《微生物学》主要知识点-08 第八章 微生物的遗传
- 格式:doc
- 大小:156.50 KB
- 文档页数:8


第八章微生物遗传和变异第一节微生物的遗传和变异遗传(heredity):亲代生物的性状在子代得到表现;亲代生物传递给子代一套实现与其相同形状的遗传信息。
特点:具稳定性。
遗传型(genotype):又称基因型,指某一生物个体所含有的全部基因的总和;------是一种内在可能性或潜力。
表型(phenotype):指生物体所具有的一切外表特征和内在特性的总和;------是一种现实存在,是具一定遗传型的生物在一定条件下所表现出的具体性状。
遗传型+ 环境条件= 表型表型是由遗传型所决定,但也和环境有关。
●变异:亲代与子代及子代个体之间,在形态结构和生理特性上的差异。
●微生物的变异分为遗传性变异和非遗传性变异。
●遗传性变异:是细菌的基因结构发生了改变,故又称基因型变异。
常发生于个别的细菌,不受环境因素的影响,变异发生后是不可逆的,产生的新性状可稳定地遗传给后代。
●非遗传性变异:细菌在一定的环境条件影响下产生的变异,其基因结构未改变,称为表型变异。
易受到环境因素的影响,凡在此环境因素作用下的所有细菌都出现变异,而且当环境中的影响因素去除后,变异的性状又可复原,表型变异不能遗传。
●遗传和变异是生物体的最本质的属性之一。
表型饰变:表型的差异只与环境有关特点:暂时性、不可遗传性、表现为全部个体的行为橘生淮南则为橘,生于淮北则为枳。
一微生物遗传变异的特点◆个体的体制极其简单;◆营养体一般都是单倍体;◆易于在成分简单的组合培养基上大量生长繁殖;◆繁殖速度快,生活史周转快,容易发生变异;◆易于积累不同的中间代谢产物或终产物;◆菌落形态特征的可见性和多样性;◆环境条件对微生物群体中各个个体作用的直接性和均一性;◆易于形成营养缺陷型;◆各种微生物一般都有相应的病毒;◆存在多种处于进化过程中的原始有性生殖方式;微生物是遗传学研究中的明星:●微生物细胞结构简单,营养体一般为单倍体,方便建立纯系。
●很多常见微生物都易于人工培养,快速、大量生长繁殖。

微生物遗传知识点总结一、微生物的遗传物质1.DNA:微生物的遗传物质主要是DNA(脱氧核糖核酸),DNA是微生物的基因组主要组成部分,承载了微生物的遗传信息。
2.RNA:微生物的遗传物质中还包括RNA(核糖核酸),RNA在微生物的蛋白质合成中起到重要的作用,有mRNA、tRNA和rRNA等不同类型。
3.质粒:微生物的遗传物质中还存在质粒,质粒是细胞外遗传物质,可以自主复制和传递,在微生物的分子遗传研究中具有重要的意义。
二、微生物的遗传变异1.突变:突变是指微生物遗传物质的突发性变异,包括点突变、插入突变和缺失突变等,突变会导致微生物表型的变化,包括对抗药物的耐药性等特征。
2.重组:重组是指微生物遗传物质的重组和重排,包括同一基因组内的DNA重组和来自不同基因组的DNA重组,重组可以导致各种遗传特征的变异和产生新的遗传组合。
3.外源基因的导入:微生物可以通过外源基因的导入来获得新的遗传特征,包括外源DNA的转化、噬菌体的侵染和质粒的转移等方式。
三、微生物的遗传传递1.垂直传递:垂直传递是指微生物遗传物质从父代到子代的传递,包括细菌的有丝分裂、芽生、孢子形成和病毒的感染传递等方式。
2.水平传递:水平传递是指微生物遗传物质在同一代的微生物个体之间的传递,包括细菌的共享基因池、DNA转化和连接转移等方式,可以导致微生物之间的基因交换和遗传多样性的增加。
四、微生物遗传的调控机制1.DNA修饰:微生物可以通过DNA修饰来调控基因的表达,包括DNA 甲基化和DNA腺苷酸修饰等方式,这些修饰可以影响基因的转录和翻译过程。
2.转录调控:微生物可以通过转录因子的结合和解离来调控基因的转录水平,包括正调控和负调控,这些调控作用可以响应内外环境的变化。
3.蛋白质修饰:微生物可以通过蛋白质的修饰来调控蛋白质的活性和稳定性,包括翻译后修饰和酶的磷酸化、乙酰化和甲基化等方式。
4. RNA干涉:微生物可以通过RNA干涉机制来调控基因表达,包括小分子RNA的介导和crispr-cas系统等方式,这些机制可以抑制或靶向性地破坏特定基因的表达。






第八章微生物的遗传概述:遗传(heredity or inheritanc® 和变异(variation)是生物体的最本质的属性之一。
遗传即生物的亲代将一整套遗传因子传递给子代的行为或功能。
变异指生物体在某种外因或内因的作用下所引起的遗传物质结构或数量的改变。
基因型(ge no type某一生物个体所含有的全部基因的总和。
表型(phe no type)某一生物所具有的一切外表特征及内在特性的总和。
饰变( modification)不涉及遗传物质结构改变而发生在转录、翻译水平上的表型变化。
8.1遗传变异的物质基础8.1.1三个经典实验1. 经典转化实验:1928年F.Griffith以Streptococcus pneumoniae为研究对象进行转化(transformation)实验。
1944年O.T.Avery等人进一步研究得出DNA是遗传因子。
S strun A2. 噬菌体感染实验:1952年Alfred D.Hershey和Martha Chase用32P标记病毒的DNA,用35S标记病毒的蛋白质外壳,证实了T2噬菌体的DNA是遗传物质。
3.植物病毒的重建实1956年H.Fraenkel-Conrat用含RNA的烟草花叶病毒(tobacco mosaic virus,TMV)与TMV 近源的霍氏车前花叶病毒(Holmes ribgrass mosaic virus,HRV)所进行的拆分与重建实验证明,RNA也是遗传的物质基础。
8.2微生物的基因组结构:基因组(genome是指存在于细胞或病毒中的所有基因。
细菌在一般情况下是一套基因,即单倍体(haploid);真核微生物通常是有两套基因又称二倍体(diploid )。
基因组通常是指全部一套基因。
由于现在发现许多非编码序列具有重要的功能,因此目前基因组的含义实际上是指细胞中基因以及非基因的DNA序列的总称,包括编码蛋白质的结构基因、调控序列以及目前功能还尚不清楚的DNA序列。
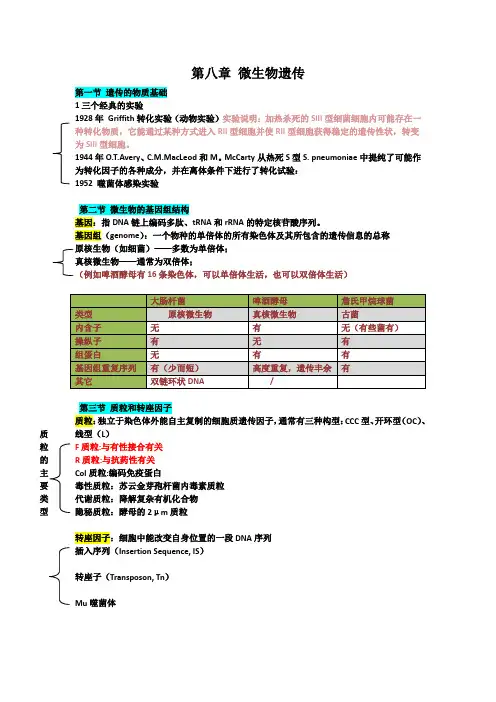
第八章 微生物遗传第一节 遗传的物质基础 1三个经典的实验1928年 Griffith 转化实验(动物实验)实验说明:加热杀死的SIII 型细菌细胞内可能存在一种转化物质,它能通过某种方式进入RII 型细胞并使RII 型细胞获得稳定的遗传性状,转变为SIII 型细胞。
1944年O.T.Avery 、C.M.MacLeod 和M 。
McCarty 从热死S 型S. pneumoniae 中提纯了可能作为转化因子的各种成分,并在离体条件下进行了转化试验: 1952 噬菌体感染实验第二节 微生物的基因组结构基因:指DNA 链上编码多肽、tRNA 和rRNA 的特定核苷酸序列。
基因组(genome ):一个物种的单倍体的所有染色体及其所包含的遗传信息的总称 原核生物(如细菌)——多数为单倍体; 真核微生物——通常为双倍体;(例如啤酒酵母有16条染色体,可以单倍体生活,也可以双倍体生活)第三节 质粒和转座因子 质粒:独立于染色体外能自主复制的细胞质遗传因子,通常有三种构型:CCC 型、开环型(OC )、线型(L )F 质粒:与有性接合有关 R 质粒:与抗药性有关 Col 质粒:编码免疫蛋白毒性质粒:苏云金芽孢杆菌内毒素质粒 代谢质粒:降解复杂有机化合物 隐秘质粒:酵母的2μm 质粒转座因子:细胞中能改变自身位置的一段DNA 序列 插入序列(Insertion Sequence, IS )转座子(Transposon, Tn )Mu 噬菌体质粒 的 主 要 类 型转座引起的遗传效应插入突变产生染色体畸变基因的移动和重排第四节基因突变和修复4.1突变类型同义突变错义突变无义突变移码突变营养缺陷型抗性突变型条件致死突变型形态突变型抗原突变型产量突变型4.2自发突变——分子基础:碱基的互变异构---转换和颠换4.3诱发突变碱基结构类似物胸腺嘧啶类似物-----5-溴尿嘧啶腺嘌呤类似物--------2-氨基嘌呤插入染料溴化乙锭、吖啶橙诱变剂亚硝酸、羟胺类化合物、烷化剂辐射和热紫外线-----嘧啶二聚体生物诱变因子:转座因子4.4 诱变剂与致癌物质——Ames实验4.5 DNA损伤的修复光复活作用:光复合酶直接修复切除修复重组修复SOS修复第五节细菌的基因转移和重组转化(transformation)结合(conjugation,mating)转导(transduction)5.1 转化定义:受体细胞直接吸收了来自供体细胞的DNA片断,并把它整合到自己的基因组中,细胞部分遗传性状发生变化的现象叫转化。

微生物学主要知识点08微生物的遗传微生物的遗传是微生物学中的一个重要知识点,包括微生物的基因组结构、遗传物质的复制和转录、重组以及突变等方面。
了解微生物的遗传不仅可以帮助科学家研究微生物的进化和适应能力,还可以应用于微生物的工业生产和疾病防治等领域。
1.微生物的基因组结构:微生物的基因组由DNA组成,DNA通过多个螺旋体嵌入细胞的细胞核或质粒中。
微生物的基因组可以分为染色体和质粒两部分,质粒是一种较小的环状DNA。
染色体和质粒中都含有基因,基因通过编码蛋白质的方式决定了微生物的特征和功能。
2.遗传物质的复制和转录:微生物的DNA通过复制和转录的方式进行遗传物质的复制。
DNA复制是指将一个DNA分子复制成两个完全相同的DNA分子,从一个细胞传递到另一个细胞。
DNA转录是指根据DNA模板合成RNA的过程,RNA复制的结果是生成一个与DNA模板相对应的RNA分子。
这些RNA分子可以进一步转录成蛋白质。
3.重组:微生物的重组是指在微生物遗传物质中发生DNA片段的重新组合。
这种重组可以发生在同一染色体上的两个相同或不同的DNA片段之间,也可以发生在不同染色体或质粒之间。
微生物的重组有助于增加遗传多样性,并提高微生物的适应能力和进化速度。
4.突变:微生物的遗传中还会发生突变现象,突变是指DNA序列的改变。
突变可以是点突变,即DNA中的一个碱基替换为另一个碱基;也可以是插入和缺失,即DNA序列中添加或删除一个或多个碱基。
突变可能对微生物的生长和繁殖产生负面影响,也可能带来新的适应优势。
5.横向基因转移:微生物的遗传中还存在横向基因转移的现象。
横向基因转移是指将一个细胞(供体)中的基因转移到另一个细胞(受体)中,无需通过细胞分裂进行。
横向基因转移可以发生在同一物种的细菌之间,也可以发生在不同物种的细菌之间。
横向基因转移是微生物进化和适应性演化的重要驱动因素之一6.基因调控:微生物的基因表达受到一系列调控机制的控制。
第八章微生物的遗传概述:遗传(heredity or inheritance)和变异(variation)是生物体的最本质的属性之一。
遗传即生物的亲代将一整套遗传因子传递给子代的行为或功能。
变异指生物体在某种外因或内因的作用下所引起的遗传物质结构或数量的改变。
基因型(genotype)某一生物个体所含有的全部基因的总和。
表型(phenotype)某一生物所具有的一切外表特征及内在特性的总和。
饰变(modification)不涉及遗传物质结构改变而发生在转录、翻译水平上的表型变化。
8.1 遗传变异的物质基础8.1.1 三个经典实验1. 经典转化实验:1928年F.Griffith以Streptococcus pneumoniae为研究对象进行转化(transformation)实验。
1944年O.T.Avery等人进一步研究得出DNA是遗传因子。
2.噬菌体感染实验:1952年Alfred D.Hershey和Martha Chase用32P标记病毒的DNA,用35S标记病毒的蛋白质外壳,证实了T2噬菌体的DNA是遗传物质。
3.植物病毒的重建实验:1956年H.Fraenkel-Conrat用含RNA的烟草花叶病毒(tobacco mosaic virus,TMV)与TMV近源的霍氏车前花叶病毒(Holmes ribgrass mosaic virus,HRV)所进行的拆分与重建实验证明,RNA也是遗传的物质基础。
8.2 微生物的基因组结构:基因组(genome)是指存在于细胞或病毒中的所有基因。
细菌在一般情况下是一套基因,即单倍体(haploid);真核微生物通常是有两套基因又称二倍体(diploid)。
基因组通常是指全部一套基因。
由于现在发现许多非编码序列具有重要的功能,因此目前基因组的含义实际上是指细胞中基因以及非基因的DNA序列的总称,包括编码蛋白质的结构基因、调控序列以及目前功能还尚不清楚的DNA序列。
微生物基因组随不同类型表现出多样性。
8.2.1大肠杆菌的基因组:大肠杆菌基因组为双链环状的DNA分子。
在细胞中以紧密缠绕成的较致密的不规则小体(拟核,nucloid)形式存在于细胞中,其上结合有类组蛋白蛋白质和少量RNA分子,使其压缩成脚手架形的(scaffold)致密结构(大肠杆菌DNA分子长度是其菌体长度的1000倍,必须以一定的形式压缩进细胞中)。
基因组全序列测定于1997年由Wisconsin大学的Blattner 等人完成。
大肠杆菌基因组结构特点:1.遗传信息的连续性:个别细菌(鼠沙门氏菌和犬螺杆菌)和古生菌的rRNA 和tRNA 含有内含子或间隔序列,其他绝大部分原核生物不含内含子,遗传信息是连续的而不是中断的。
2.功能相关的结构基因组成操纵子:大肠杆菌共有2584个操纵子,基因组测序推测出2190个操纵子。
如此多的操纵子结构可能与原核基因表达多采用转录调控有关。
此外,有些功能相关的RNA基因也串联在一起,如构成核糖体的三种RNA基因转录在同一转录产物中,依次是16S rRNA-23S rRNA-5S rRNA 。
三种RNA在核糖体中的比例是1∶1 ∶1。
3.结构基因的单拷贝及rRNA 基因的多拷贝:大肠杆菌有7个rRNA操纵子其特征都与基因组的复制方向有关即按复制方向表达。
7个rrn 就有6个分布在双向复制起点oric(83min)附近,有利于核糖体的快速组装。
4.基因组的重复序列少而短:原核生物基因组存在一定数量的重复序列,但比真核生物少得多,重复序列一般为4~40 bp,重复程度十多次、上千次不等。
8.2.2 啤酒酵母的基因组:啤酒酵母是单细孢真核生物,1997年,有欧洲、美国、加拿大和日本共96个实验室的633为科学家是艰苦努力完成了全基因组的测序工作。
是第一个完成测序的真核生物基因组。
基因组大小13.5×106 bp,分布在17个不连续的染色体中。
酵母菌的DNA与四种主要的组蛋白(H2A 、H2B、H3 、H4)结合构成染色质(chromatin)的14bp核小体核心DNA;染色体DNA 上有着丝粒(centromere)和端粒(telomere)没有明显的操纵子结构,有间隔区或内含子序列。
酵母菌基因组的特点:高度重复;tRNA有250个拷贝。
rRNA 只位于Ⅻ号染色体的近端粒处,每个长9137 bp,有100~200个拷贝。
较高同源性的DNA重复序列称为遗传丰余(genetic redundancy)——一种进化的策略(有备无患)。
8.2.3 詹氏甲烷球菌的基因组:詹氏甲烷球菌(Methanococcus jannaschii)属于古菌,发现于1982年。
生活在2600m深,2.63×107 Pa(260个大气压),94℃的海底火山口附近。
1996年由美国基因组研究所(The Institute for Genomic Research,简称TIGER)和其他5个单位共40人联合完成了该菌的基因组全测序工作。
这是完成的第一个古菌和自养型生物的基因组序列。
根据对该菌全基因组序列分析结果完全证实了1977年由Woese等人提出的三界学说。
詹氏甲烷球菌基因组的特点:詹氏甲烷球菌只有40%左右的基因与其他二界生物有同源性,其中有的类似于真细菌,有的类似于真核生物,有的二者融合。
古菌在基因组结构上类似于细菌。
詹氏甲烷球菌有 1.66×106bp的环形染色体DNA,具有1682个编码蛋白质ORF;功能相关的基因组成操纵子共转录成一个多顺反子转录子;有2个rRNA 操纵子;37个tRNA 基因,基本上无内含子;无核膜。
负责信息传递的基因类似于真核生物。
转录起始系统、RNA聚合酶的亚基组成及序列、启动子结构、翻译延伸因子、复制起始因子均与真核生物相似。
古菌还含有5个组蛋白基因。
8.3 质粒和转座因子:质粒(plasmid)和转座因子(transposable element)都是细胞中除染色体以外的另外两类遗传因子。
前者是一种独立于染色体外,能进行自主复制的细胞质遗传因子,主要存在于各种微生物细胞中;后者是位于染色体或质粒上的一段能改变自身位置的DNA序列,广泛分布于原核和真核细胞中。
8.3.1 质粒1.质粒的分子结构:通常以共价闭合环状(covalently closed circular,CCC)的超螺旋双链DNA分子存在于细胞中,从细胞中分离的质粒大多是三种构型即CCC型、OC型(open circular)、L型(linear form)。
近年来在疏螺旋体、链霉菌和酵母菌中也发现了线形双链DNA质粒和RNA质粒。
质粒分子大小范围 1 kb~1000 kb。
2.质粒的主要类型:质粒所含有的基因对宿主细胞一般是非必须的,只是在某些特殊条件下,质粒能赋予宿主细胞以特殊的机能,从而使宿主得到生长优势。
根据质粒所编码的功能和赋予宿主的表型效应,可将其分为不同的类型。
1、致育因子(fertility factor,F因子):又称F质粒,大小约100kb,是最早发现的一种与大肠杆菌的有性生殖现象(结合作用)有关的质粒。
2、抗性因子(resistance factor,R因子):包括抗药性和抗重金属两大类。
3、Col质粒(colicinogenic factor):产大肠杆菌素因子,能编码大肠杆菌素(colicin)属于细菌毒素(bacteriocin)。
4、Ti质粒(tumor inducing plasmid )5、代谢质粒(metabolic plasmid )6、隐秘质粒(cryptic plasmid ):不显示任何表型效应。
只有通过物理的方法,如凝胶电泳检测细胞抽提液等方法才能发现。
酵母的2µm 质粒。
根据质粒的拷贝数、宿主范围分:高拷贝数质粒(high copy number plasmid ):每个宿主细胞中可以有10~100个拷贝。
又称松弛型质粒(relaxed plasmid )。
低拷贝数质粒(low copy number plasmid ):每个宿主细胞中可以有1~4个拷贝。
又称严谨型质粒(stringent plasmid)。
窄宿主范围质粒(narrow host range plasmid ):复制起始点(origin of replication)较特异。
广宿主范围质粒(broad host range plasmid ):复制起始点不太特异。
附加体(episome):能整合进染色体而随染色体的复制而进行复制且又能脱离的质粒。
3.质粒的不亲和性(incompatibility):细菌通常含有一种或多种稳定遗传的质粒,这些质粒即为彼此亲和的(compatible)。
如果将一种类型的质粒通过接合或其他方式(转化)导入某一合适的但已含另一种质粒的宿主细胞,只经少数几代后,大多数子细胞只含有其中一种质粒,那么这两种质粒是不亲和的(incompatibility )。
根据某些质粒在同一细菌中能否并存的情况,可将质粒分成许多不亲和群(incompatibility group),能在同一细菌中并存的质粒属于不同的不亲和群,而在同一细菌中不能并存的质粒属于同一不亲和群。
当两种同一不亲和群的质粒共处同一细胞时,其中一种由于不能复制因而在细胞的不断分裂过程中被稀释(diluted out)或被消除(curing)。
8.3.2 转座因子:转座因子(transposable element)是细胞中能改变自身位置的一段DNA序列。
广泛存在于原核和真核细胞中。
原核和真核生物中的转座因子原核生物中的转座因子有三种类型:插入序列(insertion sequence,IS);转座子(transposon,Tn);某些特殊病毒(Mu、D108)。
转座的遗传学效应:1、插入突变;2、产生染色体畸变;3、基因的移动和重排。
8.4 基因突变和诱变育种8.4.1 基因突变(gene mutation):生物体内遗传物质的分子结构发生的可遗传的变化。
1、突变类型:营养突变型(auxotroph);抗性突变型(resistant mutant);条件致死突变型(conditional lethal mutant);形态突变型(morphological mutant);抗原突变型(antigenic mutant);产量突变型。
2、突变率(mutation rate):每一细胞在每一世代中发生某一性状突变的几率。
突变率一般为10-6~10-9。
3、突变的特点:不对应性、自发性、稀有性、独立性、诱变性、稳定性、可逆性。
4、基因突变的自发性和不对应性的证明:变量试验(fluctuation test);涂布试验(Newcombe experiment);平板影印培养试验(replica plating)。