3麂属动物的遗传变异和进化
- 格式:pdf
- 大小:327.33 KB
- 文档页数:8

动物变异知识点总结引言动物变异是指在动物种群中出现的个体性状或基因型的不同,这种不同可能是由于环境的影响,也可能是由于基因突变所导致的。
动物变异是自然界中的一种常见现象,它使得动物种群能够适应不断变化的环境,协助动物种群的进化和生存。
动物变异的原因动物种群中的变异主要由以下几种原因引起:1. 基因突变:动物的基因是决定其性状的物质基础,基因的突变可以导致动物性状的不同。
基因突变可能是由于自然辐射、化学物质或其他外界因素引起的。
2. 遗传变异:动物种群的遗传物质是动物性状遗传的基础,遗传物质的不同组合可以导致动物个体间性状的差异。
3. 外界环境:环境对动物个体的影响也是动物变异的重要原因,如气候、食物、生境等因素都会影响动物的性状变异。
动物变异的类型动物的变异是多种多样的,主要可以分为以下几种类型:1. 形态变异:动物个体的外部形态的不同,如体型大小、颜色、斑纹等。
2. 生理变异:动物个体的生理功能、生长发育等方面的差异,如抗病能力、生长速度、繁殖能力等。
3. 行为变异:动物个体的行为特征、活动习性等方面的不同,如捕食方式、4. 遗传变异:动物种群中不同个体之间遗传物质的差异,这种差异可能是由于不同基因的组合、基因突变等因素引起的。
5. 环境诱发变异:外界环境对动物个体的影响,导致其性状发生变异。
动物变异的意义动物变异对动物种群的生存和进化具有重要的意义:1. 适应环境:动物的变异使得个体能够更好地适应不断变化的环境,提高其生存能力。
2. 促进进化:动物种群中的变异为种群的进化提供了物质基础,使得种群能够适应新的生境、克服自然选择的压力,进化为更适应环境的种群。
3. 生物多样性:动物变异增加了种群内个体的差异性,有利于维持种群的多样性,维护生物多样性。
4. 科学研究:动物变异也为生物学、遗传学等科学研究提供了丰富的资料,为人们了解动物的生命规律提供了重要的信息。
动物变异的案例动物变异是自然界中的常见现象,下面我们列举一些动物变异的案例,以便更好地了解动物变异的特点和意义:1. 鸟类的颜色变异:鸟类的羽毛颜色可以发生变异,有些鸟类的羽毛呈现出奇特的色彩,如彩虹色的鸟、黑白相间的鹤、斑驳的松鸦等。


鹿科动物线粒体 DNA 概述及分子进化研究进展邵元臣;刘华淼;张然然;邢秀梅【摘要】线粒体DNA作为理想的分子遗传标记被广泛应用于鹿科动物种群遗传分析。
本文阐述了鹿科动物mtDNA结构和特点及各功能基因在鹿科动物物种识别、起源和进化、地理分化、遗传多样性等方面研究。
%Mitochondrial DNA as an ideal genetic marker has been successfully used in the research of evolutionary biology and population ge-netics deer .This article was aimed at discussing the structure and feature of mtDNA indeer .Furthermore ,emphasis was put on the applications in the fields of systematic evolution ,identification anddifferentiation ,phylogeography ,genetic diversity .【期刊名称】《特产研究》【年(卷),期】2014(000)002【总页数】4页(P67-70)【关键词】鹿;线粒体DNA;多态性;遗传多样性【作者】邵元臣;刘华淼;张然然;邢秀梅【作者单位】江苏科技大学,江苏镇江 212000; 中国农业科学院特产研究所,长春 130112;中国农业科学院特产研究所,长春 130112;中国农业科学院特产研究所,长春 130112;中国农业科学院特产研究所,长春 130112【正文语种】中文【中图分类】S825.21857年,瑞士生理学家Rudolf Albert在动物肌肉细胞中发现了线粒体;1962年,Nass发现,线粒体能够进行自我复制、转录、表达以及调控。
第3章麂属动物的遗传变异和进化麂属动物(Muntiacus)的细胞遗传学一直是国内外研究的热门课题,赤麂2n=6 ♀,7♂,是哺乳动物中已知染色体数目最少的种类。
麂属的不同种间染色体数目差异很大,其变异幅度远远超过哺乳动物中其他的属。
不同的种具有不同的核型特征,其演化过程很有规律。
在80年代后期,通过核型多态的研究又发现了一个新种——贡山麂(M.gongshanensis)(施立明,马彩霞 1988);去年以来,又有证据表明在越南、老挝边境地区又发现了两个新种,这在大型哺乳动物中是绝无仅有的。
3.1 贡山麂的新核型我们在开展中国麂属动物染色体进化与分子进化研究中,发现1只来自云南贡山县的雄性麂,无论其外部形态、头骨结构或核型特征,都与已知的5种麂有明显的不同。
为此,我们又在该地区捕获了1只雄性麂,进行了染色体制片和细胞遗传学观察,认为是一个新种。
这一看法得到了后来形态学研究的证实(马世来等 1990)。
说明了遗传学研究在动物分类学中可以起到积极的作用。
该雄性成年麂捕自云南省西部高黎贡山北段东坡,体重24kg,体长1 045mm,体背毛色暗褐,前额棕黄,无明显簇状冠毛,体侧和四肢呈暗灰黑色,腹部暗沙灰色。
我们以骨髓体外短期培养的方法制备细胞染色体标本,经100个以上中期骨髓细胞的观察计数,确定贡山麂的染色体数目为2n=9♂。
在根据染色体形态大小排列的常规核型中,Nos.1的一条同源染色体为亚中着丝粒,另一条为端着丝粒染色体,呈异形性。
Nos.2,3为亚中着丝粒染色体。
Nos.4为较小的端着丝粒染色体。
此外,核型中还有一小的亚中着丝粒染色体。
已知哺乳动物性染色体富于嗜银性并有特殊的形态和配对特征,在联会复合体分析时易于和常染色体区分鉴别(施立明1986)。
贡山麂精母细胞联会复合体分析表明,Nos.1的S.C 一端染色较深,即富嗜银性,也比较膨大,可以判断为X染色体和Y染色体配对形成短小的S.C。
在粗线期的不同阶段,配对的程度可以不同。
在早粗线期,XY靠拢,端部接近,在中粗线期,XY密切配对,在晚粗线期又开始分离。
这就证明,在Giemsa染色的体细胞核型中,亚中着丝粒的小染色体的确是Y。
Nos.1的S.C长臂实际上是常染色体。
看来,这条常染色体是端着丝粒形态,由于在进化过程中和X易位,就形成亚中着丝粒的复合染色体(XA)。
在C带核型中,可以看到每对染色体的着丝粒区都分布有大小不等的C带,即结构异染色质。
值得注意的是,在X和常染色体易位形成的复合染色体的“颈部”特别富于结构异染色质,这种特征的C带分布和预期的相吻合。
银染色结果表明,银染核仁组织者(Ag-NORs)即18s+28s核糖体基因分别出现于Nos._______________________本章作者:王文,兰宏,杨凤堂,马彩霞,张亚平20 中国动植物的遗传多样性1,4染色体的长臂。
电镜观察表明,Nos.1,4和S.C长臂附有清晰的核仁,和体细胞的银染结果相一致。
上述体细胞核型和减数分裂S.C分析表明,贡山麂的核型有其独特的特点。
已知小麂(M.reevesi)的2n=46(Wurster and Bernischke 1967),赤麂(M.mvaginalis)的2n=6♀,7♂(施立明 1976;Wurster,Bernirschke 1970),罗氏麂(M.rooseveltorum)的2n=7♂(Wurster 1985)无论是染色体的数目还是染色体形态都和贡山麂的明显不同。
分布于我国浙西、皖南地区的黑麂(M.crinifrons)的2n=8♀,9♂。
虽然两者的雄性染色体数目相同,但黑麂的一条Nos.1染色体为亚中着丝粒形态,已知也是X和常染色体易位形成的复合染色体,但另一条同源染色体却是典型的中着丝粒形态。
Nos.2染色体一条为亚中着丝粒形态,另一条同源染色体却是端着丝粒形态,这种复杂的染色体异形性涉及染色体易位和倒位(施立明 1983,1987)。
因此,黑麂的Nos.1,2染色体和贡山麂的也有明显的区别。
推测雌性贡山麂的2n=8,其染色体数目和形态可能和黑麂的相同或十分相似。
但银染的结果表明,虽然贡山麂和黑麂的Nos.4染色体长臂都带有Ag-NOR,但黑麂的另一对NOR分布于Nos.2染色体上,而贡山麂的另一对NOR分布于Nos.1染色体上,就是说这两种麂的核糖体基因的分布位置不同。
分布于马来西亚的赤麂属于指名亚种M.m.muntjak。
据Wurster和Atkin(1972)报道,其2n=8♀,推测雄性的2n=9♂,染色体数目和贡山麂相同,但通过核型的比较分析表明,虽然这两种麂的X都易位到一条常染色体上从而形成亚中着丝粒的复合染色体(XA),但这两种麂的这条染色体长度似有不同。
根据地理分布区域的不同,贡山麂也不太可能是赤麂的指名亚种M.m.muntjak。
此外,据昆明动物研究所兽类分类工作者的初步检测,贡山麂的外部形态与头骨特征和赤麂(M.muntjak)的差异较大。
Soma(1983)曾报道,泰国曼谷动物园一只雌性费氏麂(M.feae)的2n=13,因这只费氏麂是一个罗伯逊易位的携带者,故推测正常雌性动物的2n=14,雄性费氏麂的2n=15(Soma 1987)。
此外,除Nos.1为亚中着丝粒染色体外,核型中其他染色体都是端着丝粒形态。
我们的结果清晰地表明,贡山麂的核型无论是染色体数目,或是染色体形态都明显不同于原产缅甸南部和泰国南部的费氏麂。
综上所述,贡山麂的核型特征和现知的5种麂都有不同。
核型是细胞分类学的一个重要依据,特别是对于麂属(Muntiacus)更是如此(施立明 1980)。
因此,进一步收集更多的个体,并以多种染色体分带技术综合比较解剖学和分类学特征,深入研究分布于我国滇西北,包括藏南的贡山麂的细胞遗传学,确定其正确的分类地位——是新的亚种或是独立的新种,具有重要的意义。
3.2 麂属动物核型的染色体涂染比较麂属由于其核型多样性和一些种在进化中染色体重组率高而一直受到关注。
在迄今发现的哺乳类中,印度麂有最低数目的二倍体染色体数目(2n=6、7)(Wurster,Benirschke 1970);进一步研究此属的染色体数表明:黑麂(M.Crinifrons)2n=8、9(施立明 1983),贡山麂(M.gong shanensis)(施立明,马彩霞 1988)、雌性费氏种2n=13(M.feae 1983)和雄性费氏种2n=14(施立明未发表),中国麂(M.reevesi)2n=46(Wurster,Benirschke 1967)。
研究结果还表明印度麂和中国麂在核型上有20条不同染色体,但在表型第3章麂属动物的遗传变异和进化21上相似,两个种进行人工交配能产生幼仔,但后代是不育的(Shi,Pathak 1981),对这两个种的G-带比较研究表明,G-带有很高的同源性(施立明等 1980)。
此结果也支持Hsu(1975)等的看法,即印度麂染色体可能是多条染色体串联着丝粒融合的结果。
而且,通过对麂属mtDNA限制性片段长度多态分析表明,现代麂属所有种大约在200万年前发生分化,贡山麂和黑麂则不同,大约在60万年前分化(兰宏,施立明 1994)。
因此,麂属是研究核型进化和物种形成的一个很好的模型。
一些学者提出很多假说来证明串联融合理论,有两个证据支持此理论,即:①在细胞间期,中国麂的点状着丝粒非随机地聚集成束形成小的聚集体,表型上和在印度麂间期细胞核中观察到的复合着丝粒单元很相似(Brinkley等 1984);②中国麂的着丝粒区的异染色质C5序列和哺乳类端粒DNA序列在印度麂染色体上都有定位,而且,和其他麂属的G-带带型比较分析也支持此理论,即它们的核型有差异是由于祖先染色体的串联和着丝粒融合,但组合方式又有所不同(Fontana,Rubini 1990;施立明未发表)。
Elder和Hsu(1988)认为在中国麂基因组的进化过程中,其单倍数的染色体大约发生了17次染色体串联、3次着丝粒融合事件。
Lin等学者支持这一观点(1991),他在二倍染色体中检测到了30多个卫星DNA序列间质位点。
染色体涂染法是用有分类特征的染色体制成探针混合物,检测人类(Cremer等 1988;Lichter等 1988;Pinkel等 1988)、灵长类(Wienberg等 1990,1992;Jauch等 1992)和远缘哺乳类(Scherthan等 1994)在染色体进化中的细胞遗传学特征及复杂性和隐蔽的染色体重排现象。
令人庆幸的是,麂属染色体数目少而且比较大,用荧光活化细胞分类器就能把印度麂所有的染色体分开(Carrano等 1976;Levy等 1991)。
近来,随着染色体分类技术和多聚酶链式反应(PCR)技术的结合,特别是和退化低聚寡核苷酸引物PCR(DOP-PCR)的结合,能够用少数染色体快速地建立染色体特异文库和涂染点(Telenius等 1992)。
此技术也促进了反向染色体涂染法的发展,即将少数突变染色体片段经DOP-PCR产生涂染点探针,与正常中期分裂相杂交,以显示突变染色体的突变位点(Carter等 1992)。
1995年我们实验室与剑桥大学合作进行了染色体涂刷(chromosome painting)研究,以探讨麂属动物的染色体的进化方式(Yang等 1995)。
实验所用的雌性印度麂(KCB89003L)、雌性贡山麂(KCB92001)、雄性和雌性黑麂(KCB82001,KCB81002E)以及雄性中国麂(KCB92036)细胞株由中国科学院昆明动物所细胞库提供。
实验方法和结果如下。
3.2.1 增强DAPI带的核型和染色体组型模式图进行染色体涂刷,通常只能获得弱的DAPI带。
通过5×5高透滤光镜的增强作用,可提高DAPI带的强度。
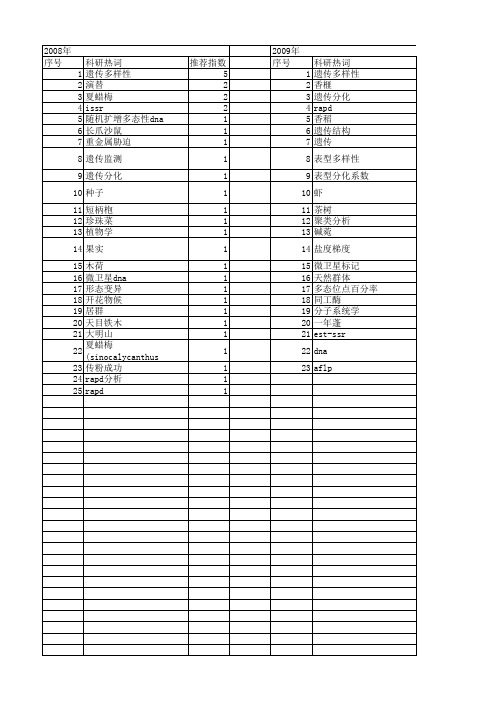
图3-1显示增强后雌印度麂、雌贡山麂、雌雄黑麂和雄中国麂的DAPI核型。
为了便于进行细胞遗传学分析,我们构建了印度麂、贡山麂、黑麂和中国麂的染色体组型模式图。
对于印度麂而言,增强的DAPI带核型是相同的,但并不同于Levy等(1993)所描述的G带染色体组型模式图。
特别是,我们发现1p3上有4条带,1q1上有7条带,而Levy 等则认为1p3上有6条带,1q1上有9条带。
增强后雌黑麂和贡山麂的DAPI带核型是相似的。
雌贡山麂的常染色体区域很大程度上相当于雌黑麂的如下区域:(X+1)q≈1q,2≈2,3≈3,4q≈(X+4)q,4p≈1p,X≈X,与贡山麂的1q,2p,(X+4)q,1p区各自相比时,发现(X+1)q区域缺少4条带,2p区多出4条带,4q区缺少2条带,而4p区又多出2条带。
在2q,3,和X染色体臂这些区域中,贡山麂和黑麂的带型是类似的。