λ噬菌体载体的类型
- 格式:ppt
- 大小:1.49 MB
- 文档页数:96

λ噬菌体载体是一种广泛应用于分子生物学和基因工程领域的载体,用于将外源DNA序列引入细菌细胞中。
噬菌体是一种寄生性病毒,可以感染细菌并在其内部复制。
而λ噬菌体是其中最为常见和常用的一种。
λ噬菌体载体通常由数万个碱基对的环状DNA组成,其中包含了多个重要的功能区域。
其中,最重要的功能区域是Origins of Replication(ORI),即复制起始点,负责引导DNA的复制。
此外,载体还包含了选择性标记基因,如抗生素抗性基因,以便在细菌培养基中筛选带有该载体的细菌。
λ噬菌体载体还含有多个限制内切酶切位点,这些切位点可以用于将外源DNA序列插入到载体的特定位置上。
通过将外源DNA与载体进行限制性内切酶切割,然后使用DNA连接酶进行连接,可以将外源DNA序列插入到载体的DNA链上。
这一过程称为重组。
一旦重组完成,λ噬菌体载体可以通过转化的方式引入到宿主细菌中。
转化是指将外源DNA 导入到细菌细胞中的过程。
一旦载体进入到细菌细胞中,它会在细菌细胞内部复制,并产生大量的噬菌体颗粒。
这些噬菌体颗粒可以感染其他细菌细胞,并将携带的外源DNA序列传递给它们。
λ噬菌体载体在分子生物学和基因工程研究中具有广泛的应用。
它可以用于构建基因文库,即将外源DNA序列插入到载体上,并通过转化的方式导入到细菌细胞中。
这样,研究人员就可以通过筛选和分析细菌细胞中的载体来获得感兴趣的外源DNA序列。
此外,λ噬菌体载体还可以用于基因表达,即将外源DNA序列插入到载体的表达位点上,以便在细菌细胞中大量产生特定的蛋白质。
总之,λ噬菌体载体是一种在分子生物学和基因工程领域中被广泛使用的载体。
它具有多个重要的功能区域,可以用于将外源DNA序列引入到细菌细胞中,并在其中进行复制和表达。
通过利用λ噬菌体载体,研究人员可以进行基因库构建、基因表达和其他相关研究,为生物技术的发展提供了重要的工具和平台。

3、λ噬菌体载体的优缺点:•优点:包装的λ噬菌体感染大肠杆菌要比质粒转化细菌的效率高。
•缺点:λ噬菌体载体的克隆操作要比质粒载体复杂。
•用途:λ噬菌体载体比质粒载体能插入的DNA长得多,常用于构建cDNA文库或基因组文库。
第一章分子克隆的工具酶和载体•第八节噬菌体载体•一、λ噬菌体•(一)λ噬菌体•(二)λ噬菌体载体的改造•(三)λ噬菌体载体举例(三)λ噬菌体载体举例•Lambda gt10•Lambda gt11•EMBL3和EMBL4Lambda gt10概述:•Lambda gt10是一种插入载体。
•在噬菌体阻遏基因cI内有单一的EcoRⅠ克隆位点。
用于插入小的cDNA片段(约6kb),构建cDNA文库或基因文库。
•该载体克隆效率很高。
•在构建cDNA文库时,利用Oligo(dT)或随机引物合成的cDNA经过EcoRⅠadaptors或Linkers修饰后,就可以和λgt10连接起来。
•克隆到λgt10的噬菌体,可用核酸探针进行筛选。
Lambda gt10map宿主:•建议用C600 and C600hf1作受体菌。
筛选:•如果有外源DNA插入,cI基因失活,该噬菌体进入裂解生长途径,在培养皿形成噬菌斑。
反之,若无插入,cI基因表达,噬菌体进入溶原生长途径,不形成噬菌斑。
•核酸探针杂交。
Insertional cloning•Insertional cloning into the cI gene of thelambda -gt10 cDNA cloning vector (DNA inserts of ~1-5 kb) can be selected in hfl (highfrequency of lysogeny ) mutant strains of E. coli. In hflA strains of E. coli, expression of the lambda cII gene is elevated, resulting in transcriptional induction of the lambda cI repressor gene which promotes lysogeny . Disruption of the lambda cI codingsequence by DNA insertion into the unique EcoRI site of the lambda gt10 cDNA cloningvector, blocks the lysogenic pathway leading to cell lysis and plaque formation.Lambda gt11•λgt 载体系列:是插入型载体。


基因工程的载体载体的特征:在寄主细胞中能够自主复制;有一种或多种限制酶的单一切割位点,并在此位点插入外源基因片段;在基因组中有遗传标记,为寄主细胞提供易于检测的表型特征;载体分子较小,以便体外基因操作;对于表达型载体还应具有与宿主细胞相适应的启动子、增强子、加尾信号等基因表达元件;载体的类型⏹质粒⏹噬菌体⏹其他载体(如:酵母人工染色体、细菌人工染色体、植物Ti质粒、动物病毒)质粒(plasmid): 多数情况下,质粒是存在于细菌染色体外的小的双链闭合环状DNA分子,能自主复制,并在细胞分裂时遗传给子代细胞⏹并不是所有的质粒都是环状分子, 在多种细菌中都发现有线性质粒⏹质粒广泛存在于原核生物中, 其大小从相对分子量小于1X106 到大于200X106质粒的形态1) cccDNA—双链闭合环状DNA2) ocDNA—开环DNA3) cDNA—线形DNA(L型)在DNA促旋酶(gyrase)作用下成负超螺旋构型, 在拓扑异构酶I的作用下解旋。
溴化乙锭(ethidium bromide,EtBr)也有解旋作用溴化乙锭插入DNA超螺旋的作用随着插入的溴化乙锭数目增加,双螺旋解旋,导致超螺旋减少直至产生环状分子的开放形式. 进一步的插入在双螺旋中引入了过多的螺旋,导致反义的超螺旋(注意B和D处的螺旋方向). 为了清楚起见,只表示了双螺旋的一条单链质粒DNA的复制类型⏹严紧型质粒这些质粒的复制是在寄主细胞严格控制之下的,与寄主细胞的复制偶联同步。
所以,往往在一个细胞中只有一份或几份拷贝⏹松弛型质粒这些质粒的复制是在寄主细胞的松弛控制之下的,每个细胞中含有10-200份拷贝,如果用一定的药物处理抑制寄主蛋白质的合成才会使质粒拷贝数增至几千份。
如较早的质粒pBR322即属于严紧型质粒,要经过氯霉素处理才能达到更高拷贝数穿梭载体(shuttle vector) 可以在两种生物体内复制的载体分子质粒的命名规则⏹小写字母p表示质粒(plasmid)⏹p后面的两个大写字母表示发现或者构建该质粒的作者或者实验室名称⏹数字表示编号⏹例:pBR322、pET21、pGEM-T质粒的宿主范围⏹质粒只编码少数几个其自身复制所需要的蛋白质,甚至在许多情况下只编码其中一个蛋白质⏹所有其他复制所需的蛋白,包括DNA聚合酶、DNA连接酶、解旋酶等都是由宿主细胞提供的⏹质粒所编码的复制蛋白质定位在ori 附近,因此只有ori 周围的一小部分区域是复制所必需的⏹因此,把质粒的其他部分删除掉,把外源序列加到质粒上,复制仍然可以继续进行⏹质粒的宿主范围是由它的ori 决定的质粒的不相容性⏹在没有选择压力的情况下,两种质粒不能共存于同一个宿主细胞内⏹如果质粒拥有相同的复制调控机制,它们就不相容显性质粒和隐蔽质粒⏹显性质粒(表达型质粒)----除了携带与本身复制和转移有关的基因外, 还携带一些其他的基因, 宿主细胞由于含有这样的质粒而呈现出新的性状, 这样的质粒称为显性质粒⏹隐蔽质粒----无异常性状表现出来克隆载体例:pBR322、pUC18、pUC19、pGEM-T表达载体例:pET-21、pGEX⏹质粒DNA的制备碱裂解法、煮沸法、层析柱过滤法碱裂解法原理:在高pH的碱性条件下,染色体DNA和蛋白质变性,质粒DNA由于其超螺旋共价闭合环状结构,尽管其DNA的大部分氢键也断裂,但是双链DNA仍然不会分离,当恢复到中性时,染色体DNA复性,并聚集形成不可溶的网架。



λDNAλDNA,就是λ噬菌体中的DNA,但是λDNA也分很多种情况的,有正常的,有突变的,还有整合了宿主染色体的。
λDNA是一种溶原性的染色体序列,可以整合到宿主的染色体组上,也可以脱离下来,他的整合和脱离所产生的失误可产生宿主的基因重组现象,所以可以用于局限转导,是一种基因转化的载体。
柯斯质粒载体目录一.柯斯质粒载体的来源二.柯斯质粒载体的特点三.柯斯克隆四.柯斯克隆的优点一.柯斯质粒载体的来源1978年由collins和hohn改建的一种新型大肠杆菌克隆载体,用正常的质粒与噬菌体λ的cos位点构成。
“cosmld”一词是由英文“cos site-carrying plasmid”缩写而成的,其原意是指带有粘性末端位点(cos)的质粒。
所谓柯斯质粒,乃是一类由人工构建的含有λ DNA的cos序列和质粒复制子的特殊类型的质粒载体。
诸如右图所示的柯斯质粒载体pHC79,就是由λ DNΑ片段和pBR322质粒DNA联合组成的。
一般长度4~6kb。
含有Amp和Tet选择标记基因。
其上的cos位点可识别噬菌体外壳蛋白。
凡具有cos位点的任何DNA分子只要在长度上相当于噬菌体的基因组,就可以同外壳蛋白结合而被包装成类似噬菌体λ的颗粒。
因此,插入柯斯质粒载体的外源DNA片段的长度可大于40kb,从而大大增加了载体的携带能力。
二.柯斯质粒载体的特点柯斯载体的特点大体上可归纳成如下四个方面:第一,具有λ噬菌体的特性。
柯斯质粒载体在克隆了合适长度的外源DNA,并在体外被包装成噬菌体颗粒之后,可以高效地转导对λ噬菌体敏感的大肠杆菌寄主细胞。
第二,具有质粒载体的持性。
柯斯质粒载体具有质粒复制子,因此在寄主细胞内能够像质粒DNA一样进行复制,并且在氯霉素作用下,同样也会获得进一步的扩增。
此外,柯斯质粒载体通常也都具有抗菌素抗性基因,可供作重组体分子表型选择标记。
第三,具有高容量的克隆能力。
柯斯质粒载体的分子仅具有一个复制起点,一两个选择记号和COS位点等三个组成部分,其分子量较小,一般只有5~7kb左右。
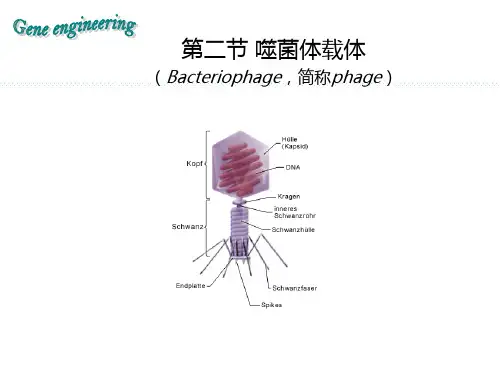
λ噬菌体载体λ噬菌体,一种大肠杆菌双链RNA噬菌体。
是一种中等大小的温和噬菌体。
迄今已经定位的λ噬菌体的基因至少有61个,其中有一半左右参与了噬菌体生命周期的活动,我们称这类基因为λ噬菌体的必要基因;另一部分基因,当它们被外源基因取代之后,并不影响噬菌体的生命功能,我们称这类基因为λ噬菌体的非必要的基因。
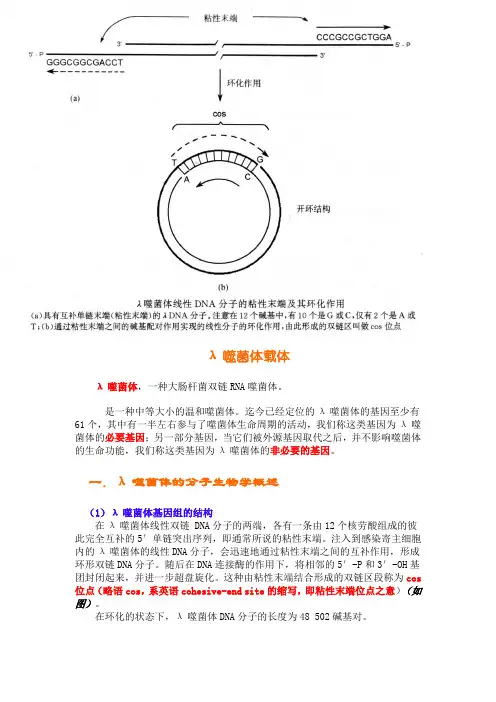
一.λ噬菌体的分子生物学概述(1)λ噬菌体基因组的结构在λ噬菌体线性双链 DNA分子的两端,各有一条由12个核劳酸组成的彼此完全互补的5′单链突出序列,即通常所说的粘性末端。
注入到感染寄主细胞内的λ噬菌体的线性DNA分子,会迅速地通过粘性末端之间的互补作用,形成环形双链DNA分子。
随后在DNA连接酶的作用下,将相邻的5′-P和3′-OH基团封闭起来,并进一步超盘旋化。
这种由粘性末端结合形成的双链区段称为cos 位点(略语cos,系英语cohesive-end site的缩写,即粘性末端位点之意)(如图)。
在环化的状态下,λ噬菌体DNA分子的长度为48 502碱基对。
(2)λ噬菌体DNA的复制在λ噬菌体感染的早期,环形的λ DNA分子按θ形式从双向进行复制。
到了感染的晚期,控制滚环复制机理的开关被启动了,合成出了由一系列线性排列的人基因组oxA组成的长多连体分子。
(3)λ噬菌体DNA的整合与删除λ噬菌体基因组的整合作用,是通过它的附着位点att,同大肠杆菌染色体DNA的局部同源位点之间的重组反应实现的。
整合作用需要int基因的表达,它是一种可逆的过程。
的复制子,这种过程叫做原噬菌体的删除作用。
λ噬菌体的删除,需要噬菌体xis基因和bio基因的协同作用才能实现。
(4)λ噬菌体DNA的转录与转译在溶菌周期,λ噬菌体DN A的转录是在三个时期,即早期、中期和晚期发生的。
大体的情况是,早期基因转录确立起溶菌周期;中期基因转录的结果导致DNA进行复制和重组;晚期基因的转录最终使DNA被包装为成熟的噬菌体颗粒。

生命科学学院病毒生物学λ噬菌体综述摘要:噬菌体是一类温和噬菌体,它们感染大肠杆菌后能进行溶菌性生长(Lytic growth)和溶源性生长(Lysogenic growth)。
其溶源特性对基因重组与遗传工程研究有很大帮助。
本文就λ噬菌体的基因组结构、溶关键词:λ噬菌体溶原性溶菌性基因克隆引言:大肠杆菌噬菌体λ为长尾噬菌体科,是一类中等大小的大肠杆菌病毒,其基因组为双链线状DNA,由48502对碱基组成,分子量3.2×107 ,约50个基因,特点是相关基因成簇排列,形成若干个操纵子。
基因组两端为粘性末端,中间有相当长的DNA片段是裂解生长非必需的,这就为其作为外源基因的克隆载体提供了方便。
λ噬菌体由头和尾构成,其基因组组装在头部蛋白质外壳内部,其序列已被全部测出。
感染时吸附位点为细胞壁。
属温和性感染;感染的DNA环化并整合于宿主基因组中。
以θ环双向复制,然后通过滚环机制单向复制。
用于感染大肠杆菌的λ噬菌体改造成的载体应用最为广泛。
1、The discovery of bacteriophage lambda1951年J. Lederberg的妻子Esther Lederberg第一个证明了 J. Lederberg和Tatum用来杂交的K-12中有原噬菌体,并命名为λ,经10年的研究搞清了溶原化的实质。
从此之后,λ噬菌体被广泛用于模式物种;1962年Esther Lederberg的同事并且还是她最好的朋友Allan Campbell首次发现了λDNA整合到细菌DN A的机制;之后由λ噬菌体改造后的载体广泛的用于基因工程。
2、The characteristics of bacteriophage lambda2.1、结构特点:λ为大肠杆菌温和性噬菌体,属长尾噬菌体科,头壳为直径约50nm的二十面体,其内包裹一长线状双链DNA分子(46500bp),因分子两端各有一含12个核苷酸的黏性末端,故又可黏合成环状分子。

第三章分子克隆载体(Molecular cloning vectors)一、名词解析二、填空题1.基因工程中有三种主要类型的载体、和。
2.就克隆一个基因来说,最简单的质粒载体也必须包括三个部分和、。
另外,一个理想的质粒载体必须具有低分子质量。
3.如果两个质粒不能稳定的存在已同一个宿主细胞中,则属于群,这是因为他们的所致。
4.pBR 322是一种改造型质粒,它的复制子来源于,它的四环素抗性基因来源于,它的氨苄青霉素抗性基因来源于。
5.Puc18质粒是目前使用较为广泛的载体。
pUC系列的载体是通过和两种质粒改造而来。
它的复制子来源于,Amp抗性基因则是来源于。
6.当λ噬菌体DNA进入宿主细胞以后是靠宿主细胞的和形成封闭的环状的DNA分子的。
7.λ噬菌体是感染大肠杆菌的噬菌体。
8.α-互补是指 lacZ基因上缺失近操纵基因区段的突变体与带有完整的近操纵基因区段的阴性的突变体之间实现互补。
9.溶源化的频率和与有关。
10.噬菌体DNA通过其上唯一的整合位点与宿主染色体DNA上的唯一整合位点发生重组,从而整合到染色体中。
11.通过分析大量缺陷型λ噬菌体的 DNA ,发现 J 基因与 Cro 基因之间的 DNA 被或替换后,不影响λ噬菌体裂解生长。
12.对野生型大肠杆菌来说,向溶源和裂解方向的转变是由和决定的。
13.代表性λ噬菌体载体有和14.粘粒的组成包括________,________,________。
15.柯斯质粒(Cosmid)载体:一种由________和_______cos尾巴构建的复合载体。
16.pcos1 EMBL 是设计用于筛选体内重组的重组粘粒文库,通过同源重组过程达到筛选目标克隆的目的。
该载体的复制起点来源于__________质粒,含_________和_________抗性基因。
17.酵母人工染色体由酵母染色体的__________、__________和__________等功能性DNA序列组成。