鱼类的性别决定与人工控制
- 格式:pdf
- 大小:186.95 KB
- 文档页数:4

水产养殖中的鱼类性别控制技术水产养殖一直是人类对海洋资源的有效利用方式之一,而鱼类性别控制技术在水产养殖中起着重要的作用。
通过控制鱼类的性别,可以更好地满足市场需求,提高养殖效益。
本文将探讨水产养殖中的鱼类性别控制技术及其应用。
一、鱼类分性别的意义鱼类的性别决定了其生长速度、体型特征以及繁殖能力等方面的差异。
在养殖业中,雄性鱼往往生长速度较快,体型较大,而雌性鱼则具有优良的繁殖能力。
通过科学地控制鱼类的性别,可以实现性别比例的均衡,提高养殖效益。
二、鱼类性别控制方法1. 温度控制法温度对鱼类的性别发育有着重要影响。
鱼类性别和温度之间存在一定的关联性,通过控制孵化阶段的水温,可以实现鱼类性别的选择。
一般而言,较高的水温有利于雌性发育,而较低的水温则有利于雄性发育。
2. 药物激素法药物激素法是一种常用的鱼类性别控制方法。
通过给鱼类注射或摄入一定剂量的雌激素或雄激素,可以影响其性腺的发育,从而控制其性别分化。
这种方法操作简单,成本低廉,但需要专业技术的支持。
3. 基因编辑法随着基因编辑技术的发展,越来越多的研究表明,通过基因的操控可以实现鱼类性别的控制。
通过修改或引入特定的基因,可以实现鱼类性别的选择。
这种方法具有较高的效率和精确性,但需要更高的技术要求和较大的投入。
三、鱼类性别控制技术的应用1. 提高养殖效益通过鱼类性别控制技术,可以实现性别比例的调控,从而更好地适应市场需求。
例如,在对高价值的雄性鱼进行特定养殖时,可以通过性别控制技术提高雄性比例,提高养殖效益。
2. 保护遗传资源对于濒危鱼类或具有重要遗传资源价值的鱼种,性别控制技术可以帮助实现种群的保护和遗传资源的合理利用。
通过性别控制,可以控制繁殖中的基因流失,保持遗传多样性。
3. 防止环境污染在水产养殖中,饲料的过剩和排泄物的积累是环境污染的主要原因之一。
通过鱼类性别控制技术,可以控制养殖密度和性别比例,减少饲料的浪费和污染物的排放,降低养殖对生态环境的影响。

鱼类的性别决定性别是生物界中最基本的属性之一,而对于鱼类来说,性别决定是一个非常关键的生物学问题。
鱼类的性别决定机制多样且复杂,涉及到遗传、环境和生殖生物学等多个方面。
本文将深入探讨鱼类性别决定的各种机制,并对其影响因素和意义进行分析。
1. 遗传性别决定在大部分鱼类中,性别是由遗传因素决定的。
这种遗传性别决定主要通过性染色体携带的基因来实现。
在一些物种中,雄性和雌性鱼类的染色体组成不同,如在某些鱼类中,XX染色体的个体为雌性,而XY染色体的个体为雄性。
这类似于人类的性染色体决定方式,但也存在一些例外情况。
例如,有些鱼类中,XY的个体变成雌性,而XX的个体变成雄性。
除了性染色体外,有些鱼类的性别决定也与其他染色体或基因相关。
例如,一些物种中,存在着多种染色体型,不同染色体型之间的个体性别表现出差异。
此外,还有一些物种中,个体的性别决定与其基因型有关。
这种情况下,同一性染色体组合的个体可能表现出不同的性别。
2. 环境因素的影响除了遗传因素外,环境因素也对鱼类性别决定起着重要作用。
某些鱼类的性别决定可以受到温度、光照周期等环境条件的影响。
例如,在一些热带鱼类中,温度对性别决定起着重要作用。
高温会导致个体发展为雌性,而低温则会促使个体发展为雄性。
类似地,光照周期也可以影响某些鱼类的性别决定。
这些环境因素通过影响个体的激素水平和发育过程来实现性别决定。
在一些鱼类中,环境因素与遗传因素相互作用,共同决定个体的性别。
例如,在七星刀鱼中,雌性个体在高温下产卵,而低温则会促使雄性个体发育。
这种性别决定形式的存在使得鱼类的性别在一定程度上具有适应环境变化的能力。
3. 性别决定的生殖生物学意义鱼类的性别决定对于其繁殖和适应生存环境至关重要。
不同性别的鱼类在不同的繁殖策略和生殖行为上存在差异。
例如,某些鱼类中的雄性个体会进行颜色变化或其他形式的繁殖行为来吸引雌性个体。
此外,一些鱼类还表现出性别的变态现象,即个体在一定阶段内具有男性特征,而在另一阶段则具有女性特征。

水产动物遗传学罗非鱼性别控制研究及应用水产动物遗传学是研究水产动物遗传与遗传变异的科学,可以为水产养殖提供重要的遗传改良手段。
在水产动物中,罗非鱼是一种重要的养殖鱼类,在全球范围内广泛分布。
了解罗非鱼的性别控制机制对于进行性别控制和性别选择具有重要的意义。
罗非鱼的性别决定机制是XX/XY,即雌性为XX,雄性为XY。
在罗非鱼研究中,存在着两种不同的性别控制模式:传统绝对性别控制和环境敏感性别控制。
传统绝对性别控制是指罗非鱼的性别完全由基因决定,与环境无关。
研究表明,Y染色体上的一个性别决定基因SDY是罗非鱼性别的主要决定因素。
它编码了一种性别决定蛋白,通过调节其他基因的表达来决定鱼的性别。
这种基于基因的性别决定机制具有稳定性高、遗传性强的特点,使得养殖者可以通过基因筛选和基因转导等手段,进行性别选择和性别控制,从而提高养殖效益。
环境敏感性别控制是指罗非鱼的性别决定不仅受基因的影响,还受环境条件的影响。
环境敏感性别控制的主要原因是温度。
研究表明,温度可以通过影响SDY基因的表达来控制罗非鱼的性别分化。
高温条件下,SDY基因表达量降低,导致雌性分化;低温条件下,SDY基因表达量增加,导致雄性分化。
因此,通过调节养殖水体温度,可以实现对罗非鱼性别的控制。
环境敏感性别控制具有灵活性强、操作简单的特点,对于养殖业来说具有重要的应用价值。
首先,利用基因筛选技术,选择具有良好性状的个体作为繁殖材料,加快罗非鱼的遗传改良进程。
通过筛选和配对,可以选择生长快、抗病能力强、食性适应广等优良性状的个体进行繁殖,提高后代的遗传水平。
其次,利用基因转导技术,实现罗非鱼性别控制。
通过将性别决定基因SDY导入罗非鱼胚胎,可以实现性别的选择和控制。
这种基因转导技术具有很大的潜力,在种质改良和繁殖中具有广阔的应用前景。
再次,利用温度调控技术,实现环境敏感性别控制。
通过调节水体温度,可以控制罗非鱼雌性和雄性的比例,在达到一定的温度条件下,实现性别的选择和控制。

鱼类分子生物学中的性别决定机制鱼类是一种非常特殊的生物,在其生命早期就需要决定其性别。
与哺乳动物和爬行动物不同,鱼类的性别决定机制更加灵活,可能受到环境和遗传因素的共同作用。
本文将详细介绍鱼类分子生物学中的性别决定机制。
一、鱼类性别决定基因的发现首先,我们需要知道鱼类的性别是由哪些基因决定的。
20世纪70年代以前,人们对鱼类性别决定机制的理解非常有限。
直到1972年,日本科学家Yasuo Nagahama和他的团队才首次发现了鲤鱼的性别决定基因。
这个基因被命名为sex-determining region Y(sry),是一个决定雄性性别的关键基因。
从此以后,人们开始运用基因工程和分子生物学技术在不同种类的鱼类中探索其性别决定机制。
通过对不同种类鱼类基因组的比较分析,人们发现鱼类性别决定基因形式多样,包括性染色体、单倍体基因、多倍体基因等。
二、鱼类性别决定基因的形式1. 性染色体性别决定许多鱼类的性别决定与哺乳动物和爬行动物类似,是由XY或ZW性染色体控制的。
在这种情况下,X或Z染色体是性别决定基因,从而决定了个体的性别。
例如,牛鱼的性别决定与人类的性别决定非常类似,都是由XY性染色体控制。
雌鱼有两个X染色体,而雄鱼则有一个X和一个Y染色体。
2. 单倍体基因性别决定在一些鱼类中,性别决定基因是由单个基因控制的,这类基因被称为性候选基因。
据统计,大多数这种鱼类的性别决定都与单倍体基因有关。
例如,日本鳞甲鲤就是一种由单倍体基因决定性别的鱼类。
日本鳞甲鲤的性别决定基因被命名为dmrt1,它能够控制个体的性别,并且还能控制生殖细胞的形成和发育。
3. 多倍体性别决定在鲈鱼等一些鱼类中,其性别决定机制被认为与多倍体基因有关。
这种性别决定形式在鱼类中比较罕见,但是它具有一定的普适性,能够解释鱼类性别决定中的一些奇异现象。
例如,鲈鱼的性别决定是由多倍体基因 cyp19a1b 控制的。
“cyp19a1b”基因编码酵素 aromatase,能够将雄性鱼体内的雄激素转化为雌激素。


鱼类的性别转换和性别控制1. 鱼类的性别大多数硬骨鱼类,一生或者只具有精巢,或者只具有卵巢(雌雄异体)。
但对于某些鱼类来说,体内同时存在卵巢和精巢(雌雄同体)则是一种正常生理现象,而且有的种类还能自体受精。
目前发现的雌雄同体鱼类约有400种,根据其生活史中卵巢和精巢在不同年龄阶段的发育进展情况,大致可分为3种类型:①雄性先成熟雌雄同体(protandrous hermaproditism)在生活史中由雄性转为雌性。
在性腺的发育过程中,早期卵巢的发育受到抑制,而精巢发育较快,低龄鱼表现为雄性,只能排精,不能产卵。
随着年龄增大,精巢逐渐萎缩,卵巢逐渐发育成熟,表现为雌鱼。
鲷科(Sparidae)鱼类中的黑鲷(Sparus macrocephalus)、黄鳍鲷(Sparus latus)、金头鲷(Sparus auratus)等属于这一类型。
②雌性先成熟雌雄同体(Protogynous hermaphroditism)与第一种相反,生活史中由雌性转为雄性。
低龄鱼卵巢先成熟,表现为雌性。
随着年龄的增大,卵巢萎缩吸收,精巢发育成熟。
在海水鱼类中有石斑鱼类中的Epinephelus aeneus、巨石斑鱼(Epinephalus tauvina)、灰石斑鱼(Epinephalus guttatus)等;淡水鱼类中有黄鳝(Monopterus albus)等。
这些鱼类第一次性成熟时都是雌鱼,产过卵以后才逐渐变为雄鱼。
有些自然性转换的鱼类,并不同时具有雌雄两性生殖腺,隆头鱼科中的盔鱼(Coris julis)是先表现为雌性功能,然后才转换为雄性功能的雄鱼,但没有观察到它同时有卵巢和精巢。
盔鱼的性转换特点是雌性生殖细胞完全为雄性生殖细胞所代替。
在性转换开始时,先是卵母细胞的萎缩,然后才出现精原细胞。
精原细胞是由分布在卵巢壁上的原生殖细胞分化出来的。
盔鱼的性细胞转换是在卵巢内部发生。
自然性逆转早期阶段的赤点石斑鱼性腺组织学切片埋植AI后赤点石斑鱼性腺组织结构的变化鱼类同时具有雌雄性腺,但并不同时成熟,不同的年龄表现为不同的性别,即在生活史中性别有一个转换的过程,这种现象我们称为性转换,也有人称为“性逆转”、“性位移”或“性邻接”。

控制鱼类性别的方法
摘要:控制鱼类性别是提高鱼产量一个有效方法。
一般利用性激素就能诱导鱼的性别向人们希望的方向发展。
性激素分两种,睾丸分泌的为雄性激素,卵巢分泌的为雌性激素。
鱼类的原始生殖上皮组织,同时具有决定向雌性发育和向雄性发育的髓质部的两个组成部分。
在雌性激素作用下皮质控制鱼类性别是提高鱼产量一个有效方法。
一般利用性激素就能诱导鱼的性别向人们希望的方向发展。
性激素分两种,睾丸分泌的为雄性激素,卵巢分泌的为雌性激素。
鱼类的原始生殖上皮组织,同时具有决定向雌性发育和向雄性发育的髓质部的两个组成部分。
在雌性激素作用下皮质部分化为卵巢,髓质部不发育而呈雌性具有产卵生殖功能;反之,在雄性激素作用下,鱼会变成雄性。
因此,在发展渔业生产过程中,若想增加雌性鲤鱼和鲫鱼等鱼类的数量,可采用雌性激素进行处理,即在鱼苗池中子鱼开始摄食时,将饲料中添入一定量的苯甲酸二醇,连续处理2个~3个月,雌性比例就可以达到95%左右。
同样,用雄性激素甲基睾丸酮处理罗非鱼、黄鳝等鱼苗,可获得99%以上的雄性鱼。
经过处理后的鱼类因性别单一,密度固定,不仅生长快,而且成本低,一般可
增产30%左右。


浅谈鱼类的性别与性别控制摘要鱼类的性别控制对于提高鱼类养殖效益具有明显的应用价值, 目前已经成为水产养殖业的研究热点之一。
为此本文综述了鱼类性别的研究现状和性别控制的主要方法,并展望其发展前景。
关键词鱼类;性别;性别控制除病毒外, 几乎所有生物都有性别。
性别这一生命现象经历了漫长的进化过程, 它与有性生殖同时出现[]1。
鱼类性别控制对于提高鱼类的养殖效益具有十分明显的应用价值。
因为许多鱼类雌雄个体之间的经济性状存在着明显的差异(如生长率、个体大小等), 大多数鱼类性成熟后生长速度会减慢, 自然生殖活动还带来生长的停滞, 体组织可食部分减少。
因而选择具有最佳生长性能的性别进行单性养殖, 有利于提高养殖对象的生产量和经济价值。
另外, 鱼类性别控制的研究, 对阐明鱼类性别分化和性别决定机制等理论问题, 也是非常有用的[]2。
1鱼类的性别鱼类的性别与其它性状一样,是受基因控制的。
染色体是基因的载体,因此,研究鱼类的染色体及其核型,有时可以发现控制性别的异形染色体,从而可判出某种鱼类性染色体类型。
但是,在目前所分析的约2000种鱼类染色体核型中,大部分并没有发现性染色体。
就目前所知的某些鱼类的性染色体类型,有的种类为XY 型,有的为ZW型,有的为20型,有的甚至具有多条性染色性。
为什么在现存的鱼类中,有的具有性染色体,有的却没有性染色体呢?有的专家认为,实际上这些鱼都具有性染色体,只是运用现有的生物学技术还不能准确地鉴别出来而已。
现已证实,鱼类除性染色体以外,常染色体上也具有影响性别的基因。
例如,尼罗罗非鱼的性别就不只由一对染色体决定,还与常染色体有关。
日本学者Yamazaki认为,大部分没有性染色体的鱼类,其常染色体所具有的雄性或雌性异配性别基因不仅可以使后代雌雄性别出现1:1的比率,而且还可产生雌雄同体的现象。
另外,还有人利用Southern印迹杂交的方法,证明了胡子鱿和斑马鱼基因组DNA均能与人的SRY(Sex一determin ingregiony)探针结合,表明了这两种鱼都存在SRY基因的同源序列。

鱼类两栖类的性别决黄扬光Hyghyg305@动物有无性生殖和有性生殖两种基本生殖方式,有性生殖与全部以有丝分裂为基础的无性生殖不同,雌雄两个体各自产生一个单倍体配子,两者结合成合子,合子发育成新个体。
有性生殖通过等位基因的分离、位于非同源染色体上的非等位基因的自由组合以及连锁基因的交换重组等过程为动物带来了巨大的遗传多样性,既提高了亲本生殖成功的机会,又有利于后代的生存。
多数动物都以雌雄两种互朴的性别形式存在,那么,性别是怎样决定的呢? 1性染色体决定性别自20世纪初发现某些生物体细胞内除含有成对的常染色体外,还有一对随性别不同组成也不同的性染色体。
这对性染色体与性别决定直接相关,性染色体决定性别有以下两种形式:1.1 XY型性别决定此类型在动物界很普遍,哺乳动物、双翅目昆虫、某些鱼类和某些两栖类的性别决定属于该类型。
XY型性别决定是指雌性为同配性别,体细胞内含有两条X染色体,所产生的性细胞只有一种,仅含一条X染色体;雄性为异配性别,体细胞内含一条X染色体和一条Y染色体,产生两种性细胞,一种只含X染色体,一种只含Y染色体。
如人的体细胞有46条染色体,其中22对为常染色体,一对为性染色体,男女之别在于女性性染色体是XX,男性性染色体为XY,婚后生男育女的概率各为1/2。
生男育女完全决定于受精的精于所含的性染色体是X还是Y。
XY型性别决定有一种特殊形式即XO型性别决定。
McClong在1902年就发现蝗虫的性别决定属XO型。
雌蝗虫是同配性别,只产生一种含X染色体的卵子。
雄蝗虫为异配性别,只含一条X染色体而无Y染色体,产生两种精子,一种只含一条X染色体,一种不含性染色体,两者各占一半。
受精卵有XX型(雌性)和XO(雄性)两种,雌雄比约为1:1。
1.2 ZW型性别决定鸟类、鳞翅目昆虫、某些鱼类、某些两栖类及某些爬行类属于该类型。
此类型的雄性为同配性别,体细胞内有两个相同的Z染色体,仅产生一种含Z染色体的精子。
鱼类性别发育和性别分化的分子机制鱼类的性别分化是一个非常复杂的过程,它涉及到一系列的分子机制,包括性别决定基因、性别逆转、雌雄激素等多个方面。
这些分子机制相互作用,共同掌控着鱼类性别的发育和分化。
1. 性别决定基因性别决定基因是指在不同的物种中,控制性别发育的基因。
在哺乳动物中,这个基因是SRY基因,而在鱼类中,则不止一个性别决定基因。
例如,虹鳟鱼和斑马鱼中的性别决定基因是dmrt1和gdf6,而草鱼和鲫鱼中则是amh和CYP19a1a。
这些性别决定基因的作用是通过调节下游基因的表达来控制性别分化。
在虹鳟鱼和斑马鱼中,dmrt1和gdf6基因分别调节睾丸和卵巢的发育。
而在草鱼和鲫鱼中,amh和CYP19a1a基因则分别调节雄性和雌性生殖细胞的发育。
2. 性别逆转性别逆转是指在性别分化过程中,由于外界环境的影响,使得一个本来应该是雌性的个体变为雄性或一个本来应该是雄性的个体变为雌性的现象。
在鱼类中,这个现象比较常见,例如泥鳅、鲤鱼和草鱼等品种都会出现性别逆转的情况。
性别逆转的原因是因为鱼类的性别分化过程受到环境因素的影响较大。
例如水温、光照、水质和饲料等环境因素会影响到鱼类的性腺发育和激素水平,从而导致性别逆转。
目前,科学家们已经发现了一些与性别逆转相关的基因,这些基因参与调节鱼类的性别发育过程,从而影响到性别逆转的发生。
3. 雄性激素和雌性激素雄性激素和雌性激素是影响鱼类性别分化的两种重要因素。
在鱼类中,雄性激素主要包括睾酮和11-酮雄酮,而雌性激素则主要包括雌二醇和17-酮雌酮。
这些激素可以通过直接调节下游基因的表达来影响到鱼类的性别分化,从而控制睾丸和卵巢的发育。
例如,在梭子蟹鱼和东方草鱼中,睾酮和11-酮雄酮可以抑制卵巢的发育,从而促进雄性的发育。
而在鲤鱼中,雌激素则可以促进雌性生殖细胞的发育,从而使得鱼体变成雌性。
总之,鱼类性别发育和性别分化的分子机制是一个复杂的过程,它涉及到多种基因、激素和环境因素的相互作用。
水产养殖中的养殖动物性别控制与繁殖技术水产养殖在满足人类食物需求方面起着至关重要的作用。
而在水产养殖过程中,养殖动物的性别控制与繁殖技术是一个重要的研究方向。
本文将探讨水产养殖中的性别控制技术、繁殖技术以及其对养殖业的影响。
读者将从中了解到水产养殖中性别控制与繁殖技术的重要性以及其应用场景。
一、性别控制技术1.1 基因控制法基因控制法是一种通过调控动物基因来实现性别控制的技术。
研究表明,在某些鱼类和软体动物中,存在一些性别决定基因,如具有Y染色体的个体为雄性,而没 Y 染色体的则为雌性。
通过对养殖动物基因进行调控,科学家们可以实现选择性别的目的。
例如,在鳗鲡养殖中,通过基因编辑技术成功实现了男性鳗鲡的繁殖,从而促进了养殖效益的提高。
1.2 温度控制法温度控制法是通过调控养殖环境的温度来影响动物性别的技术。
不同的温度对养殖动物的性别发育具有重要影响。
例如,在爬行动物中,卵的孵化温度可以决定幼体的性别。
适当调整温度可以实现性别的控制,这种方法被广泛用于鳄鱼、龟类等的养殖中。
然而,温度控制法对于不同种类的养殖动物可能存在差异,需要根据具体情况进行调整。
二、繁殖技术2.1 人工授精技术人工授精技术是一种通过人工手段将雄性的精子与雌性的卵子结合从而实现繁殖的方法。
这种技术在水产养殖中得到广泛应用。
例如,在鱼类养殖中,通过人工授精技术可以有效控制种间杂交,提高品种纯度和生长性能。
此外,人工授精技术还可以用于养殖动物种质资源的保存和遗传改良,对水产养殖业的发展具有重要意义。
2.2 催熟技术催熟技术是一种通过调控动物的生理状态来提高其繁殖能力的技术。
这种技术常用于增加养殖动物的繁殖频率和产仔量。
例如,在贝类养殖中,通过调控组织营养、养殖环境等因素,可以刺激贝类的繁殖行为,提高其繁殖效益。
催熟技术的应用可以有效促进水产养殖业的可持续发展。
三、性别控制与繁殖技术的影响性别控制与繁殖技术的应用对水产养殖业带来了诸多好处。
收稿日期:2001-08-20《淡水渔业》2002年第32卷第2期鱼类的性别决定和性染色体常重杰 杜启艳(河南师范大学生命科学学院,新乡453002) 全世界现存鱼类约有24618种,分属57目,482科,4258属,是脊椎动物中分布最广,种类最多的类群,具有多种多样的生物学特性和重大的经济价值。
在脊椎动物系统进化中,鱼类处于承先启后的地位,有着长久的进化历史和繁多演化分枝。
鱼类在进化过程中表现出各种趋同性、趋异性和保守性,变化纷繁,物种进化异常活跃[1]。
鱼类作为较低等的脊椎动物,在性别决定中也有多种表现形式,例如鱼类中有些物种是雌雄同体的,这在其它高等脊椎动物中是没有的。
即在同一个体中同时具有两种性腺,行自体受精。
而另有些物种则是连续性雌雄同体鱼类即一个个体先作为某一性别出现,在其生命周期的某个时刻再转化为另一性别,如黄鳝。
显然在雌雄同体鱼中只有性别的分化,而没有性别的遗传决定机制,然而在大部分鱼类中是存在真正的性别决定遗传机制。
深入研究其性别决定机制,对于水产养殖来说,具有重要的实用价值。
因为许多鱼类雌雄之间的经济性状如生长速度等存在差异,因此通过人为控制性别的方法专门生产全雌或全雄鱼苗,进行单性养殖将可以显著提高其经济效益。
同时,由于鱼类性染色体的进化尚处于早期阶段,广泛的研究其性染色体将有可能勾划出脊椎动物性染色体的演化历程。
经过数十年的艰苦探索,人们提出了多种识别鱼类性决定体系和性染色体的方法。
1 鱼类性别决定系统和性染色体的识别1.1 通过性连锁性状的遗传研究,判断性决定体系遗传研究表明在许多物种中,有一些表型性状是与性别联系在一起的。
鱼类中也不例外,并可根据其与性别连锁的关系进而推断其性决定体系。
Gubby (Poe cila reticulata )是一种小型鱼类。
其雄性比较鲜艳,体色有多种不同形式,而雌性则是单一灰色。
经典遗传研究证明,使雄性身体产生红色斑点,背鳍上出现黑色斑点是由Ma 基因控制的,这一性状限于雄性中;还有许多体色基因具有类似的遗传方式,由此说明Guppy 是XY 型性决定。
第22卷第2期2003年3月水 产 科 学FISHERIES SCIENCEVol122,No12Mar1,2003鱼类的性别决定与人工控制①刘良国[1,2],赵 俊2,崔 淼2(11常德师范学院生物系,湖南 常德 415000;21华南师范大学生命科学学院,广东 广州 510631)摘 要:鱼类性别的人工控制是鱼类育种中一个十分重要的领域。
本文概述了鱼类的性别决定机制及鱼类生理性别的表现方式,同时介绍了鱼类性别的多种人工控制方法,为育种单位的生产实践提供参考。
关键词:鱼类;性别决定;人工控制中图分类号:S969 文献标识码:A 文章编号:100321111(2003)022******* 在动物界中,雌雄个体之间在外部形态或生理上的差异是较为普遍的。
由于某些差异会带来极其重要的经济差别,因而认识动物的性别决定机制,以及如何控制动物的性别具有重大的意义。
许多鱼类雌雄个体之间存在着明显的生物学性状诸如生长率、成熟年龄、繁殖方式、体色、体型、个体大小等差异,而且大多数鱼类性成熟后生长速度显著下降,大部分能量消耗在性腺发育上,致使可食部分减少,肉质变差。
因此,人们可以根据需要专门生产全雌或全雄苗种进行单性养殖,或者培育出性腺不发育的中性鱼以消除性成熟带来的不利影响,从而大大提高养殖鱼类的经济效益。
很早以来,鱼类的性别决定机制以及鱼类性别的人工控制就一直引起人们的重视,发展迅速,有些研究成果已进入应用阶段,现就鱼类的性别决定及鱼类性别控制的研究进展作一介绍。
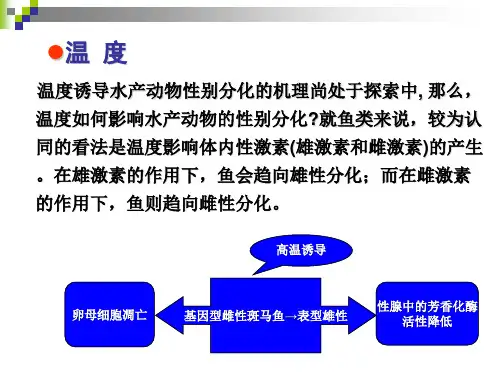
1 鱼类的性别决定在脊椎动物中,性别是由受精时两个配子所携带的性染色体所决定的,鱼类的性别决定亦是如此。
然而这种遗传性的决定并非一成不变,在鱼类胚胎发育过程中,原始的生殖腺同时具有向雌性和雄性发展的双向潜力,某些外部和内部因素,如水温、光照以及激素、代谢物等都可能通过某种途径改变机体的新陈代谢,特别是改变与性别决定有关的某些生理生化过程,从而影响鱼类的性别分化。
因此鱼类性别的表现方式多种多样,包括各种类型的性染色体机制、雌雄异体、雌雄同体及各种外部性别和行为性别等。
111 鱼类性别的染色体机制每种鱼都有它自己独特的染色体性别。
据统计,目前世界上大约有1600多种真骨鱼类进行过染色体组型的研究,但其中大多数鱼类用常规核型分析方法都难以分别出性染色体,已被证明具有性染色体机制的种类只有72种,能从细胞学上鉴定出性别的不超过30种。
鱼类中存在的与性别相关的异型性染色体类型有:XX/XY,XX/XO,ZW/ZZ,ZO/WW;复数性染色体也有报道,如X1X2X1X2/X1X2Y[1,2];有的鱼类存在复杂的性别决定系统,如在月鱼Xiphophorus m aculates(Poecilide,花鱼将科)中存在3个性染色体W、X、Y,此物种雌性个体的性染色体类型为WY、WX或XX,雄性个体的性染色体类型为XY或YY[3]。
我国目前已研究过的染色体鱼类中,只有12种具有异型性染色体,分布于进化历史不同的5个分类群(目)中(表1)。
Hammerman和Avtalion[5]根据Chen(1969)对罗非鱼不同种之间杂交和回交的实验结果,于1979年提出鱼类性别决定的“常染色体平衡理论”,该理论认为调节鱼类性别的基因位于性染色体和常染色体上,鱼类的性别是由染色体上雄性和雌性的基因产物数量优势来决定:如果雄性决定因素超过雌性决定因素,则雄性比例较高,反之,雌性决定因素超过雄性决定因素,则雌性比例较高。
在行天然雌核发育的银鲫、彭泽鲫等种群中存在少量的雄性比例,这一事实说明鱼类的常染色体似乎确有控制性别的基因存在。
表1 我国已研究过的具有异型性染色体的鱼类[4]目 种 名雌性雄性作 者鲤形目鲫Carassius aurat us auratas XX XY昝瑞光,1982银鲫C.a.gibelio XX XY鲶形目胡子鲶Clarias f uscus XX XY邬国民等,1986革胡子鲶zera XX XY斑点胡子鲶C.macrocephalus XX XT蟾胡子鲶C.bat rachus XX XY白缘Leiobagrus marginalus XX XY李康等,1985黑尾L.nigricauda XX XY拟缘L.marginatoides XX XT鲈形目刺鳅Mastacembelus aculeatus XX XY余先觉等,1989鳗鲡目星康吉鳗Conger myriaster ZW ZZ喻子牛等,1995鲱形目短颌鲚Colia brachygnathus ZO ZZ洪云汉等,1984①收稿日期:2002-09-29; 修回日期:2002-12-051作者简介:刘良国(1970-),男,讲师,现为华南师范大学生命科学学院2001级硕士研究生1主要从事鱼类系统进化和遗传育种的研究1致谢:本文撰写过程中承蒙陈湘麟教授的指导、修正,特此致谢! 综上所述,鱼类性别的染色体决定机制有以下特点:一、性染色体异型的分化不完美,大多数鱼类还难以分辨出性染色体;二、性染色体的分化发生于进化历史不同的分类群中,没有一个统一的模式;三、性别决定类型复杂,染色体可塑性强;四、常染色体上存在有与鱼类性别决定有关的基因。
112 鱼类的生理性别鱼类的生理性别又称表现型性别或生化性别,是在性别染色体基因控制下通过个体发育的生殖生理和生化过程而形成。
生理性别的基础取决于原始器官的类型。
大多数鱼类体内仅存精巢或卵巢,属雌雄异体(G onochorism )。
这类鱼可分为分化型和未分化型两种,前者最初的性腺直接分化成精巢或卵巢,如虹鳟、大麻哈鱼、青鱼将等属此类型;后者最初的性腺首先发育成卵巢样的性腺,以后一半个体发育成雄性,另一半则发育成雌性,许多鱼类被认为是未分化型的。
无论是分化型还是未分化型,雌雄异体的鱼类在外形上往往都很难辨别雌雄。
但也有一些鱼类存在雌雄异形现象,这种异形现象不仅表现为那些直接参与繁殖活动的体型特征,如软骨鱼类雄鱼的鳍脚、鱼雌鱼的产卵管、 鱼雄鱼的臀突等,有时还表现为不参与生殖活动而与生殖活动密切相关的特征,如婚姻妆、追星等。
通过组织形态学经典研究方法发现,目前有近400种鱼类在同一个体中出现雌雄两性性腺的现象,即雌雄同体(Hermaphroditism )。
雌雄同体鱼类主要存在于鲈形目的鱼旨科Serranidae 与鲷科Xenocyprinae 1养殖鱼类中较为常见的有黄鳝Monopterusalbus 和石斑鱼属Epinejphelus 中的一些种类。
早在1944年,刘建康就发现了黄鳝的雌雄同体现象,此后,刘修正等[6]对黄鳝性逆转时生殖腺的组织学与超微结构变化研究表明:雌性时期黄鳝的性腺中充满着不同发育期的卵母细胞;当进入雌雄间期时,滤泡细胞吞噬卵黄颗粒,卵母细胞退化吸收而产生空泡,这些空泡被新形成的生殖褶填充,褶上分布着精细胞;然后进入雄性时期,卵巢完全退化消失,曲细精管逐渐形成扩大,最终占领整个生殖腔。
因而一般认为雌雄同体是性逆转的一个过渡时期。
此外,鱼类的产卵行为和性行为方式也是生理性别的一种表现。
如大麻哈鱼雌鱼产卵前摆动尾鳍挖掘产卵坑,产卵后又用尾鳍扇动砂砾覆盖产卵坑的行为以及尼罗罗非鱼、莫桑比克罗非鱼、奥利亚罗非鱼的雌鱼口腔孵卵行为等。
2 鱼类性别的人工控制211 激素处理雄性激素可以将遗传上的雌鱼转变为生理表现型的雄鱼;雌性激素可以将遗传上的雄鱼转变成生理表现型雌鱼。
用来诱导人工雄性化的激素有很多,其中以17α-甲基睾酮(17α2methylu 2estosterone )应用最广泛,口服效果稳定而有效。
李家乐等[7]用含20×1026甲基睾酮的饲料投喂尼罗罗非鱼吉富品系,得到了100%全雄鱼,且成活率与生长率都比对照组显著提高;将遗传型雄鱼转变成生理型雌鱼,所用的雌激素主要是17β-雌二醇(estradial )和雌酮(estrone )。
如刘少军等[8]用一定剂量的雌二醇投喂革胡子鲶仔鱼一段时间后发现,除在性腺的表面残留少许精巢外,性腺绝大部分为卵巢,说明雌二醇在很大程度上已把遗传型雄性的个体转化为生理雌性。
性激素的处理方法一般都是投喂药饵或投喂与浸浴相结合。
开始处理时间一般都在仔鱼开口摄食之时,因为此鱼类性腺才开始分化。
激素处理持续时间因不同种类而异,一般约1~2个月。
此外,不同含量的甲酮对鱼类具有不同的生物效应。
低剂量(1×1026~5×10-6)可促进金鱼和鲤的生长[9,10],中剂量(10×1026~15×10-6)会导致金鱼或罗非鱼由雌性向雄性完全性转变[11,12],高剂量(100×10-6以上)可使异育银鲫产生雄激素阉割而变为中性不育鱼[13]。
从目前的研究进展来看,用较高剂量类固醇激素确实可以抑制性腺发育,因而可能是诱异不育鱼的一种有希望的方法。
但使用的浓度和处理时间将会严重地影响生长率和死亡率,必须严加控制。
事实上,生产中单纯用性激素大批量生产单性或不育鱼的方法很少采用,原因在于:一、药物来源困难,处理时间长,繁琐;二、对每一代鱼苗都必须进行处理,当代处理的鱼苗只能用于当代,不经济也不方便;三、直接把用激素处理过的鱼作为商品鱼养殖出售,可能会对人体有不利影响。
212 三系配套法以XX ♀2XY ♂型性别决定机制鱼类为例,三系配套法的具体操作方法是:先选择原系雄鱼(XY ♂),用雌激素诱导其转化为生理性雌鱼(XY “♂”);再用转化系(XY “♂”)与原系雄鱼(XY ♂)交配,获得雄性纯合系(超雄鱼,YY ♂);最后雄性纯合系再与正常原系雌鱼(XX ♀)交配获杂交全雄鱼(XY ♂)。
1955年,日本学者Y amamoto [14]用雌酮处理金鱼和青鱼将仔鱼获得转化系雌鱼(XY “♂”),然后与正常雄鱼(XY ♂)交配,在世界上首次获得超雄鱼(YY ♂)。
我国学者于70年代开始用三系配套法对莫桑比克罗非鱼(Tilapia mossambica )进行性别控制及单性养殖研究,杨永铨[15]用这种技术共获得YY 型超雄莫桑比克罗非鱼683尾,并用它们繁殖出了正常的全雄莫桑比克罗非鱼和莫桑比克罗非鱼与尼罗罗非鱼(T.nilotica )的杂交全雄鱼。
经实验测得,全雄莫桑比克罗非鱼比自然两性群体生长快4016%,群体产量平均提高4610%;全雄莫尼杂交种则有“全雄”和“杂种”的双重优势,其生长速度比尼罗罗非鱼两性群体快3815%,群体产量平均提高4314%。
与直接用激素控制性别的方法相比,采用三系配套法控制鱼类性别,所用激素不与商品鱼直接发生联系,具有用药量少,成本低,没有污染和便于推广等优点,已成为生产单性鱼的有效方法之一。
生产单位只需从制种单位购买少量的YY 型超雄鱼,就能与自然群体中的正常的雌鱼交配而得到大量的纯种全雄鱼或杂种全雄鱼。