m b e r s 1,2a n d 3r e p r e s e n t r e p e a t e d c o r r e s po n d e n c e o f e a c h s a m pl e F i g .6 D i f f e r e n t i a l t r a n s c r i p t i o n f a c t o r e x pr e s s i o n h e a t m a p u n d e r h y p e r -g r a v i t y st r e s s 2605个,在500~1000n t 之间的转录本有6250个,而大多数转录本的序列长度在1000n t 以上,有37725个(图3,A ),占全部转录本79.15%㊂对表达量分析可知,C K ㊁T 2㊁T 4样品中FP KM 大于50的转录本分别有1787㊁2371㊁2337个,分别占转录本3.82%㊁5.07%㊁5.00%,F P KM 小于5.0的转录本分别有21412㊁20162和22368个(图3,B ),分别占转录本45.78%㊁43.10%㊁47.82%;相对应地,对新基因中表达量分析可知,C K ㊁T 2㊁T 4样品中F P KM 大于50的转录本分别为16㊁21㊁22个,F P -KM 小于5.0的转录本数分别为1352㊁1411和1406个(图3,C ),占新转录本的比例分别为0.96%㊁1.27%㊁1.33%和81.45%㊁85.00%㊁84.70%㊂2.2.3 差异表达基因G O 功能分类注释 对本次试验的差异表达基因情况进行G O 功能注释结果表明,与C K 相比较,T 2超重力处理造成差异表达基2251西 北 植 物 学 报 40卷因共有5478个(上调2201个,下调3277个),有3959个差异表达基因被注释到G O分类中,生物过程有16个小类别,细胞组分中有14个小类别,分子功能中有11个小类别(图4,A);与C K比较,T4超重力处理造成差异表达基因共有10082个(上调基因数4766个,下调基因数5316个),共有7226个差异表达基因被G O注释,生物过程里有16个小类别,细胞组分有14个小类别,分子功能里有13个小类别(图4,B);T2与T4超重力处理比较,差异表达基因共有3677个(上调1819个基因,下调1858个基因),其中有2656个差异基因被G O注释,同样在生物过程中有16个小类别,细胞组分中有14个和分子功能中有13个小类别(图4,C)㊂2.2.4差异表达基因G O富集分析综合对超重力处理后差异表达基因G O功能注释情况,再对差异表达基因进行G O富集分析㊂对 平邑甜茶 超重力处理后的差异表达基因进行G O富集分析(图5)表明,C K与T2差异表达基因被富集在转录因子活性上,数量达60且显著性更高;C K和T4比较,差异表达基因被富集在核糖体的结构成分功能上,个数仅为16;T2和T4比较,20个差异表达基因被富集在蛋白质丝氨酸/苏氨酸磷酸化功能上㊂随着超重力处理强度的加大,差异表达基因在G O富集分类中存在明显不同的途径,说明 平邑甜茶 对超重力的响应是复杂的,但主要集中在转录因子活性㊁蛋白质去磷酸化过程方面㊂2.2.5差异转录因子表达分析转录因子是位于基因上游特定序列专一性结合位点㊁保证目的基因在特定的时空表达的蛋白质分子,因此对转录因子差异表达基因进一步分析㊂为进一步解释超重力处理 平邑甜茶 幼苗对转录组的影响,对G O富集后的转录因子(F P KM>20)进行B l a s t比对后绘制热图,结果(图6)表明,基因表达上调幅度比较大的基因家族有b Z I P家族㊁W R K Y家族和E R F家族㊂随着超重力处理强度的增加,在b Z I P转录因子家族中的基因表达量呈上升趋势,其中M D06G1002100基因表达量最高且变化最为明显;在E R F家族中的基因表达量变化趋势较为复杂,M D05G1311400基因表达量同样随着超重力处理强度的增加显著提高,而M D01G1177100㊁M D08G1096000㊁M D13G1130700基因表达量随超重力处理强度的增加不但没有提高,反而有下降的趋势;在WR K Y家族里,所有基因的表达量随着超重力处理强度的增加而增加,而且是所有转录因子中表达量最高的基因家族㊂据此推测超重力处理对 平邑甜茶 的影响主要集中在b Z I P㊁WR K Y和E R F转录因子家族㊂3讨论叶绿素是植物在进行光合作用的重要物质,负责吸收并传递太阳光能[23],其含量的多少会直接影响光合作用的强弱[24]㊂本试验中 平邑甜茶 幼苗叶片总叶绿素含量在T4处理下最高,说明超重力处理有利于叶绿素含量的升高,超重力处理造成叶绿素含量的升高可能是由于超重力处理导致细胞壁发生变化,进而影响C O2在叶片内的扩散,进一步影响植物光合作用[25],同时K a o r i等[26]对苔藓进行10g超重力处理发现,超重力处理可以增加叶绿体的直径和厚度,同时提高了苔藓的光合速率㊂本试验对 平邑甜茶 超重力处理时间虽短,但处理强度远远大于10g,猜测对 平邑甜茶 叶片细胞壁和叶绿体造成的影响也远强于苔藓等植物,造成叶绿素含量升高较明显㊂可溶性蛋白和脯氨酸均是渗透调节物质,植物可以通过渗透调节方式提高原生质胶体的稳定性,对细胞和生物膜等起到保护作用[2],本试验中 平邑甜茶 幼苗叶片在T1处理下脯氨酸含量最高,表明超重力处理有利于脯氨酸含量的升高,但可溶性蛋白和脯氨酸含量在各超重力处理间无显著性差异,其原因可能是超重力处理抑制了有机溶质和渗透调节物质的转运,超重力处理部分破坏细胞膜H+梯度的转运进而破环渗透调节物质的转运[27],类似的情况也出现在超重力处理对红豆上胚轴渗透调节物质的影响上[28]㊂植物在逆境条件下首先伤害的是细胞膜系统,膜结构遭到破坏后,膜透性增大,细胞内容物外渗增多,同时M D A含量迅速增加,严重影响植物正常的生长过程㊂本试验中 平邑甜茶 幼苗叶片膜破坏强度在T4处理下最大,膜透性和M D A含量最高,显著高于相应对照㊂这些结果与转录组分析中G O功能注释分类差异表达基因主要集中在代谢过程和膜组分上是一致的㊂转录组测序作为后基因组时代的重要研究方法为基因表达和调控提供了新方法[29],转录组测序目前已经应用于多种植物[30-31]和多种逆境[32-33]条件,挖掘出许多与信号转导途径㊁离子运输过程以及代谢过程相关基因㊂但目前对超重力处理植物的研究多集中在种子和形态学观察方面,超重力直接处理幼苗试验较少[34-37],本试验通过超重力处理 平邑甜茶 幼苗转录组分析发现,超重力处理对 平邑甜茶 的主要影响在b Z I P㊁WR K Y和E R F转录因子32519期刘媛媛,等:超重力胁迫下 平邑甜茶 幼苗叶片生理指标及其转录组分析家族的表达上㊂与M a r t z i v a n o v等[38]对超重力处理影响拟南芥转录组研究结果相同的是差异表达基因显著富集在蛋白质磷酸化和去磷酸化过程,W a t-s o n等[39]认为3-磷酸甘油脱氢酶和磷酸丝氨酸转氨酶等丝氨酸生物合成酶或精氨酸脱羧酶在叶同源器官生长和新陈代谢中起重要作用;不同的是超重力处理拟南芥大部分差异表达基因集中在编码代谢关键酶,而超重力处理 平邑甜茶 更多地集中在转录因子,造成这种不同的原因可能与超重力处理强度不同和拟南芥㊁ 平邑甜茶 两种植物本身对超重力的感知敏感性和忍耐性不同有关㊂同时,本研究还与P a o l a等[40]对松果种子在萌发过程中涉及到的关键酶基因表达有差异,造成这种差异的原因可能与植物当时所处的生长发育阶段有关,松果在萌发期乙醛酸循环起着重要的作用,更多的酶基因集中在乙醛酸循环系统内,而 平邑甜茶 处于生长发育阶段,更多的差异表达基因集中在转录因子上㊂这些转录因子家族与植物生物胁迫和非生物胁迫都有很大关系,会诱导植物免疫系统产生脯氨酸㊁可溶性蛋白及多种抗氧化酶来保护植物自身㊂首先,WR K Y转录因子家族是植物在受到外界胁迫发生自身免疫反应时抗病基因表达量最大的转录因子家族之一[41]㊂栾倩倩[42]通过克隆黄瓜C s WR K Y50基因并对其进行黄瓜遗传转化发现,过表达转基因C s WR K Y植株在增强黄瓜对霜霉病抗性的同时其叶绿素含量明显高于野生型;在病菌侵染初期过表达转基因黄瓜植物中丙二醛含量增长较迅速,后期含量较为平稳,而野生型在整个侵染周丙二醛含量变化不大;同样在侵染霜霉病12h后,过表达转基因黄瓜植株中可溶性蛋白含量高于野生型,但随着侵染时间的延长,转基因株系和野生型中可溶性蛋白质含量均有明显的上升㊂杨柳慧[43]通过克隆细叶百合L p WR K Y20基因并对其进行烟草遗传转化得出过表达转基因株系可以增强植株的抗旱性,虽在干旱胁迫下所有株系丙二醛含量均有升高,但转基因株系含量最低㊂转基因株系之所以能够提高植株抗性是因为WR K Y转录因子可以调控相关防御基因,如WR K Y转录因子可以通过S A㊁J A和E T内源分子信号作用诱导一些抗性蛋白(P R蛋白)[44]的表达,也可以激活下游相关的结构基因(S A G T㊁P A L㊁I C S)的表达㊂其次,植物b Z I P转录因子是在真核生物中广泛分布并且相对保守的一类蛋白,其碱性区域高度保守,约包含20个氨基酸残基㊂根据b Z I P结构的不同,可将其划分为10个亚族,不同亚族的转录因子行使不同的功能,主要包括调控植物生长发育过程㊁进行光信号转导㊁预防病害㊁胁迫应答和A B A 敏感性等各种信号的反应㊂王雅琼[45]对苹果转录因子M d b Z I P26基因进行抗逆的功能性研究发现,过表达转M d b Z I P26基因拟南芥在提高对干旱造成的渗透胁迫的同时其电导率和丙二醛含量明显低于野生型,而叶绿素含量比野生型高㊂李雪垠[46]通过对小麦T a b Z I P174基因进行功能研究发现在正常供水条件下转基因和野生型株系在生长周期内没有观察到形态学差异,而进行干旱胁迫处理时,二者在早期阶段生长均正常,而到后期时野生型株系出现明显枯萎现象,转基因株系大多数叶片依旧保持绿色,叶绿素含量和脯氨酸含量表现为转基因株系高于野生型拟南芥㊂T a b Z I P174基因可能通过A B A依赖途径调控植物干旱胁迫下应答反应, A B A激活S n R K2蛋白激酶,进一步磷酸化A R E B 蛋白激活A R E B来调控下游基因的表达[47]㊂再次,E R F(E t h y l e n e-r e s p o n s i v e f a c t o r)转录因子是A P2/E R F大家族中的一个大的亚家族,仅含1个A P2/E R F结构域,每个成员都含有1个由大约60个氨基酸组成的非常保守的D N A结合域㊂每种植物有100种以上E R F转录因子,其功能各不相同,分别具有调节植物生长发育㊁抗生物胁迫和非生物胁迫的作用等㊂张春霄[48]对黄花苜蓿中M f E R F049基因进行抗逆方面功能验证得出,在盐㊁干旱㊁低温和苜蓿中华根瘤菌的胁迫下,过表达转基因植株抗性明显提高且根中脯氨酸含量高于对照植株,丙二醛含量则明显低于对照植株㊂当黄花苜蓿受到外界非生物胁迫时,E R F转录因子会激活下游C A S15A(冷诱导基因家族一员)和L e a14(胚胎晚期发育蛋白基因家族一员)的表达,转基因植株通过负调控C A S15A和正调控L e a14的作用抵抗植物逆境胁迫㊂综上所述,超重力处理对 平邑甜茶 幼苗的影响复杂,超重力作为一种外界环境胁迫,在对植物造成某种伤害的同时也在一定程度上激发植株本身抵抗逆境的机制㊂本试验通过对 平邑甜茶 幼苗叶片抗逆性相关生理指标检测和转录组学分析发现,超重力处理 平邑甜茶 幼苗叶片的膜透性和丙二醛㊁叶绿素含量明显上升,脯氨酸和可溶性蛋白等渗透调节物质含量也有不同程度增加,同时差异表达基因主要集中在WR K Y㊁b Z I P和E R F转录因子㊂前人通过对WR K Y㊁b Z I P和E R F转录因子进行克隆4251西北植物学报40卷和转基因研究发现,过表达相关基因可以增强植株对干旱和霜霉病的抗性等㊂因此,超重力处理对 平邑甜茶 造成的影响可以为植物交叉适应提供一定的理论支持,为后续抗性基因的挖掘㊁筛选等提供一定的参考㊂参考文献:[1]武迪,黄林周,高谨,等.植物重力反应的分子调控机制[J].遗传,2016,38(7):589-602.WU D,HU A N G L Z,G A O J,e t a l.T h e m o l e c u l a r m e c h a-n i s m o f p l a n t g r a v i t r o p i s m[J].H e r e d i t a s(B e i j i n g),2016,38(7):589-602.[2]张铭顺,吕福堂,褚鹏飞,等.外源C a2+提高百合耐热性的生理机制初探[J].河南农业科学,2019,48(2):120-125.Z H A N G M S,LÜF T,C HU P F,e t a l.P r i m a r y s t u d i e s o n p h y s i o l o g i c a l m e c h a n i s m o f e x o g e n o u s c a l c i u m i m p r o v i n g t h e r-m o t o l e r a n c e i n l i l y[J].J o u r n a l o f H e n a n A g r i c u l t u r a l S c i-e n c e s,2019,48(2):120-125.[3]杨锁宁,张黎,等.温度和土壤酸度对八仙花 B a i l m e r 生长发育的影响[J].中国农业科技导报,2020,22(5):24-34.Y A N G S N,Z HA N G L,e t a l.I n f l u e n c e s o f t e m p e r a t u r e a n d s o i l s a c i d i t y o n t h e g r o w t h a n d d e v e l o p m e n t o f H y d r a n g e a m a c r o p h y l l a B a i l m e r [J].J o u r n a l o f A g r i c u l t u r a l S c i e n c ea n d T e c h n o l o g y,2020,22(5):24-34.[4]吴玲利,李建安,王楠,等.低温胁迫对两个油茶品种开花结实及生理特性的影响[J].植物生理学报,2020,56(4):681-692.WU L L,L I J A,WA N G N,e t a l.E f f e c t s o f l o w t e m p e r a-t u r e s t r e s s o n f l o w e r i n g,f r u i t i n g a n d p h y s i o l o g i c a l c h a r a c t e r i s-t i c s o f t w o C a m e l l i a o l e i f e r a v a r i e t i e s[J].P l a n t P h y s i o l o g y J o u r n a l,2020,56(4):681-692.[5]孙朝华,李开,雷喜红,等.不同光周期对水培生菜生长和品质的影响[J].蔬菜,2020,(5):55-59.S U N C H,L I K,L E I X H,e t a l.E f f e c t s o f d i f f e r e n t p h o t o-p e r i o d o n g r o w t h a n d q u a l i t y o f h y d r o p o n i c l e t t u c e[J].V e g e-t a b l e s,2020,(5):55-59.[6]施福军,粟春青,韦艺,等.光氮互作对闽楠幼苗叶片光合生理特性的影响[J].西北植物学报,2020,40(4):667-675.S H I F J,S U C Q,W E I Y,e t a l.E f f e c t s o f l i g h t a n d n i t r o g e ni n t e r a c t i o n o n p h o t o s y n t h e t i c p h y s i o l o g i c a l c h a r a c t e r i s t i c s i n l e a v e s o f P h o e b e b o u r n e i s e e d l i n g s[J].A c t a B o t a n i c a B o r e a l i-O c c i d e n t a l i a S i n i c a,2020,40(4):667-675.[7]李华,张夏,张扬,等.H2S调节小麦干旱胁迫依赖于钙信号途径[J].河南农业大学学报,2019,53(1):10-14.L I H,Z HA N G X,Z HA N G Y,e t a l.R e g u l a t i o n o f d r o u g h t s t r e s s b y h y d r o g e n s u l f i d e d e p e n d i n g o n t h e c a l c i u m s i g n a l i n g p a t h w a y i n w h e a t[J].J o u r n a l o f H e n a n A g r i c u l t u r a l U n i-v e r s i t y,2019,53(1):10-14.[8]陈丽飞,孟缘,陈翠红,等.3种耧斗菜属植物幼苗对不同水分处理的形态生理响应及其抗性[J].西北农林科技大学学报(自然科学版),2020,48(11):1-10.C H E N L F,M E N G Y,C H E N C H,e t a l.M o r p h o l o g i c a l a n dp h y s i o l o g i c a l r e s p o n s e s a n d r e s i s t a n c e o f s e e d l i n g s o f3A q u i l e-g i a L.s p e c i e s t o d i f f e r e n t w a t e r t r e a t m e n t s[J].J o u r n a l o fN o r t h w e s t A&F U n i v e r s i t y(N a t u r a l S c i e n c e E d i t i o n),2020, 48(11):1-10.[9]L I Y M,L I S H,H E X R,e t a l.C O2e n r i c h m e n t e n h a n c e dd r o u g h t re s i s t a n c e b y r e g u l a t i n g g r o w t h,h y d r a u l i c c o n d u c t i v i-t y a n d p h y t o h o r m o n e c o n t e n t s i n t h e r o o t o f c u c u m b e r s e e d-l i n g s[J].P l a n t P h y s i o l o g y a n d B i o c h e m i s t r y,2020,152: 62-71.[10]袁俊霞,陈瑜,李飞,等.模拟失重下铁源对辣椒叶片结构及叶绿素荧光参数的影响[J].航天医学与医学工程, 2018,31(4):425-426.Y U A N J X,C H E N Y,L I F,e t a l.E f f e c t s o f s i m u l a t e d m i-c r o g r a v i t y o n l e a f a n a t o m i c a l s t r u c t u r e c a nd c h l o r o p h y l l p a-r a m e t e r s o f C a p s i c u m a n n u u m g r o w i n g u n d e r d i f f e r e n t i r o ns o u r c e s[J].S p a c e M e d i c i n e&M e d i c a l E n g i n e e r i n g,2018, 31(4):425-426.[11] D U A N G F,G U O X Y,WA N G Z Z,e t a l.E f f e c t s o f h y-p e r g r a v i t y o n p h y s i o l o g i c a l a n d b i o c h e m i c a l c h a r a c t e r s t i c s o fc h i n e s e c a b b a g e[J].A g r i c u l t u r a l S c i e n c e&T e c h n o l o g y,2014,15(10):1631-1633.[12] V O L KMA N N D,B A L UŠK A F.G r a v i t y:o n e o f t h e d r i v i n gf o r c e s f o r e v o l u t i o n[J].P r o t o p l a s m a,2006,229(2/4):143-148.[13] K N I G H T T A.O n t h e d i r e c t i o n o f t h e r a d i c l e a n d g e r m e nd u r i n g t he v e g e t a t i o n of s e e d s[J].P h i l o s o p h i c a l T r a n s a c-t i o n s o f t h e R o y a l S o c i e t y,1832,1:99-108.d o i.o r g/10.1098/r s p l.1800.0119[14]金静.植物对重力信号的感受㊁传递和反应机理的研究[D].杭州:浙江大学,2004.[15] B HA R G A V A A,C L A B A U G H I,T O J P,e t a l.I d e n t i f i c a-t i o n o f c y t o k i n i n-r e s p o n s i v e g e n e s u s i n g m i c r o a r r a y m e t a-a-n a l y s i s a n d R N A-s e q i n A r a b i d o p s i s[J].P l a n t P h y s i o l o g y, 2013,162(1):272-294.[16]赵振宇,王仕玉,郭凤根,等.转录组测序及其在药用植物上的应用[J].基因组学与应用生物学,2017,36(2):820-825.Z H A O Z Y,WA N G S Y,G U O F G,e t a l.R N A-s e q u e n c i n ga n d i t s a p p l i c a t i o n i n m e d i c i n a l p l a n t s[J].G e n o m i c s a n d A p-p l i e d B i o l o g y,2017,36(2):820-825.[17]刘红亮,郑丽明,刘青青,等.非模式生物转录组研究[J].遗传,2013,35(8):955-970.L I U H L,Z H E N G L M,L I U Q Q,e t a l.S t u d i e s o n t h e t r a n s c r i p t o m e s o f n o n-m o d e l o r g a n i s m s[J].H e r e d i t a s(B e i-j i n g),2013,35(8):955-970.[18] L I U L,L I Q Z,J I N W,e t a l.R e v e a l i n g g e n e f u n c t i o n a n dt r a n s c r i p t i o n r e l a t i o n s h i p b y r e c o n s t r u c t i n g g e n e-l e v e l c h r o-m a t i n i n t e r a c t i o n[J].C o m p u t a t i o n a l a n d S t r u c t u r a l B i o t e c h-n o l o g y J o u r n a l,2019,17:195-205.[19] K H A L I D M,S A E E D-U R-R A HM A N,B I L A L M,e t a l.R o l eo f f l a v o n o i d s i n p l a n t i n t e r a c t i o n s w i t h t h e e n v i r o n m e n t a n da g a i n s t h u m a n p a t h o g e n s A r e v i e w[J].J o u r n a l o f I n t e-g r a t i v e A g r i c u l t u r e,2019,18(1):211-230.[20]周春然.平邑甜茶根系转录组分析及凋亡基因的表达[D].52519期刘媛媛,等:超重力胁迫下 平邑甜茶 幼苗叶片生理指标及其转录组分析山东泰安:山东农业大学,2016.[21] C L A A S S E N D E,S P O O N E R B S.I m p a c t o f A l t e r e d G r a v i t yo n A s p e c t s o f C e l l B i o l o g y[M]//I n t e r n a t i o n a l R e v i e w o f C y-t o l o g y-a S u r v e y o f C e l l B i o l o g y.A m s t e r d a m:E l s e v i e r, 1994:301-373.[22]高俊凤.植物生理学指导[M].北京:高等教育出版社.2015.[23]安钰,刘华,李明,等.外源钙对干旱胁迫下甘草生理特性的影响[J].中国现代中药,2019,21(10):1397-1401.A N Y,L I U H,L I M,e t a l.E f f e c t s o f e x o g e n o u s c a l c i u m o nt h e p h y s i o l o g i c a l c h a r a c t e r i s t i c s o f G l y c y r r h i z a u r a l e n s i s u n-d e r d r o u g h t s t r e s s[J].M o d e r n C h i n e s e M e d i c i n e,2019,21(10):1397-1401.[24]童琪,钟雁,李婧,等.不同温度对迷人杜鹃种子萌发与幼苗生长及生理特性的影响[J].西北植物学报,2020,40(3):471-477.T O N G Q,Z H O N G Y,L I J,e t a l.S e e d g e r m i n a t i o n,s e e d-l i n g g r o w t h a n d p h y s i o l o g i c a l c h a r a c t e r i s t i c s o f R h o d o d e b d r o na g a s t u m u n d e r d i f f e r e n t t e m p e r a t u r e s[J].A c t a B o t a n i c a B o-r e a l i-O c c i d e n t a l i a S i n i c a,2020,40(3):471-477.[25] T E R A S H I MA I,H A N B A Y T,T H O L E N D,e t a l.L e a ff u n c t i o n a l a n a t o m y i n r e l a t i o n t o p h o t o s y n t h e s i s[J].P l a n tP h y s i o l,2011,155:108-116.[26] T A K E MU R A K,K AMA C H I H,K UM E A,e t a l.A h y-p e r g r a v i t y e n v i r o n m e n t i n c r e a s e s c h l o r o p l a s t s i z e,p h o t o s y n-t h e s i s,a n d p l a n t g r o w t h i n t h e m o s s.P h y s c o m i t r e l l a p a t e n s[J]P l a n t R e s.,2017,130:181-192.[27] B U S H D R.P r o t o n-c o u p l e d s u g a r a n d a m i n o a c i d t r a n s p o r t-e r s i n p l a n t s[J].A n n u a l R e v i e w of P l a n t P h y s i o l og y a n dP l a n t M o l e c u l a r B i o l o g y,1993,44(1):513-542. [28] Z HA N G Y,S O G A K,WA K A B A Y A S H I K,e t a l.E f f e c t so f g r a v i s t i m u l i o n o s m o r e g u l a t i o n i n a z u k i b e a n e p i c o t y l s[J].A d v a n c e s i n S p a c e R e s e a r c h,2013,51(3):458-464.[29]S A N GWA N R S,T R I P A T H I S,S I N G H J,e t a l.D e n o v os e q u e n c i n g a n d a s s e m b l y o f C e n t e l l a a s i a t i c a l e a f t r a n s c r i p-t o m e f o r m a p p i n g o f s t r u c t u r a l,f u n c t i o n a l a n d r e g u l a t o r yg e n e s w i t h s p e c i a l r e f e r e n c e t o s e c o n d a r y m e t a b o l i s m[J].G e n e,2013,525(1):58-76.[30]王玉书,王欢,范震宇,等.基于转录组测序的羽衣甘蓝叶色相关基因分析[J].基因组学与应用生物学,2020,39(1):200-206.WA N G Y S,WA N G H,F A N Z Y,e t a l.I d e n t i f y i n g g e n e sa s s o c i a c t e d w i t h l e a f c o l o r i n k a l e(B r a s s i c a o l e r a c e a L.v a r.a c e p h a l a D C.)b a s e d o n t r a n sc r i p t o m e a n a l y s i s[J].G e n o m-i c s a n d A p p l i e d B i o l o g y,2020,39(1):200-206.[31]刘合霞,李博.紫花报春苣苔差异表达基因分析及离子转运蛋白基因挖掘[J].分子植物育种,2019,17(24):8029-8038.L I U H X,L I B.C o m p a r a t i v e t r a n s c r i p t o m e a n a l y s i s t o e x-p l o r e r e l a t e d g e n e s i n i o n t r a n s p o r t o f P r i m u l i n a p u r p u r e a[J].M o l e c u l a r P l a n t B r e e d i n g,2019,17(24):8029-8038.[32] P O S T N I K O V A O A,HU L T M,S H A O J,e t a l.T r a n s c r i p-t o m e a n a l y s i s o f r e s i s t a n t a n d s u s c e p t i b l e a l f a l f a c u l t i v a r s i n-f e c t e d w i t h r o o t-k n o t n e m a t o d e M e l o i d og y n e i n c o g n i t a[J].P L o S O n e,2015,10(2):e0118269.D O I:10.1371/j o u r n a l.p o n e.0118269.[33] C A L Z A D I L L A P I,MA I A L E S J,e t a l.T r a n s c r i p t o m e r e-s p o n s e m e d i a t e d b y c o l d s t r e s s i n L o t u s j a p o n i c u s[J].F r o n-t i e r s i n P l a n t S c i e n c e,2016,7:374.D O I:10.3389/f p l s.2016.00374.[34]S O G A K,WA K A B A Y A S H I K,e t a l.G r o w t h r e s t o r a t i o n i na z u k ib e a n a n d m a i z e s e e d l i n g s b y r e m o v a l o f h y p e r g r a v i t ys t i m u l i[J].A d v a n c e s i n S p a c e R e s e a r c h,2003,31(10): 2269-2274.[35]WA K A B A Y A S H I K,K AM I S A K A S,H O S O N T,e t a l.G r a v i p e r c e p t i o n i n g r o w t h i n h i b i t i o n o f p l a n t s h o o t s u n d e r h y-p e r g r a v i t y c o n d i t i o n s p r o d u c e d b y c e n t r i f u g a t i o n i s i n d e p e n d-e n t of t h a t i ng r a v i t r o p i s m a n d m a y i n v o l v e m e ch a n o r e c e p t o r s[J].P l a n t a,2004,218(6):1054-1061.[36] MA R T Z I V A N O U M,H AM P P R.H y p e r-g r a v i t y e f f e c t s o nt h e A r a b i d o p s i s t r a n s c r i p t o m e[J].P h y s i o l o g i a P l a n t a r u m, 2003,118(2):221-231.[37] WA K A B A Y A S H I K,S O G A K,e t a l.C h a n g e s i n l e v e l s o fc e l l w a l l c o n s t i t u e n t s i n w h e a t s e ed l i n g s g r o w n u n de r c o n t i n u-o u s h y p e r g r a v i t y c o n d i t i o n s[J].A d v a n c e s i n S p a c e R e-s e a r c h,2005,36(7):1292-1297.[38] MA R T Z I V A N O U M,H AM P P R.H y p e r-g r a v i t y e f f e c t s o nt h e A r a b i d o p s i s t r a n s c r i p t o m e[J].P h y s i o l o g i a P l a n t a r u m, 2003,118(2):221-231.[39] WA T S O N M B,E MO R Y K K,P I A T A K R M,e t a l.A r g i-n i n e d e c a r b o x y l a s e(p o l y a m i n e s y n t h e s i s)m u t a n t s o f A r a b i-d o p s i s t h a l i a n ae x h i b i t a l t e r e d r o o t g r o w t h[J].T h e P l a n tJ o u r n a l,1998,13(2):231-239.[40] F A R A O N I P,S E R E N I E,G N E R U C C I A,e t a l.G l y o x y l a t ec y c l e a c t i v i t y i n P i n u s p i n e a s e ed s d u r i n g ge r m i n a t i o n i n a l-t e r e d g r a v i t y c o n d i t i o n s[J].P l a n t P h y s i o l o g y a n d B i o c h e m-i s t r y,2019,139:389-394.[41] Y A N G B,J I A N G Y Q,e t a l.I d e n t i f i c a t i o n a n d e x p r e s s i o na n a l y s i s o f WR K Y t r a n s c r i p t i o n f a c t o r g e n e s i n c a n o l a(B r a s-s i c a n a p u s L.)i n r e s p o n s e t o f u n g a l p a t h o g e n s a n d h o r m o n e t r e a t m e n t s[J].B M C P l a n t B i o l o g y,2009,9(1):68. [42]栾倩倩.黄瓜C s WR K Y50基因的克隆与功能研究[D].山东泰安:山东农业大学,2018.[43]杨柳慧.细叶百合L p WR K Y20基因的克隆及其对烟草的遗传转化[D].哈尔滨:东北林业大学,2019.[44]J O N E S J D G,D A N G L J L.T h e p l a n t i mm u n e s y s t e m[J].N a t u r e,2006,444(7117):323-329.[45]王雅琼.苹果转录因子M d b Z I P26基因的抗逆功能研究[D].陕西杨陵:西北农林科技大学,2018.[46]李雪垠.小麦b Z I P转录因子家族中花药发育及抗逆相关基因的功能研究[D].陕西杨陵:西北农林科技大学,2016.[47] K O B A Y A S H I Y,M U R A T A M,M I N A M I H,e t a l.A b s c i s i ca c i d-a c t i v a t e d S N R K2p r o t e i n k i n a s e s f u n c t i o n i n t h e g e n e-r e g u l a t i o n p a t h w a y o f A B A s i g n a l t r a n s d u c t i o n b y p h o s p h o-r y l a t i n g A B A r e s p o n s e e l e m e n t-b i n d i n g f a c t o r s[J].T h eP l a n t J o u r n a l,2005,44(6):939-949.[48]张春霄.A P2/E R E B P家族M f E R F049基因对提高豆科植物生物胁迫和非生物胁迫抗性的功能分析[D].呼和浩特:内蒙古大学,2019.(编辑:裴阿卫)6251西北植物学报40卷。

干旱胁迫下平邑甜茶叶片差异表达蛋白分析杨小妮;王俊花;杨剑锋;祁香宁;段宇敏;韩明玉;张林森【期刊名称】《西北农林科技大学学报(自然科学版)》【年(卷),期】2018(046)011【摘要】[目的]探究平邑甜茶在干旱胁迫下的蛋白应激反应,为苹果砧木抗旱分子改良提供参考.[方法]以苹果砧木平邑甜茶为试材,在正常灌水后对其进行干旱胁迫处理(Mh-GD),以持续灌水为对照(Mh-CK),采用生理学和蛋白质组学相结合的研究方法,测定土壤水势、黎明前叶水势、光合作用及其叶片荧光参数,运用iTRAQ技术筛选鉴定响应干旱胁迫的差异表达蛋白,并对成功鉴定的蛋白进行GO注释及参与KEGG通路的分析.[结果]①与Mh-CK处理相比,Mh-GD处理平邑甜茶叶片相对含水量(RWC)、黎明前叶水势(ψleaf)、光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)均显著下降.②Mh-GD处理植株叶片PSⅡ的最大光合效率(Fv/Fm)、潜在活性(Fv/Fo)、性能指标(PI)、单位反应中心吸收的光能(ABS/RC)均降低,而荧光动力学曲线升高.③运用iTRAQ技术共鉴定出1 897个蛋白,其中181个蛋白质表达量在Mh-CK、Mh-GD处理叶片间有显著差异,上调蛋白81个(P<0.05,差异倍数>1.5),下调蛋白100个(P<0.05,差异倍数<0.67).181个蛋白中,86个蛋白功能已知,95个蛋白功能未知;已知功能可归为10类.[结论]在干旱胁迫下,平邑甜茶的差异表达蛋白质主要参与了光合作用、抗逆、信号转导和碳代谢等过程.【总页数】11页(P70-80)【作者】杨小妮;王俊花;杨剑锋;祁香宁;段宇敏;韩明玉;张林森【作者单位】西北农林科技大学园艺学院,陕西杨凌712100;西北农林科技大学园艺学院,陕西杨凌712100;西北农林科技大学园艺学院,陕西杨凌712100;西北农林科技大学园艺学院,陕西杨凌712100;西北农林科技大学园艺学院,陕西杨凌712100;西北农林科技大学园艺学院,陕西杨凌712100;西北农林科技大学园艺学院,陕西杨凌712100【正文语种】中文【中图分类】S661.1【相关文献】1.干旱胁迫下橡胶树叶片差异表达蛋白的鉴定与功能解析 [J], 林秀琴;袁坤;王真辉;邓军;杨礼富2.白羊草叶片和根系干旱胁迫下microRNAs差异表达分析 [J], 李春艳;董洁;钟华;董宽虎3.超重力胁迫下'平邑甜茶'幼苗叶片生理指标及其转录组分析 [J], 刘媛媛;申成丞;吕红柳;吴非;段国锋4.外源褪黑素对干旱胁迫下绥农26大豆鼓粒期叶片碳氮代谢调控的途径分析 [J], 曹亮;杜昕;于高波;金喜军;张明聪;任春元;王孟雪;张玉先5.外源甜菜碱对干旱胁迫下平邑甜茶叶片生理效应的影响 [J], 王贵平;王金政;薛晓敏;路超;聂佩显因版权原因,仅展示原文概要,查看原文内容请购买。


中国农业科学 2010,43(15):3176-3183Scientia Agricultura Sinica doi: 10.3864/j.issn.0578-1752.2010.15.015平邑甜茶叶片光合速率及叶绿素荧光参数对氯化镉处理的响应王 利1,2,杨洪强1,3,范伟国3,张 召2(1山东农业大学资源与环境学院农业资源利用博士后流动站,山东泰安 271018;2山东农业大学林学院农业生态与环境重点实验室,山东泰安 271018;3山东农业大学园艺科学与工程学院/作物生物学国家重点实验室,山东泰安 271018)摘要:【目的】研究氯化镉处理对平邑甜茶叶片光系统Ⅱ(PSⅡ)活性、光合速率影响及其相互关系,为进一步揭示镉伤害机理提供理论依据。
【方法】平邑甜茶在含不同浓度氯化镉1/2 Hoagland营养液中培养30 d后,测定其叶片光合速率(Pn)、气孔导度、胞间CO2浓度和荧光参数等,分析氯化镉处理后这些参数间的关系。
【结果】在氯化镉处理下,平邑甜茶叶片光合速率和气孔导度显著降低,胞间CO2浓度增加,300 μs时的叶绿素荧光强度(Fk)提高,PSⅡ最大光化学效率(Fv/Fm,φPo)、用于电子传递的量子产额(φEo)、光化学性能指数(PI ABS)以及有活性的反应中心的密度(RC/CS)明显下降,并且这些参数的变化幅度随着氯化镉浓度的增加而提高;通径分析显示,300 μs时的相对可变荧光强度(V K)及其可变荧光Fv占(J相的荧光强度Fj-O相的荧光强度Fo)振幅的比例(W K)对Pn的直接作用高于其它荧光参数。
【结论】氯化镉使平邑甜茶叶片PSⅡ供体侧、受体侧和反应中心受到显著伤害,从而降低了PSⅡ活性和光合速率;在氯化镉处理下,V K和W K对Pn的直接作用比较大。
关键词:平邑甜茶;氯化镉;光合速率;光系统Ⅱ;叶绿素荧光Effect of CdCl2 Treatment on PhotosyntheticRate and Chlorophyll Fluorescence Parameters inMalus hupehensis LeavesWANG Li 1,2, YANG Hong-qiang 1,3, FAN Wei-guo3, ZHANG Zhao2(1Post-Doctoral Mobile Station of Agricultural Resource Utilization, College of Resources and Environment, Shandong AgriculturalUniversity, Taian 271018, Shandong; 2Key Laboratory of Agricultural Ecology and Environment, College of Forestry, ShandongAgricultural University, Taian 271018, Shandong; 3State Key Laboratory of Crop Biology/College of Horticultural Science andEngineering, Shandong Agricultural University, Taian 271018, Shandong)Abstract: 【Objective】For discovering the mechanism of Cd damage on leaves of Malus hupehensis Rehd., the activity of photosystemⅡ (PSⅡ), net photosynthetic rate (Pn) and their correlation in leaves treated with CdCl2 were studied. 【Method】 After30 days of treatment by CdCl2 in 1/2 Hoagland solution, the Pn, stomatal conductance (Gs), intercellular CO2 concentration (Ci) andchlorophyll fluorescence parameters in leaves of Malus hupehensis Rehd. were measured, and the relationship between theseparameters under CdCl2 treatment were analyzed. 【Result】Under the treatment of CdCl2, the Pn and Gs reduced, the Ci and the fluorescence intensity Fk at 300 μs increased, and the maximum photochemistry efficiency of PSⅡ(Fv/Fm, φPo), the quantum yieldfor electron transport (φEo) , the performance index on absorption basis (PI ABS) and the density of active reaction center (RC/CS) alldecreased significantly. Furthermore, the range of variation of these parameters increased with the increasing of CdCl2 concentration.The direct effect of the relatively variable fluorescence intensity V K and the ratio of variable fluorescence Fv on the amplitude Fj-Fo(W K) at 300 μs for Pn were higher than that of others through the path analysis. 【Conclusion】 CdCl2 damaged the sides of acceptorand donor and the reaction centers of PSⅡ of leaves of Malus hupehensis Rehd. The activity of PSⅡand Pn decreased, and the direct收稿日期:2009-12-02;接受日期:2010-03-01基金项目:山东农业大学博士后项目、国家自然科学基金项目(30671452)作者简介:王利,副教授,博士。



砷、镉胁迫对茶树生育的影响唐茜;朱忻钰;李晓林;谭和平【期刊名称】《河南农业科学》【年(卷),期】2008(000)003【摘要】采用室内水培法,分别研究了砷(As)、镉(Cd)单一胁迫对茶树新梢生育和光合作用的影响以及毒害症状,结果表明:随着砷胁迫浓度的升高,茶树发芽和新梢生长受到严重抑制;茶树的叶绿素含量、光合速率、气孔导度和蒸腾速率均随砷胁迫浓度的升高而降低,同时各胁迫处理的茶树均出现明显的受害症状.镉胁迫的茶树在处理80 d后出现轻微毒害症状,低浓度镉胁迫(≤3 mg/L)对新梢生长、叶绿素合成和光合作用有一定促进作用;当胁迫浓度≥4 mg/L时,则表现负面影响.砷胁迫的毒害作用强于镉胁迫.【总页数】5页(P37-41)【作者】唐茜;朱忻钰;李晓林;谭和平【作者单位】四川农业大学,林学园艺学院,四川,雅安,625014;四川农业大学,林学园艺学院,四川,雅安,625014;四川农业大学,林学园艺学院,四川,雅安,625014;中国测试技术研究院,四川,成都,610021【正文语种】中文【中图分类】S571.1【相关文献】1.茶树新梢生育的内源激素水平及其调控机理*(第四报)外源激素对茶树内源激素的影响及其与新梢生长的关系 [J], 潘根生;钱利生;吴伯千;沈生荣2.铜与锌对茶树生育特性及生理代谢的影响Ⅳ.锌对茶树体内矿质元素含量及分布的影响 [J], 韩文炎3.铜与锌对茶树生育特性及生理代谢的影响Ⅲ.施铜对茶树体内铜及其他矿质元素吸收的影响 [J], 韩文炎4.茶树新梢生育的内源激素水平及其调控机理(第三报) 干旱胁迫对茶树内源激素的影响 [J], 潘根生;钱利生;吴伯千;沈生荣5.砷对茶树生长的影响及其在茶树体内的吸收与积累 [J], 朱忻钰;谭和平;叶善蓉;陈能武;张苏敏;唐茜因版权原因,仅展示原文概要,查看原文内容请购买。

㊀山东农业科学㊀2023ꎬ55(9):121~127ShandongAgriculturalSciences㊀DOI:10.14083/j.issn.1001-4942.2023.09.016收稿日期:2022-11-25基金项目:国家自然科学基金项目(32001988ꎬ32071788)ꎻ山东省自然科学基金项目(ZR2021MC102ꎬZR2020MC153)作者简介:吴康(1997 )ꎬ男ꎬ云南曲靖人ꎬ硕士研究生ꎬ研究方向为果树遗传育种ꎮE-mail:1760480049@qq.com通信作者:杨静静(1992 )ꎬ女ꎬ山东海阳人ꎬ博士ꎬ讲师ꎬ主要研究方向为果树育种学ꎮE-mail:ggyangjingjing@163.com宿红艳(1976 )ꎬ女ꎬ山东淄博人ꎬ博士ꎬ教授ꎬ主要研究方向为林木遗传育种ꎮE-mail:suhongyan66@126.com叶面喷施羧基化碳纳米管对平邑甜茶幼苗生长及抗氧化系统的影响吴康ꎬ苏安琪ꎬ田雪娇ꎬ李传收ꎬ付志顺ꎬ杨静静ꎬ宿红艳(鲁东大学农学院ꎬ山东烟台㊀264025)㊀㊀摘要:为研究碳纳米管(carbonnanotubesꎬCNTs)对植物生长发育的影响ꎬ本试验以苹果砧木平邑甜茶(Malushupehensis)为材料ꎬ设置叶面喷施浓度为0㊁10mg/L和20mg/L羧基化单壁碳纳米管(SWCNTs)处理ꎬ研究其对幼苗地上部生长和叶片光合作用㊁抗氧化系统等生理生化指标的影响ꎮ结果表明ꎬ叶面喷施10㊁20mg/LSWCNTs均能显著促进平邑甜茶幼苗地上部生长和叶片光合作用ꎬ有效增强叶片过氧化物酶(POD)㊁超氧化物歧化酶(SOD)㊁过氧化氢酶(CAT)㊁抗坏血酸过氧化物酶(APX)㊁谷胱甘肽还原酶(GR)和脱氢抗坏血酸还原酶(DHAR)活性ꎬ提高抗坏血酸(AsA)和谷胱甘肽(GSH)含量ꎮ与对照相比ꎬ喷施10㊁20mg/LSWC ̄NTs处理叶片脱氢抗坏血酸(DHA)和氧化型谷胱甘肽(GSSG)含量显著降低ꎬ而过氧化氢(H2O2)和丙二醛(MDA)含量没有显著变化ꎮ透射电镜结果显示ꎬSWCNTs能穿透并聚集在叶片细胞内部ꎮ综上ꎬ叶面喷施10㊁20mg/LSWCNTs能促进平邑甜茶地上部生长ꎬ提高叶片叶绿素含量和光合作用ꎬ上调抗氧化酶活性ꎬ并加快AsA-GSH循环系统的高效运转ꎬ使细胞保持较高的抗氧化能力ꎮ关键词:碳纳米管ꎻ平邑甜茶ꎻ叶片ꎻ抗氧化系统ꎻ光合作用中图分类号:S661.401㊀㊀文献标识号:A㊀㊀文章编号:1001-4942(2023)09-0121-07EffectsofFoliarApplicationofCarboxylatedCarbonNanotubesonGrowthandAntioxidantSystemofMalushupehensisWuKangꎬSuAnqiꎬTianXuejiaoꎬLiChuanshouꎬFuZhishunꎬYangJingjingꎬSuHongyan(CollegeofAgronomyꎬLudongUniversityꎬYantai264025ꎬChina)Abstract㊀Inordertostudytheeffectofcarbonnanotubes(CNTs)onplantgrowthanddevelopmentꎬtheapplerootstockMalushupehensiswasusedasexperimentalmaterialꎬandthefoliarapplicationof0ꎬ10and20mg/Lcarboxylatedsingle ̄walledcarbonnanotubes(SWCNTs)weresettostudytheireffectsontheabovegroundgrowthofseedlingsꎬleafphotosynthesisꎬantioxidantsystemandotherphysiologicalandbiochem ̄icalindexes.Theresultsshowedthatspraying10and20mg/LSWCNTssignificantlypromotedshootgrowthandleafphotosynthesisꎬeffectivelyenhancedtheactivitiesofperoxidase(POD)ꎬsuperoxidedismutase(SOD)ꎬcatalase(CAT)ꎬascorbateperoxidase(APX)ꎬglutathionereductase(GR)anddehydroascorbatereductase(DHAR)inleavesꎬandincreasedthecontentsofascorbicacid(AsA)andglutathione(GSH).Comparedwiththecontrolꎬthecontentsofdehydroascorbicacid(DHA)andoxidizedglutathione(GSSG)inleavessprayedwith10and20mg/LSWCNTsdecreasedsignificantlyꎬwhilethecontentsofhydrogenperoxide(H2O2)andmalondialdehyde(MDA)didnotchangesignificantly.TheresultsofTEMdetectionshowedthatSWCNTsparticlescouldpenetrateandaggregateinsideleafcells.Inconclusionꎬfoliarapplicationof10and20mg/LSWCNTscouldpromotethegrowthofM.hupehensisshootsꎬimprovethechlorophyllandphotosynthe ̄siscontentofleavesꎬenhancetheactivitiesofantioxidantenzymes(PODꎬSODandCAT)ꎬacceleratetheef ̄ficientoperationofASA ̄GSHcirculationsystemꎬandmaintainthehighantioxidantcapacityofcells.Keywords㊀CarbonnanotubesꎻMalushupehensisꎻLeavesꎻAntioxidantsystemꎻPhotosynthesis㊀㊀纳米材料和植物纳米生物技术是新兴的正处于发展中的国际前沿研究领域ꎬ其对世界经济发展以及人们的生产生活会产生非常重要的影响ꎮ碳纳米管(carbonnanotubesꎬCNTs)是一种工程碳基纳米材料ꎬ按照所含石墨烯片的层数可分为单壁碳纳米管(SWCNTs)和多壁碳纳米管(MWC ̄NTs)ꎮ目前ꎬCNTs已被广泛应用于航空航天㊁医疗㊁工业等领域[1-4]ꎮ随着纳米技术的发展ꎬCNTs在植物生物技术以及农业领域(例如纳米农药㊁纳米肥料㊁纳米杀虫剂或纳米仿生)的应用受到越来越多的关注[5-6]ꎮ因此ꎬ探究CNTs对植物生长发育的影响及其在植物生长发育中的响应机制ꎬ对促进其更好地应用于农业生产和作物管理具有重要意义ꎮ目前国内外已经开展过一系列关于CNTs影响植物生长发育的研究ꎮ现阶段有关CNTs对植物生长影响的研究大致持两种观点:一部分研究认为CNTs对植物生长有促进作用或者没有明显毒性作用ꎬ另一部分研究则认为CNTs能够通过影响植物根系水分吸收㊁光合作用等过程抑制种子萌发和幼苗生长ꎮCañas等[7]研究表明ꎬ相对低剂量的SWCNTs可以穿透种皮ꎬ促进番茄幼苗生长ꎻ相反地ꎬStampoulis等[8]的研究表明ꎬ1000mg/LMWCNTs会诱导细胞产生大量活性氧(ROS)并损伤抗氧化系统ꎬ扰乱正常代谢及信号转导过程导致其生物量降低ꎮ此外ꎬ有研究表明适宜浓度的CNTs可以调节细胞内抗氧化酶(如SOD㊁POD和CAT)活性ꎬ进而提高植物对盐胁迫的耐受性[9-11]ꎮ可以看出ꎬCNTs对植物生长的利弊与其自身特性㊁环境条件及植物种类密切相关ꎮ虽然目前关于CNTs对作物生长发育的影响已有大量研究报道ꎬ但受试物种主要集中在拟南芥㊁番茄㊁小麦㊁红波菜和水稻等模式植物及禾本科植物上ꎬ而关于CNTs对木本植物苹果生长影响的研究尚未见报道ꎮ鉴于此ꎬ本试验以苹果砧木平邑甜茶幼苗为材料ꎬ测定分析不同浓度(0㊁10㊁20mg/L)羧基化单壁碳纳米管(SWCNTs)对平邑甜茶幼苗生长㊁叶片光合作用㊁叶绿素含量以及抗氧化酶活性的影响ꎬ旨在探讨SWCNTs对平邑甜茶幼苗生长的影响和作用机理ꎬ以期为CNTs在环境和农业生产上的安全使用和处理处置提供一定的科学依据ꎮ1㊀材料与方法1.1㊀试验材料试验于2020 2021年在鲁东大学农学院试验基地进行ꎮ2021年2月1日挑选饱满一致的平邑甜茶种子ꎬ用0.2%高锰酸钾消毒30minꎬ流水冲洗12hꎬ然后置于4ħ下沙藏40天ꎮSWC ̄NTs干粉购自中国科学院成都有机化学有限公司(http://www.cioc.ac.cn/)ꎬ产品具体参数见表1ꎮ㊀㊀表1㊀羧基化单壁碳纳米管(SWCNTs)参数SWCNTs参数参数值外径/mm1~2纯度/W%>90羧基(-COOH)含量/W%2.73长度/μm5~30比表面积/(m2/g)>320灰分/W%<51.2㊀试验设计与处理2021年3月中旬选择露白一致的平邑甜茶种子播种于穴盘(540mmˑ280mm)中ꎬ其育苗基质为蛭石与营养土混合(1ʒ1)基质ꎬ待长至4片真叶时移栽到培养钵(7cmˑ7cm)中ꎬ之后进行正常肥水管理ꎮ称取一定量的SWCNTs粉末加入到去离子水中ꎬ超声分散(100W㊁40Hz)1hꎬ配制成不同浓度(0㊁10mg/L和20mg/L)SWCNTs悬浮液ꎮ待植株长至8片真叶时ꎬ选择长势一致的幼苗ꎬ分为3组ꎬ以15棵苗为1组ꎬ叶面喷施0㊁10mg/L和20mg/LSWCNTs悬浮液ꎬ分别记为CNTs0㊁CNTs10和CNTs20处理ꎮ每个处理独立重复3次ꎮ约20天后ꎬ植株表型出现明显差异ꎬ收集暴露于SWCNTs的植株叶片以供指标分析ꎮ221㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第55卷㊀1.3㊀测定项目及方法1.3.1㊀生物量测定㊀处理20天时ꎬ每组随机选取5株平邑甜茶幼苗ꎬ用米尺测定株高ꎻ取地上部茎叶用分析天平称取鲜重ꎬ之后放入烘箱内ꎬ105ħ杀青30min后70ħ连续烘干72hꎬ至恒重时记为干重ꎮ1.3.2㊀光合速率及叶绿素含量测定㊀选择上午9ʒ00 11ʒ00ꎬ用LI-6400光合测定仪测定植株功能叶的光合速率ꎮ叶绿素含量测定采用分光光度法[12]ꎬ即利用95%乙醇提取色素ꎬ分别测定665㊁649nm处的OD值ꎬ计算叶绿素a㊁叶绿素b和叶绿素总含量ꎮ1.3.3㊀抗氧化酶活性和其它理化指标测定㊀超氧化物歧化酶(SOD)活性采用氮蓝四唑(NBT)法测定[13]ꎻ过氧化物酶(POD)活性采用愈创木酚法测定[14]ꎻ过氧化氢酶(CAT)活性采用紫外分光光度法测定[15]ꎮ丙二醛(MDA)含量采用硫代巴比妥酸法测定[16]ꎻ过氧化氢(H2O2)含量采用TiCl4沉淀法测定[17]ꎮ1.3.4㊀AsA-GSH循环系统指标测定㊀抗坏血酸(AsA)和脱氢抗坏血酸(DHA)含量参照Ma等[18]的方法测定ꎮ谷胱甘肽(GSH)和氧化型谷胱甘肽(GSSG)含量参照Anderson等[19]的方法测定ꎮ抗坏血酸过氧化物酶(APX)活性的测定[20]ꎬ监测APX氧化AsA引起的290nm处光吸收值的下降ꎮ脱氢抗坏血酸还原酶(DHAR)活性的测定[21]ꎬ监测由AsA的生成引起的265nm处吸光值的上升ꎮ谷胱甘肽还原酶(GR)活性的测定[22]ꎬ监测由NADPH氧化所引起的340nm处吸光值的下降ꎮ1.3.5㊀透射电子显微镜(TEM)观察SWCNTs在叶片细胞中的分布㊀将对照和暴露于SWCNTs的植株叶片放于固定液中ꎬ用手术刀切割组织取样1mm3左右ꎬ经脱水㊁包埋㊁切片㊁染色后于透射电子显微镜下观察ꎬ并采集图像分析ꎮ1.4㊀数据处理与分析试验数据采用MicrosoftExcel2019和IBMSPSSStatistics25.0软件进行统计分析和作图ꎬ用One-wayANOVA方法进行Duncan s检验(P<0.05)ꎬ数据取平均值ʃ标准差(n=3ꎬ3个生物学重复)ꎮ2㊀结果与分析2.1㊀叶面喷施SWCNTs对平邑甜茶幼苗地上部生长的影响如图1所示ꎬ叶面喷施SWCNTs可显著影响平邑甜茶幼苗株高㊁地上部鲜重和干重ꎮ与对照相比ꎬ不同浓度SWCNTs处理下ꎬ平邑甜茶幼苗株高㊁地上部鲜重和干重均有不同程度提高ꎬ其中10mg/LSWCNTs处理下平邑甜茶幼苗株高显著增高21.2%ꎬ地上部鲜重显著增加25.7%ꎮ柱上不同小写字母表示处理间差异显著(P<0.05)ꎬ下同ꎮ图1㊀不同浓度SWCNTs处理下平邑甜茶幼苗表型2.2㊀叶面喷施SWCNTs对平邑甜茶幼苗叶片光合作用和叶绿素含量的影响图2显示ꎬ与对照相比ꎬ叶面喷施SWCNTs显著提高平邑甜茶幼苗叶片的净光合速率ꎬ其中10㊁20mg/LSWCNTs处理净光合速率分别提高10.9%和12.3%ꎮ随着SWCNTs浓度升高ꎬ叶绿素a㊁叶绿素b和叶绿素总含量均有不同程度提高ꎬ其中ꎬ20mg/LSWCNTs处理叶绿素a含量提高11.7%ꎬ叶绿素b含量提高16.4%ꎬ叶绿素总含量提高13.3%ꎮ2.3㊀叶面喷施SWCNTs对平邑甜茶幼苗叶片MDA和H2O2含量的影响如图3所示ꎬ与对照相比ꎬ喷施10㊁20mg/LSWCNTs处理平邑甜茶幼苗叶片的丙二醛(MDA)和过氧化氢(H2O2)含量略微增加ꎮ这表明该浓度范围内SWCNTs对平邑甜茶幼苗叶片没有产生明显的毒性作用ꎮ321㊀第9期㊀㊀㊀㊀㊀吴康ꎬ等:叶面喷施羧基化碳纳米管对平邑甜茶幼苗生长及抗氧化系统的影响图2㊀不同浓度SWCNTs对平邑甜茶幼苗叶片光合作用的影响图3㊀不同浓度SWCNTs对平邑甜茶幼苗㊀㊀叶片MDA和H2O2含量的影响2.4㊀叶面喷施SWCNTs对平邑甜茶幼苗叶片抗氧化酶活性的影响如图4所示ꎬ叶面喷施SWCNTs可提高平邑甜茶幼苗叶片SOD㊁POD㊁CAT㊁DHAR㊁APX㊁GR活性ꎮSWCNTs处理组平邑甜茶幼苗叶片SOD㊁CAT活性显著高于对照ꎬ其中10mg/LSWCNTs处理叶片CAT活性较对照提高23.7%(P<0.05)ꎬ20mg/LSWCNTs处理叶片SOD㊁POD㊁CAT活性较对照分别提高25.5%㊁11.9%和19.6%(P<0.05)ꎮ叶面喷施SWCNTs后平邑甜茶幼苗叶片APX㊁DHAR㊁GR活性均有不同程度的升高ꎬ与对照相比ꎬ10mg/LSWCNTs处理叶片APX㊁DHAR㊁GR活性分别提高28.9%㊁44.8%和16.6%(P<0.05)ꎬ但10㊁20mg/LSWCNTs处理之间DHAR和GR活性无显著差异ꎮ2.5㊀叶面喷施SWCNTs对平邑甜茶幼苗叶片非酶抗氧化物质含量的影响如图5所示ꎬ与对照相比ꎬ10mg/LSWCNTs处理下ꎬ平邑甜茶幼苗叶片AsA㊁GSH含量分别提高26.1%和18.2%(P<0.05)ꎬ而DHA㊁GSSG含量分别降低21.9%和10.5%(P<0.05)ꎬAsA/DHA和GSH/GSSG的比值分别升高49.8%和49.3%ꎮ图4㊀不同浓度SWCNTs对平邑甜茶幼苗㊀㊀叶片抗氧化酶活性的影响图5㊀不同浓度SWCNTs对平邑甜茶幼苗叶片㊀㊀非酶抗氧化物质含量的影响421㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第55卷㊀2.6㊀SWCNTs在平邑甜茶幼苗叶片中的分布为了探究SWCNTs在平邑甜茶幼苗叶片中的吸收和积累情况ꎬ利用透射电子显微镜(TEM)对平邑甜茶叶片进行观察ꎮ如图6所示ꎬ经SWC ̄NTs处理的叶片细胞中分布着黑色颗粒团聚体ꎬ而对照叶片细胞中没有黑色束或团聚体ꎮ这一结果表明ꎬSWCNTs能被平邑甜茶叶片吸收ꎮ图6㊀对照(A)和20mg/LSWCNTs处理(B)平邑甜茶幼苗叶片透射电镜(TEM)结果3㊀讨论与结论碳纳米管(CNTs)对植物生长的积极影响及其潜在机制先前已有报道ꎮ例如ꎬSeddighinia等[23]研究发现50mg/LMWCNTs促进了苦瓜根系和茎的伸长ꎮ我们最近的研究表明ꎬ土壤浇灌50㊁100mg/LSWCNTs显著上调杨树茎中纤维素和木质素合成酶(CesA㊁PAL和C4H)的活性ꎬ促进杨树茎中纤维素和木质素的积累[24]ꎮ本研究中ꎬ叶片喷施10㊁20mg/LSWCNTs促进平邑甜茶幼苗地上部的生长ꎮ光合作用在植物生命活动中具有重要意义ꎬ纳米材料的应用有助于提高植物光合速率ꎬ促进植物生长[25]ꎮZhang等[26]发现ꎬ20mg/LSWC ̄NTs增加水稻叶片叶绿素含量ꎬ促进水稻幼苗叶片的光合作用及其生长发育ꎬ显著上调水稻叶绿体发育所必需的VIRESCENT2和YGL138(t)基因的转录水平ꎮ本试验中ꎬ叶片喷施10㊁20mg/LSWCNTs显著提高叶片叶绿素a㊁叶绿素b和总叶绿素含量ꎮ光合色素可以通过清除ROS保护叶绿体ꎬ从而减少叶片中的氧化损伤[27]ꎮ叶绿素含量增加表明平邑甜茶幼苗叶片叶绿体保护机制可能被SWCNTs激活ꎮ最近的几项研究表明ꎬSWC ̄NTs可以有效地附着在植物光合反应中心的蛋白质上并与光合反应中心交换电子ꎬ提高电子传递的阈值以及光能利用效率ꎬ将光合作用活性提高3倍以上[28-30]ꎮ相反地ꎬ另一项研究表明ꎬ高剂量SWCNTs(300mg/L)会引起光系统Ⅱ(PSⅡ)反应中心失活以及光系统Ⅰ(PSⅠ)末端的受体减少ꎬ导致叶绿体基粒和基质区的膨胀ꎬ显著降低净光合速率[30]ꎮ本研究结果显示ꎬSWCNTs能被平邑甜茶幼苗叶片吸收ꎬ但其是否能够附着在叶绿体表面以及是否与叶绿体进行电子交换尚不确定ꎮ因此ꎬ平邑甜茶幼苗叶片增加的光合活性是由于SWCNTs和叶绿体之间电子转移的增加还是叶绿素含量增加所致仍需要进一步的探索ꎮ许多研究表明ꎬ碳基纳米材料可作为非生物激发因子诱导植物发生一定程度的氧化胁迫ꎬ但是植物可通过启动体内抗氧化系统ꎬ包括抗氧化物质(AsA和GSH)以及抗氧化酶(POD㊁SOD㊁CAT㊁APX㊁GR和DHAR)ꎬ响应CNTs造成的氧化胁迫[31-34]ꎮ本研究结果显示ꎬ10㊁20mg/LSWCNTs均能促进平邑甜茶幼苗叶片POD㊁SOD㊁CAT㊁APX㊁GR㊁DHAR活性升高ꎬ这与CNTs处理苹果无菌苗㊁蚕豆和鼠尾草等植物中的结果相一致[35-37]ꎮ此外ꎬ10㊁20mg/LSWCNTs处理提高平邑甜茶幼苗叶片AsA㊁GSH含量ꎬ一定程度上降低DHA㊁GSSG含量ꎬ从而显著提高AsA/DHA㊁GSH/GSSG比值ꎮ较高的AsA/DHA和GSH/GSSG比值使抗氧化物质维持在较高的还原态ꎬ能够直接猝灭ROS[38-39]ꎮ本研究中ꎬ10㊁20mg/LSWCNTs处理下平邑甜茶幼苗叶片H2O2㊁MDA含量比对照略高ꎬ但差异不显著ꎬ表明抗氧化系统能正常清除ROSꎮ综上所述ꎬ叶面喷施SWCNTs能提高叶片叶绿素含量和光合作用ꎬ促进平邑甜茶地上部生长ꎬ上调抗氧化酶活性ꎬ加快ASA-GSH循环系统的高效运转ꎬ使植物细胞保持较高的抗氧化能力ꎮ521㊀第9期㊀㊀㊀㊀㊀吴康ꎬ等:叶面喷施羧基化碳纳米管对平邑甜茶幼苗生长及抗氧化系统的影响参㊀考㊀文㊀献:[1]㊀BaughmanRHꎬZakhidovAAꎬdeHeerWA.Carbonnano ̄tubes theroutetowardapplications[J].Scienceꎬ2002ꎬ297(5582):787-792.[2]㊀FakruddinMꎬHossainZꎬAfrozH.Prospectsandnanobiotechn ̄ology:amedicalperspective[J].JournalofNanobiotechnologyꎬ2012ꎬ10(1):31.[3]㊀Herrero ̄LatorreCꎬÁlvarez ̄MéndezJꎬBarciela ̄GarcíaJꎬetal.Characterizationofcarbonnanotubesandanalyticalmethodsfortheirdeterminationinenvironmentalandbiologicalsamples:areview[J].AnalyticaChimicaActaꎬ2015ꎬ853:77-94. [4]㊀FuLꎬWangZYꎬDhankherOPꎬetal.Nanotechnologyasanewsustainableapproachforcontrollingcropdiseasesandincreas ̄ingagriculturalproduction[J].JournalofExperimentalBota ̄nyꎬ2020ꎬ71:507-519.[5]㊀DeVolderMFLꎬTawfickSHꎬBaughmanRHꎬetal.Carbonnanotubes:presentandfuturecommercialapplications[J].Sci ̄enceꎬ2013ꎬ339(6119):535-539.[6]㊀MukherjeeAꎬMajumdarSꎬServinADꎬetal.Carbonnanomate ̄rialsinagriculture:acriticalreview[J].FrontiersinPlantScienceꎬ2016ꎬ7:172.[7]㊀CañasJEꎬLongMꎬNationsSꎬetal.Effectsoffunctionalizedandnonfunctionalizedsingle ̄walledcarbonnanotubesonrootelongationofselectcropspecies[J].EnvironmentalToxicologyandChemistryꎬ2008ꎬ27(9):1922-1931.[8]㊀StampoulisDꎬSinhaSKꎬWhiteJC.Assay ̄dependentphytotox ̄icityofnanoparticlestoplants[J].EnvironmentalScienceandTechnologyꎬ2009ꎬ43(24):9473-9479.[9]㊀KhanMNꎬMobinMꎬAbbasZKꎬetal.Roleofnanomaterialsinplantsunderchallengingenvironments[J].PlantPhysiologyandBiochemistryꎬ2017ꎬ110:194-209.[10]PandeyKꎬLahianiMHꎬHicksVKꎬetal.Effectsofcarbon-basednanomaterialsonseedgerminationbiomassaccumulationandsaltstressresponseofbioenergycrops[J].PLoSONEꎬ2018ꎬ13(8):e0202274.[11]ZulfiqarFꎬAshrafM.Nanoparticlespotentiallymediatesaltstresstoleranceinplants[J].PlantPhysiologyandBiochemis ̄tryꎬ2021ꎬ160:257-268.[12]张以顺ꎬ黄霞ꎬ陈云凤.植物生理学实验教程[M].北京:高等教育出版社ꎬ2009.[13]赵世杰ꎬ史国安ꎬ董新纯.植物生理学实验指导[M].北京:中国农业科学技术出版社ꎬ2002.[14]李玲.植物生理学模块实验指导[M].北京:科学出版社ꎬ2009.[15]BeauchampCꎬFridovichI.Superoxidedismutaseimprovedas ̄saysandanassayapplicabletoacrylamidegels[J].AnalyticalBiochemistryꎬ1971ꎬ44(1):276-287.[16]HammerschmidtRꎬNucklesEMꎬKuc'J.Associationofen ̄hancedperoxidaseactivitywithinducedsystemicresistanceofcucumbertoColletotrchumlagenarium[J].PhysiologicalPlantPathologyꎬ1982ꎬ20(1):73-82.[17]CakmakIꎬMarschnerH.Magnesiumdeficiencyandhighlightintensityenhanceactivitiesofsuperoxidedismutaseꎬascorbateperoxidaseꎬandglutathionereductaseinbeanleaves[J].PlantPhysiologyꎬ1992ꎬ98(4):1222-1227.[18]MaYYꎬHuangDDꎬChenCBꎬetal.Regulationofascorbat ̄eglutathionecycleinpeachesvianitricoxidetreatmentduringcoldstorage[J].ScientiaHorticulturaeꎬ2019ꎬ247:400-406. [19]AndersonME.Determinationofglutathioneandglutathionedi ̄sulfideinbiologicalsamples[J].MethodsEnzymolꎬ1985ꎬ113:548-555.[20]NakanoYꎬAsadaK.Hydrogenperoxideisscavengedbyascor ̄bate-specificperoxidaseinspinachchloroplasts[J].PlantandCellPhysiologyꎬ1981ꎬ22(5):867-880.[21]ShanCJꎬZhangYYꎬZhangHX.ABAparticipatesinthereg ̄ulationofvitaminCcontentinthefruitofstrawberryusinglan ̄thanumnitrate[J].ScientiaHorticulturaeꎬ2018ꎬ233:455-459.[22]HodgesDMꎬForneyCF.Theeffectsofethyleneꎬdepressedox ̄ygenandelevatedcarbondioxideonantioxidantprofilesofse ̄nescingspinachleaves[J].JournalofExperimentalBotanyꎬ2000ꎬ51:645-655.[23]SeddighiniaFSꎬIranbakhshAꎬArdebiliZOꎬetal.Seedprim ̄ingwithcoldplasmaandmulti ̄walledcarbonnanotubesmodi ̄fiedgrowthꎬtissuedifferentiationꎬanatomyꎬandyieldinbittermelon(Momordicacharantia)[J].JournalofPlantGrowthRegulationꎬ2020ꎬ39(1):87-98.[24]LiMJꎬWangLꎬZhangJꎬetal.Single ̄walledcarbonnanotubespromoteswoodformationinPopulusdavidianaˑP.bolleana[J].PlantPhysiologyandBiochemistryꎬ2022ꎬ184:137-143. [25]LiuYLꎬYueLꎬWangZYꎬetal.Processesandmechanismsofphotosynthesisaugmentedbyengineerednanomaterials[J].EnvironmentalChemistryꎬ2019ꎬ16(6):430-445. [26]ZhangHꎬYueMXꎬZhengXKꎬetal.Physiologicaleffectsofsingle ̄andmulti ̄walledcarbonnanotubesonriceseedlings[J].IEEETransonNanobioscienceꎬ2017ꎬ16(7):563-570. [27]MittlerR.Oxidativestressꎬantioxidantsandstresstolerance[J].TrendsinPlantScienceꎬ2002ꎬ7:405-410.[28]DorogiMꎬBalintZꎬMikóCꎬetal.Stabilizationeffectofsingle ̄walledcarbonnanotubesonthefunctioningofphotosyntheticreactioncenters[J].TheJournalofPhysicalChemistryBꎬ2006ꎬ110:21473-21479.[29]MackowskiS.Hybridnanostructuresforefficientlightharves ̄ting[J].JournalofPhysicsCondensedMatterꎬ2010ꎬ22(19):193102.[30]VelikovaVꎬPetrovaNꎬKovácsLꎬetal.Single ̄walledcarbonnanotubesmodifyleafmicromorphologyꎬchloroplastultrastruc ̄tureandphotosyntheticactivityofpeaplants[J].International621㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第55卷㊀JournalofMolecularSciencesꎬ2021ꎬ22(9):4878. [31]袁刚强ꎬ龚继来ꎬ曾光明.单壁碳纳米管材料对水稻幼苗的毒性效应[J].环境科学学报ꎬ2015ꎬ35(12):4143-4149. [32]MarslinGꎬSheebaCJꎬFranklinG.Nanoparticlesalterseconda ̄rymetabolisminplantsviaROSburst[J].FrontiersinPlantScienceꎬ2017ꎬ8:00832.[33]VermaSKꎬDasAKꎬGantaitSꎬetal.Applicationsofcarbonnanomaterialsintheplantsystem:aperspectiveviewontheprosandcons[J].ScienceofTheTotalEnvironmentꎬ2019ꎬ667:485-499.[34]VermaSKꎬDasAKꎬPatelMKꎬetal.Engineerednanomateri ̄alsforplantgrowthanddevelopment:aperspectiveanalysis[J].ScienceofTheTotalEnvironmentꎬ2018ꎬ630:1413-1435.[35]刘海涛ꎬ汪承润ꎬ刘玲ꎬ等.羧基化多壁碳纳米管与铅镉对蚕豆幼苗叶片的氧化损伤[J].生态与农村环境学报ꎬ2018ꎬ34(8):745-754.[36]王鸿雪.碳纳米管处理对嘎啦叶片再生及无菌苗生根的影响[D].南京:南京农业大学ꎬ2020.[37]RahmaniNꎬRadjabianTꎬSoltaniBM.Impactsoffoliarexpo ̄suretomulti ̄walledcarbonnanotubesonphysiologicalandmo ̄leculartraitsofSalviaverticillataL.ꎬasamedicinalplant[J].PlantPhysiologyandBiochemistryꎬ2020ꎬ150:27-38. [38]WangPꎬYinLHꎬLiangDꎬetal.Delayedsenescenceofappleleavesbyexogenousmelatonintreatment:towardregulatingtheascorbate ̄glutathionecycle[J].JournalofPinealResearchꎬ2012ꎬ53(1):11-20.[39]汪宽鸿ꎬ祝彪ꎬ朱祝军.GSH/GSSG在植物应对非生物胁迫中的作用综述[J].园艺学报ꎬ2021ꎬ48(4):647-660.721㊀第9期㊀㊀㊀㊀㊀吴康ꎬ等:叶面喷施羧基化碳纳米管对平邑甜茶幼苗生长及抗氧化系统的影响。

重金属镉对茶树生理特性的影响作者:唐迪徐晓燕李树炎蒋新宇来源:《湖北农业科学》2013年第12期摘要:以二年生福鼎大白茶为试验材料,采用盆栽试验方法,探讨了重金属镉胁迫对茶树生理特性的影响。
结果表明,在试验设置的镉胁迫浓度范围(5~40 mg/kg)内,镉在茶树叶片中的含量随着镉胁迫浓度的增加而提高,反映出茶树对镉具有富集特性。
在生理指标方面,叶片可溶性蛋白含量随着镉胁迫浓度的增加而降低;20 mg/kg以下浓度的镉胁迫能提高茶树叶片的超氧化物歧化酶活性,10 mg/kg以下浓度的镉胁迫能增加茶树叶片的丙二醛、叶绿素、类胡萝卜素的含量;而超过上述镉胁迫浓度后则降低了超氧化物歧化酶活性,减少了丙二醛、叶绿素、类胡萝卜素的含量,并且影响了茶树的正常生长。
这个结果可为茶园生产中镉毒害的田间预报、监测指标的确认、数据可信度辨析、污染程度评价等方面提供有价值的参考。
关键词:镉;茶树;胁迫生理中图分类号:O614.24+2;S571.1;Q945.78 文献标识码:A 文章编号:0439-8114(2013)12-2839-05Effect of Heavy Metal Cadmium on Physiological Characteristics of Tea PlantTANG Di,XU Xiao-yan,LI Shu-yan,JIANG Xin-yu(Jiangsu Agricultural and Forestry Vocational Technical College,Zhenjiang 212400,Jiangsu,China)Abstract: Using 2-year-old Camellia sinensis (L.) O. Kuntze cv. Fuding Dabaicha as test material, pot method was conducted to explore the effects of heavy metal cadmium stress on physiological characteristics of tea plant. Results showed that in the setting range of cadmium concentration (5~40 mg/kg), cadmium content in tea plant leaves increased with the increasing of cadmium concentration, showing that tea plant could enrich cadmium. In terms of physiological indicators, leaf soluble protein content decreased with the increase of cadmium concentration;cadmium concentration under 20 mg/kg increased superoxide dismutase(SOD) activity in tea plants leaves, cadmium concentration under 10 mg/kg increased chlorophyll, Malondialdehyde (MDA)and carotenoid content. However, higher cadmium concentration decreased SOD activity and MDA, chlorophyll and carotenoid content, and affected normal growth of tea plant. The results could provide valuable reference for field forecasting,identification of monitoring indicators,discrimination of data credibility, assessment of pollution degree in tea garden.Key words: cadmium; tea plant[Camellia sinensis (L.) O. Kuntze]; physiological characteristics镉(Cadmium,Cd)是一种生物非必需的重金属元素,其化合物对人类、动物、植物等生物有很强的毒性[1]。

水分胁迫对茶树叶片叶绿素荧光影响的实验报告本文旨在描述水分胁迫对茶树叶片叶绿素荧光的影响。
实验使用了光合作用测量仪来测量叶片的叶绿素荧光,并进行数据处理和分析。
结果表明,水分胁迫可以显著影响茶树叶片的叶绿素荧光特性。
引言茶树(Camellia sinensis)是一种重要的园艺作物,广泛种植于亚洲和其他地区。
茶树叶片中的叶绿素是进行光合作用的重要物质之一,同时也是研究茶树生长和抗逆性的关键指标。
由于环境因素的变化,茶树叶片的叶绿素荧光特性会受到影响,而水分胁迫是其中最常见的一个因素。
因此,了解水分胁迫对茶树叶片的叶绿素荧光的影响,对于茶树生长和抗逆性的研究具有重要意义。
材料和方法1. 实验材料茶树样本:在沙土中种植的茶树,分别处于正常浇水和水分胁迫处理下。
2. 实验仪器光合作用测量仪:用于测量叶片叶绿素荧光。
3. 实验步骤1)在处理茶树样本生长的条件下进行三天的适应期。
2)将茶树样本分为两组,一组正常浇水,另一组进行水分胁迫处理,连续处理七天。
3)在实验过程中记录茶树样本的生长情况,包括叶片颜色、形态和生长速度等方面。
5)进行数据处理和统计分析。
结果1. 叶片表现在实验中,处于正常浇水条件下的茶树叶片表现出绿色,生长良好,茶树正常生长的情况。
而另一组茶树样本,由于持续的水分胁迫,其叶片表现出较明显的黄色或枯黄色,树冠部位叶片的生长速度较慢,部分叶片出现萎缩和干枯现象。
2. 叶绿素荧光数据使用光合作用测量仪测量的数据表明,处于水分胁迫条件下的茶树叶片,其最大光化学效率(Fv/Fm)显著低于正常浇水处理茶树叶片。
在7天的实验中,正常浇水的茶树样本的Fv/Fm值稳定在0.85左右,而水分胁迫处理下的茶树样本Fv/Fm值下降至0.60左右。
同时,水分胁迫处理下的茶树叶片,其最大光化学效率值的波动性较大,而正常浇水下的茶树样本Fv/Fm值波动性较小。
讨论通过本实验的结果,可以发现水分胁迫对茶树叶片叶绿素荧光特性影响较大。
不同苹果砧木叶绿素含量与光合速率研究作者:宋莎江旭生韩秀梅等来源:《江苏农业科学》2015年第03期摘要:测定10个苹果砧木的叶绿素含量、叶面积和净光合速率(Pn),并进行相关性分析。
结果表明,不同苹果砧木叶绿素含量、叶绿素a/叶绿素b、叶面积、净光合速率存在明显差异,其中平邑甜茶的叶绿素含量最高,叶面积最大;八棱海棠的叶绿素a/叶绿素b值最大;花红的净光合速率最高。
苹果砧木叶绿素含量与净光合速率呈显著正相关。
关键词:苹果砧木;叶绿素含量;净光合速率中图分类号: Q945.11 文献标志码: A文章编号:1002-1302(2015)03-0149-02苹果(Malus pumila Mill.)是我国主要水果之一,栽培面积和总产量均居世界第一位。
苹果生产中主要采用砧木嫁接繁殖,砧木可以提高栽培品种的抗逆性,不同的砧木对于果树生长发育和品质均会产生影响[1-3]。
生产中常用砧木有八棱海棠、平邑甜茶等,贵州省拥有丰富的苹果野生砧木资源,但迄今为止对其生理特性评价等研究工作较少,致使这些资源未能充分开发利用。
叶片的光合作用是果树碳水化合物的重要来源,研究果树光合作用与叶片形态及生理特性的关系,也是栽培生理研究主要内容之一。
目前在果树方面对叶绿素含量与光合速率的关系有较多研究报道,但以苹果砧木为材料的研究鲜有报道[4-6]。
本试验测定了10份苹果砧木中叶绿素含量、叶面积及光合速率,并探讨相互间的相关性,以期为苹果砧木的利用提供理论依据。
1 材料与方法1.1 材料本试验材料为10种(花红、八棱海棠、平邑甜茶、青砧1号、青砧2号、垂丝海棠、毛山荆子、湖北海棠、西府海棠、丽江山荆子)苹果砧木实生苗,其中八棱海棠、平邑甜茶、青砧1号和青砧2号种子由烟台果树科学研究所提供,其余采自贵州省威宁县。
于2012年3月播种,选取生长一致的实生苗用于后续试验。
1.2 方法试验于2013年7月进行,选取生长势一致的相似植株,从顶端向下第5张健康叶用于光合作用、叶面积和叶绿素含量测定。
土壤紧实度对甜茶幼苗光合参数和水分利用率的影响吴亚维;马锋旺;邹养军【期刊名称】《贵州农业科学》【年(卷),期】2009(037)004【摘要】以平邑甜茶(Malus hupehensis.Rehd)幼苗为材料,采用盆栽模拟试验,测定了不同土壤容重(1.25g/cm3、1.40g/cm3、1.55g/cm3)下平邑甜茶幼苗叶片的光合参数和水分利用效率.结果表明,3个处理植株叶片净光合速率、叶片组织含水量和组织相对含水量差异不显著,1.55g/cm3处理的气孔导度、胞间CO2浓度和叶片蒸腾速率显著低于1.25g/cm3和1.40g/cm3的处理,而植株叶片瞬时水分利用效率显著高于1.25g/cm3和1.40g/cm3的处理,但是1.25g/cm3和1.40g/cm3处理以上5项指标均差异不显著;叶绿素含量表现为1.40g/cm3>1.25g/cm3>1.55g/cm3.【总页数】2页(P137-138)【作者】吴亚维;马锋旺;邹养军【作者单位】西北农林科技大学,园艺学院,陕西,杨凌712100;贵州省果树科学研究所,贵州,罗甸,550100;西北农林科技大学,园艺学院,陕西,杨凌712100;西北农林科技大学,园艺学院,陕西,杨凌712100【正文语种】中文【中图分类】S151.9+2【相关文献】1.土壤紧实度对平邑甜茶幼苗生长及叶绿素荧光参数的影响 [J], 吴亚维;邹养军;马锋旺;李明军;高锡明;韩明玉;束怀瑞2.土壤紧实度对楸子幼苗根系生长及活力的影响 [J], 吴亚维;马锋旺;邹养军3.钛离子注入种子对油茶幼苗叶绿体DNA、叶绿素含量和光合参数的影响 [J], 李维伟;黎傲日;郭怡菲;李观林;肖文静;曾艳玲4.钛离子注入种子对油茶幼苗叶绿体DNA、叶绿素含量和光合参数的影响 [J], 李维伟;黎傲日;郭怡菲;李观林;肖文静;曾艳玲5.不同土壤紧实度对平邑甜茶根系特征及氮代谢的影响 [J], 生利霞;冯立国;束怀瑞因版权原因,仅展示原文概要,查看原文内容请购买。
有机物料对平邑甜茶实生苗光合特性的影响毛志泉;束怀瑞;张继祥;姬谦龙【期刊名称】《山东农业大学学报(自然科学版)》【年(卷),期】2002(033)002【摘要】试验结果表明,不同有机物料栽培的平邑甜茶(Malus hupenensis Rehd.)实生苗光合速率不同,鸡粪处理表现为植株生长前期叶片光合速率高于羊粪处理,生长后期则刚好相反,花生秧处理的光合速率整个生长季节均较低.夏季平邑甜茶光合作用有明显的午休现象,但不同处理引起午休的原因则不同,鸡粪处理主要为气孔限制,而羊粪和花生秧处理则为非气孔限制.在栽培介质脱水过程中,有机基质栽培的平邑甜茶叶片光合速率下降幅度远小于土壤栽培.【总页数】4页(P230-233)【作者】毛志泉;束怀瑞;张继祥;姬谦龙【作者单位】山东农业大学园艺学院,山东,泰安,271018;山东农业大学园艺学院,山东,泰安,271018;山东农业大学园艺学院,山东,泰安,271018;山东农业大学园艺学院,山东,泰安,271018【正文语种】中文【中图分类】S66【相关文献】1.施用有机物料和氮肥对平邑甜茶实生苗生物学效应的灰色系统评价 [J], 许衡;杨和生;束怀瑞2.有机物料对平邑甜茶根系32P吸收动力学参数的影响 [J], 毛志泉;张继祥;胡艳丽;张江红;王丽琴;束怀瑞3.有机物料发酵流体和堆肥对苹果连作土壤环境及平邑甜茶幼苗生物量的影响 [J], 张国栋;展星;李园园;沈向;陈学森;吴树敬;毛志泉4.有机物料对平邑甜茶实生苗根系呼吸强度的影响 [J], 毛志泉;王丽琴;沈向;束怀瑞;邹岩梅5.有机物料对连作平邑甜茶幼苗生长及微生态环境的影响 [J], 苏立涛;沈向;郝云红;毛志泉因版权原因,仅展示原文概要,查看原文内容请购买。
甜瓜叶片光合产物输出能力对弱光的响应杨柳燕;陈菁菁;陈年来【期刊名称】《中国农业科学》【年(卷),期】2018(051)013【摘要】[目的]研究甜瓜果实发育期坐果节位叶片光合能力、蔗糖合成能力和水苏糖装载能力对遮光的响应,分析不同耐弱光性甜瓜品种同化物输出能力的差异,为进一步研究甜瓜耐弱光品种果实糖分卸载与积累机理奠定基础.[方法]以耐弱光甜瓜品种‘玉金香’和不耐弱光品种‘钰雪三号’为试材,在日光温室条件下于授粉后进行遮光处理,每5d取样一次,取坐果节位叶片测定叶绿素含量、气体交换参数、葡萄糖、果糖、蔗糖、肌醇半乳糖苷、棉子糖、水苏糖和淀粉含量,同时测定叶片蔗糖磷酸合成酶(SPS)、蔗糖合成酶(SS)、中性转化酶(NI)、酸性转化酶(AI)、肌醇半乳糖苷合成酶(GS)和水苏糖合成酶(STS)活性.[结果]遮光处理后,两个甜瓜品种叶片叶绿素a/b、净光合速率(Pn)、葡萄糖、果糖和蔗糖含量均显著降低,淀粉含量升高,‘玉金香’叶绿素a/b降幅(1 0.0%)大于‘钰雪三号’(5.8%),而Pn、蔗糖含量降幅(分别为30.3%和30.9%)和淀粉含量增幅(3.6%)均显著小于‘钰雪三号’(分别为45.2%、60.6%和20.4%).遮阴条件下,两个品种叶片蔗糖代谢相关酶SPS、SS、AI和NI活性均显著降低,‘玉金香’SPS和SS酶活性降幅(分别为16.5%和30.0%)显著小于‘钰雪三号’(31.6%和40.5%),而AI和NI降幅(分别为23.8%和12.7%)高于‘钰雪三号’.遮光后甜瓜叶片肌醇半乳糖苷含量和GS活性均显著降低,但品种间差异不显著.棉子糖、水苏糖和STS活性均显著降低,‘玉金香’棉子糖含量降幅(65.3%)显著高于‘钰雪三号’(35.0%),而水苏糖和STS活性降幅(分别为79.5%和23.8%)小于‘钰雪三号,.[结论]遮光条件下,耐弱光品种‘玉金香’叶片蔗糖合成能力和水苏糖装载能力下降较少,具有较强的同化物输出能力.【总页数】9页(P2561-2569)【作者】杨柳燕;陈菁菁;陈年来【作者单位】甘肃农业大学园艺学院,兰州730070;甘肃农业大学园艺学院,兰州730070;甘肃农业大学园艺学院,兰州730070【正文语种】中文【相关文献】1.西葫芦幼苗叶片对低温弱光胁迫的响应 [J], 安福全;于龙凤;李富恒2.辣椒幼苗叶片光合特性对低温、弱光及盐胁迫3重逆境的响应 [J], 张志刚;尚庆茂3.弱光对黄瓜幼苗叶片光合产物积累的影响 [J], 李丹丹;张丹;张有利;张欣悦;宿畅;李季;张洋;林玉4.甜瓜叶片对弱光转强光的生理响应 [J], 吴燕;高青海5.低温弱光胁迫下甜瓜幼苗叶片叶绿素和根系丙二醛变化规律研究 [J], 宋慧;张凌霄;黄芸萍;臧全宇因版权原因,仅展示原文概要,查看原文内容请购买。
化学农药在茶树叶片上的光解速率预测
陶思明;陈宗懋
【期刊名称】《环境科学学报》
【年(卷),期】1994(14)1
【摘要】根据光化学基本理论,推导出描述均匀薄层固相农药光解速率的数学模型.依此模型,提出了固相农药在313nm波长处的光化学反应量于产率的测定方法.利用人工紫外光和自然光对5种农药在惰性玻璃表面和茶树叶表面的光解速率进行了研究,建立了茶树叶表上农药在自然日光下的光解速率和半衰期预测数学模型.实测表明,模型预测值与实测值吻合.
【总页数】8页(P64-71)
【关键词】茶树;农药;光解速率;残留;预测
【作者】陶思明;陈宗懋
【作者单位】中国农业科学院茶叶研究所,湖南省化工研究院
【正文语种】中文
【中图分类】X592.032.3
【相关文献】
1.茶树叶片净光合速率对生态因子的响应 [J], 林金科;赖明志;詹梓金
2.影响茶树叶片净光合速率的生态生理因子的初步分析 [J], 吴敏生;王守才;戴景瑞
3.小贯小绿叶蝉在5个茶树品种(系)上的蜜露排泄量与茶树叶片结构比较 [J], 刘丰静;冉伟;李喜旺;汪素琴;孙晓玲
4.茶树叶片净光合速率与儿茶素组分的相关性研究 [J], 涂良剑;林用松;杨志坚;黄则栋;赵文净;孙平;严婷婷;郑金贵
5.利用茶树叶片解剖结构指数预测茶树种质材料的抗寒性 [J], 王玉;洪永聪;丁兆堂;张新富;王漪;范凯
因版权原因,仅展示原文概要,查看原文内容请购买。
10种茶花叶片的光合和荧光特性刘落鱼;佘汉基;周彤彤;薛立【期刊名称】《经济林研究》【年(卷),期】2018(036)004【摘要】为给茶花的培育和筛选提供参考依据,以广东省佛山市林业科学研究所山茶园的10种茶花为材料,通过测定其光合特性参数和叶绿素荧光参数,揭示其光合荧光的基本生理特征.结果表明:10种茶花的主要光合生理特性因子中,净光合速率(Pn)、气孔导度(Gs)、胞间二氧化碳浓度(Ci)、蒸腾速率(Tr)、气孔限制值(Ls)、水分利用率(WUE)的范围值分别为3.77~11.66μmol·m-2s-1、0.04~0.17 mol·m-2s-1、236.37~380.79μmol·mol-1、0.71~3.14 mmol·m-2s-1、0.05~0.41、1.81~5.43μmol·mol-1;10种茶花的主要叶绿素荧光参数中,最大荧光(Fm)、PSII实际光量子效率(YI)、表观电子传递速率(ETR)、PSII非调节性能量耗散(YNO)、非光化学淬灭系数(NPQ)的范围分别为229.75~358.00、0.04~0.07、3.15~5.11、0.66~0.72、0.14~0.30.10种山茶中,茶梅和张氏红山茶均为高光合能力品种,适宜栽培在水肥条件较好的地方;烈香和甜香水均为低光合、低蒸腾和高水分利用效率的品种,适宜栽植于较干旱的环境中;另外,烈香、杰作和张氏红山茶均适宜于强光环境中生长,而甜香水、茶梅、海泡和赛桃红则与之相反.【总页数】5页(P145-149)【作者】刘落鱼;佘汉基;周彤彤;薛立【作者单位】华南农业大学林学与风景园林院,广东广州 510642;华南农业大学林学与风景园林院,广东广州 510642;华南农业大学林学与风景园林院,广东广州510642;华南农业大学林学与风景园林院,广东广州 510642【正文语种】中文【中图分类】S685.14【相关文献】1.光质对芹菜叶片光合色素和光合荧光特性的影响 [J], 尹娟2.玉米(Zea mays L.)杂交种(中单14)及其亲本部分光合特性的研究Ⅰ.杂交种和亲本自交系叶片荧光特性和光合强度比较 [J], 赵琦;唐崇钦;匡廷云3.施用生物炭和硅肥对增温水稻叶片光合及荧光特性的影响 [J], 邢钰媛;娄运生;王坤;刘健4.不同树形对灰枣叶片光合及叶绿素荧光特性的影响 [J], 王文军;陈奇凌;郑强卿;王晶晶;王振东5.不同耐/感玉米品种的叶片光合荧光特性、授粉结实和产量构成因素对花期高温的反应 [J], 穆心愿;齐建双;赵霞;唐保军;马智艳;张兰薰;付景;刘天学;丁勇;夏来坤;张凤启;张君因版权原因,仅展示原文概要,查看原文内容请购买。
中国农业科学 2010,43(15):3176-3183Scientia Agricultura Sinica doi: 10.3864/j.issn.0578-1752.2010.15.015平邑甜茶叶片光合速率及叶绿素荧光参数对氯化镉处理的响应王 利1,2,杨洪强1,3,范伟国3,张 召2(1山东农业大学资源与环境学院农业资源利用博士后流动站,山东泰安 271018;2山东农业大学林学院农业生态与环境重点实验室,山东泰安 271018;3山东农业大学园艺科学与工程学院/作物生物学国家重点实验室,山东泰安 271018)摘要:【目的】研究氯化镉处理对平邑甜茶叶片光系统Ⅱ(PSⅡ)活性、光合速率影响及其相互关系,为进一步揭示镉伤害机理提供理论依据。
【方法】平邑甜茶在含不同浓度氯化镉1/2 Hoagland营养液中培养30 d后,测定其叶片光合速率(Pn)、气孔导度、胞间CO2浓度和荧光参数等,分析氯化镉处理后这些参数间的关系。
【结果】在氯化镉处理下,平邑甜茶叶片光合速率和气孔导度显著降低,胞间CO2浓度增加,300 μs时的叶绿素荧光强度(Fk)提高,PSⅡ最大光化学效率(Fv/Fm,φPo)、用于电子传递的量子产额(φEo)、光化学性能指数(PI ABS)以及有活性的反应中心的密度(RC/CS)明显下降,并且这些参数的变化幅度随着氯化镉浓度的增加而提高;通径分析显示,300 μs时的相对可变荧光强度(V K)及其可变荧光Fv占(J相的荧光强度Fj-O相的荧光强度Fo)振幅的比例(W K)对Pn的直接作用高于其它荧光参数。
【结论】氯化镉使平邑甜茶叶片PSⅡ供体侧、受体侧和反应中心受到显著伤害,从而降低了PSⅡ活性和光合速率;在氯化镉处理下,V K和W K对Pn的直接作用比较大。
关键词:平邑甜茶;氯化镉;光合速率;光系统Ⅱ;叶绿素荧光Effect of CdCl2 Treatment on PhotosyntheticRate and Chlorophyll Fluorescence Parameters inMalus hupehensis LeavesWANG Li 1,2, YANG Hong-qiang 1,3, FAN Wei-guo3, ZHANG Zhao2(1Post-Doctoral Mobile Station of Agricultural Resource Utilization, College of Resources and Environment, Shandong AgriculturalUniversity, Taian 271018, Shandong; 2Key Laboratory of Agricultural Ecology and Environment, College of Forestry, ShandongAgricultural University, Taian 271018, Shandong; 3State Key Laboratory of Crop Biology/College of Horticultural Science andEngineering, Shandong Agricultural University, Taian 271018, Shandong)Abstract: 【Objective】For discovering the mechanism of Cd damage on leaves of Malus hupehensis Rehd., the activity of photosystemⅡ (PSⅡ), net photosynthetic rate (Pn) and their correlation in leaves treated with CdCl2 were studied. 【Method】 After30 days of treatment by CdCl2 in 1/2 Hoagland solution, the Pn, stomatal conductance (Gs), intercellular CO2 concentration (Ci) andchlorophyll fluorescence parameters in leaves of Malus hupehensis Rehd. were measured, and the relationship between theseparameters under CdCl2 treatment were analyzed. 【Result】Under the treatment of CdCl2, the Pn and Gs reduced, the Ci and the fluorescence intensity Fk at 300 μs increased, and the maximum photochemistry efficiency of PSⅡ(Fv/Fm, φPo), the quantum yieldfor electron transport (φEo) , the performance index on absorption basis (PI ABS) and the density of active reaction center (RC/CS) alldecreased significantly. Furthermore, the range of variation of these parameters increased with the increasing of CdCl2 concentration.The direct effect of the relatively variable fluorescence intensity V K and the ratio of variable fluorescence Fv on the amplitude Fj-Fo(W K) at 300 μs for Pn were higher than that of others through the path analysis. 【Conclusion】 CdCl2 damaged the sides of acceptorand donor and the reaction centers of PSⅡ of leaves of Malus hupehensis Rehd. The activity of PSⅡand Pn decreased, and the direct收稿日期:2009-12-02;接受日期:2010-03-01基金项目:山东农业大学博士后项目、国家自然科学基金项目(30671452)作者简介:王利,副教授,博士。
E-mail:liwang6868@。
通信作者杨洪强,教授。
E-mail:hqyang@15期王利等:平邑甜茶叶片光合速率及叶绿素荧光参数对氯化镉处理的响应 3177effects of V K and W K on the important fluorescence parameters for Pn were larger than other parameters under CdCl2 treatment.Key words: Malus hupehensis Rehd.; CdCl2; photosynthetic rate; photosystemⅡ; chlorophyll fluorescence0 引言【研究意义】农药化肥的不合理使用,以及工业“三废”和生活垃圾的不合理排放,使农业生产环境和农产品质量中的重金属等有害物质呈现增加的趋势,这对人类的生产和生活构成了严重威胁[1]。
镉是毒性很强的重金属,它不能被微生物所降解,进入土壤后会长期存在,但在过去50年中,全球排放到环境中的镉(Cd)达到22 000 t[2]。
重金属会破坏植物光合系统,其中光系统Ⅱ(PSⅡ)是对环境变化比较敏感的部位,PSⅡ活性降低会导致光合速率下降[3]。
平邑甜茶属于湖北海棠(Malus hupehensis Rehd.)的一个变种,是中国特有的木本植物资源,常用作苹果砧木和果树根系研究[16];此外,平邑甜茶也是一种类茶植物,可加工成饮料供人们饮用 [17],但不论用作苹果砧木还是用来制茶,环境中的镉对平邑甜茶和人类健康都存在严重威胁。
叶绿素荧光参数能够反映叶片PSⅡ活性,研究氯化镉处理下叶片光合速率和叶绿素荧光参数等的变化,对于揭示植物镉伤害的机制以及探讨缓解镉的措施具有重要意义。
【前人研究进展】土壤镉污染会直接损伤植物根系,抑制根系质子泵的活性和营养元素的吸收及转运[4],同时也严重干扰叶片的光合作用,改变细胞质膜脂肪酸构成[5-6],并通过活性氧引起膜脂过氧化,破坏细胞质膜的完整性,严重时会导致细胞和植株死亡[7-8]。
光合作用是植物将光能转化为化学能的最重要过程,PSⅡ反应中心的功能是限制光能利用和光合作用正常运行的关键环节,而叶片叶绿素荧光诱导动力学可以在非离体条件下方便快速反映PSⅡ反应中心、电子供体侧和受体侧的功能变化[9-11]。
人们根据荧光动力学曲线的特征变化,发现逆温、强光、盐及干旱等严重影响多种植物PSⅡ的活性[12-14],还发现铅、镉等重金属能够降低玉米、杨树叶片PSⅡ的活性,破坏光系统的功能[3,15]。
【本研究切入点】重金属如何以及在PSⅡ的哪个位点损伤光系统目前还不明确,重金属处理下光合速率与荧光参数之间的相关关系也未见报道。
【拟解决的关键问题】本文以平邑甜茶为材料,通过测定氯化镉处理后叶片叶绿素荧光诱导动力学曲线及光合速率等参数,分析氯化镉处理与PSⅡ活性和光合速率的关系,为进一步揭示镉伤害机理奠定基础。
1 材料与方法1.1 材料与处理取大小一致的平邑甜茶种子,经消毒层积1个多月后播在育苗容器中。
当幼苗第2片真叶刚出现时,选生长相近的植株移至1/2 Hoagland营养液中,每3 d 换1次培养液,培养至出现第6片叶后,在营养液中加入氯化镉,使其终浓度为0(CK)、0.5 mg·L-1、5 mg·L-1、10 mg·L-1,每处理4株,重复3次,30 d后,检测叶片的光合速率和叶绿素荧光参数。
1.2 光合速率和气孔导度的测定利用CIRAS-2型便携式光合作用测定系统(PP-Systems,英国)在室内人工控制光强 1 000 μmol·m-2·s-1的条件下,对利用氯化镉处理30 d后的平邑甜茶幼苗测定其净光合速率(Pn)、气孔导度(Gs)和胞间CO2浓度(Ci),分别测定各处理每单株第4片叶片的Pn 、Gs 、Ci求其平均值。