第七章 废水生物化学处理基础
- 格式:doc
- 大小:242.50 KB
- 文档页数:13


废水生化处理基础第一讲废水处理物理、化学、生物法优缺点分析1、污水处理的物理法污水处理的物理法是通过沉淀,过滤处理,净化污水。
优点:不需要害怕会残留化学物质(污水处理所用的)、物理法速率较快,只是准备工作较多。
缺点:可能会处理的不干净。
2、污水处理的化学法污水处理的化学法是指向废水中加入化学药剂如明矾等化学药品,使其与污染物发生化学反应而生成无害物的过程。
优点:化学法不必基建、且原污水中的物质处理的干净、时间周期较长,可持续性不错。
缺点:运行期间需要添加化学药剂,可能会残留化学物质(污水处理所用的)、前期准备周期慢。
3、污水处理的生物法污水处理的生物法是利用微生物降解代谢有机物为无机物来处理废水。
通过人为的创造适于微生物生存和繁殖的环境,使之大量繁殖,以提高其氧化分解有机物的效率。
优点:与化学法相比,微生物处理法具有经济、高效的优点,并可实现无害化、资源化,所以长期以来始终占重要位置。
缺点:生物法需要基建,占地面积大,水量比较大的话合适。
从污水处理的历史来看,早期的污水处理都是物理法、化学法,简单的说就是机械隔离,投加絮凝剂,物理自由沉淀,但是这样的办法导致药耗量很大,污泥量极大,所以之后的污水处理普遍转向生物法,即通过活性污泥来分解污水中的有机物。
污水处理厂分一级、一级强化、二级、三级等,只有在二级及以后的才是采取生物法。
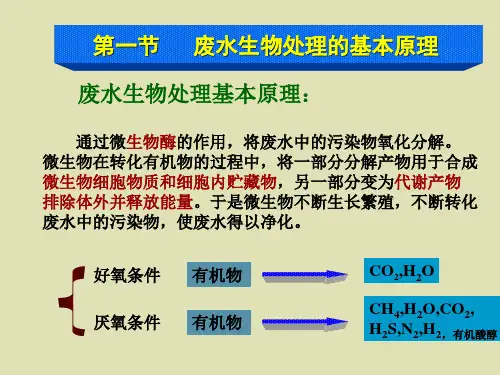
第二讲废水生物化学处理基础1、在好氧条件下,废水中有机物的去除主要是由哪几个生物过程完成的?请分别给出其反应方程式。
(1) 分解反应(氧化反应、异化代谢、分解代谢)CHONS(有机物的组成元素)+O2→CO2+H2O+NH3+SO42-+…+能量(2) 合成反应(合成代谢、同化作用)CHONS+能量→C5H7N02(细胞物质)(3) 内源呼吸(细胞物质的自身氧化)C5H7N02+ O2→CO2+H2O+NH3+SO42-+…+能量有机物质在生物体细胞内氧化分解产生二氧化碳、水,并释放出大量能量的过程称为生物氧化。


废水生化处理理论基础废水处理是指对工业、农业、生活等生产和生活活动中所产生的废水进行处理,将废水中的各种有害物质去除或降低,使其达到环境排放标准,保护环境、维护生态平衡。
废水处理技术较为复杂,其中生化处理是一种常用的处理方法。
本文将介绍废水生化处理的理论基础。
1. 废水生化处理概述废水生化处理是利用微生物的生物化学作用,将有机物质降解成较为稳定、不易污染环境的无机物质,以实现对废水的净化处理。
生化处理一般包括好氧生物处理和厌氧生物处理两种方式。
•好氧生物处理:好氧生物处理是指在充氧的条件下,利用好氧微生物将废水中的有机物质氧化分解为二氧化碳和水。
这种处理方式对细菌的要求较高,需要提供足够的氧气。
•厌氧生物处理:厌氧生物处理是指在没有氧气的条件下,利用厌氧微生物将废水中的有机物质降解成沼气、二氧化碳等产物。
这种处理方式对微生物的适应能力要求较高,处理效果也较好。
2. 废水生化处理原理废水生化处理的基本原理是将废水中的有机物质通过生物作用转化为无机物质。
有机物质能够为微生物提供能量和生长所需的碳、氮、磷等元素,而微生物则通过代谢作用将有机物质降解为无机物质。
生化处理的主要过程包括:•底物的降解:微生物利用底物(有机物质)作为碳源和能源,在水体中进行降解反应,生成底物降解产物和生物体。
•底物的转化:底物降解产物经过一系列酶类的作用,逐步转化为无害的终产物,如CO2、H2O等。
•生物体的生长:底物的降解还伴随着微生物的生长和繁殖,微生物的数量和种类变化也会影响处理效果。
3. 废水生化处理的关键技术废水生化处理的关键技术包括微生物培养、废水处理工艺设计、氧气供给等方面。
其中,微生物在生化处理中扮演着重要的角色,其培养和管理对处理效果至关重要。
•微生物培养:合理选择适应性强、活性高的微生物种类,进行培养和管理,提高其降解效率和处理能力。
•工艺设计:根据废水特性和处理要求设计合理的生化处理工艺,包括反应器设置、曝气方式、混合方式等。




废水生化处理理论基础废水生化处理理论基础第一节废水处理微生物基础一般说来,废水中所含的污染物质是多种多样的,因此不能期望只用一种处理方法就能把所有的污染物质去除殆尽,往往需要由几种方法组成一个处理系统,才能完成所要求的处理功能。
废水生化处理是利用生物的新陈代谢作用,对废水中的污染物质进行转化和稳定、使之无害化的处理方法。
对污染物进行转化和稳定的主体是微生物。
所谓微生物是肉眼不能看见,只能凭借显微镜才能观察到的单细胞及多细胞生物。
从狭义角度说主要是指菌类生物,包括细菌、放线菌、真菌以及病毒等。
从广义角度说,除了菌类生物及病毒外;还包括藻类、原生动物和一些后生动物。
由于微生物具有来源广、易培养、繁殖快、对环境适应性强、易变异等待性,在生产上能较容易地采集菌种进行培养增殖,并在特定条件下进行驯化,使之适应有毒工业废水的水质条件,从而通过微生物的新陈代谢使有机物无机化,有毒物质无害化。
加之微生物的生存条件温和,新陈代谢过程中不需高温高压,它是不需投加催化剂的催化反应,用生化法促使污染物的转化过程与一般化学法相比优越得多,其处理废水的费用低廉,运行管理较方便;所以生他处理是废水处理系统中员重要的过程之一,目前,这种方法已广泛用作生活污水及工业有机废水的二级处理。
一、微生物的新陈代谢微生物在生命活动过程中,不断从外界环境中摄取营养物质,并通过复杂的酶催化反应将其加以利用,提供能量并合成新的生物体,同时又不断向外界环境排泄废物。
这种为了维持生命活动过程与繁殖下代而进行的各种化学变化称为微生物的新陈代谢,简称代谢。
各种生物的生命活动,如生长、繁殖、遗传及变异,都需要通过新陈代谢来实现,可以说,没有新陈代谢,就没有生命。
根据能量的释放和吸取,可将代谢分为分解代谢和合成代谢。
在分解代谢过程中,结构复杂的大分子有机物或高能化合物分解为简单的低分子物质或低能化合物,逐级释放出其固有的自由能,微生物将这些能量转变成三磷酸腺苷(ATP),以结合能的形式储存起来。


第七章废水生物化学处理基础本章重点:如何建立单个细菌以及生物膜或生物絮体的数学模型。
1947年,首次出现了“生物化学工程”( Biochemical engineering)一词。
1965年Aiba等人的专著《物化学工程》(Biochemical Engineering)出版,标志着这一学科的正式出现。
1971年Coulson及Richardson等著述的化学工程标准教材新添了第三卷,其中包括了一章生物化学反应工程,标志着生物化学工程已成为化学工程的—个新的组成部分。
此后出版的生物化学工程专著有Atkinson的《生物化学反应器》(Biochemical Reactors,1974年),Bailey及ollis 的《生物化学工程基础》(Biochemical Engineering Fundamentals.1977年)等书。
生物化学工程中应用的发酵器有两种基本类型,一种是利用微生物絮体的作用,这与废水处理中的活性污泥法相类似;另一种是利用微生物膜的作用,这与废水处理中的生物滤池法相类似。
以生物化学工程的方法来研究废水的生物处理,提高了它的理论深度,应该是发展的方向。
把废水的生化处理看成是生物化学工程的一个重要分支,在学科体系上可能更合适—些。
§7.1 单个细菌的模型从细菌结构及代谢途径来看,如果要按实际情况建立一个数学模型,几乎无法着手。
所以目前一般采用一个远为简化的模型,而这个模型也起到了对营养物传入细菌内的整个过程,给出明确概念的作用。
底物一般是通过细胞的粘液层、细胞壁与细胞膜进入细胞内部的,而代谢作用只发生在细胞内部的细胞质区。
发生代谢作用后,底物也就消失了。
这里,我们假设:①不考虑复杂的代谢过程;②把底物的消失引用流体力学中“汇”的概念来解释;③粘液层、细胞壁、细胞膜等作为底物传递的边界。
这样就得到一个细菌的简化模型,如图7-1所示。
扩散区指细胞壁外粘液层的部分,其表面积为a d cm 2,,底物通过扩散区时服从Fick 的第一扩散定律,即底物的通量为:Nd = -Dγρd d (7-1) 式中,下标d 表示扩散区,γρd d 表示晏半径γ方向的浓度梯度,D 仍然表示分子扩散系数。
第一部分废水处理基础知识绪论1、简述废水的来源?按其来源可分成:生活污水、工业废水和降水。
2、废水的物理性质和化学性质包括哪些内容?废水的物理性质有温度、气味、色度、悬浮物和流量变化。
化学性质有:有机化合物、无机物和气体等。
3、BOD5\COD cr\TOC的定义?BOD5\是指1升废水中的有机污染物在好氧微生物作用下进行氧化分解所消耗的溶解氧量。
BOD5\即水样在20℃条件下培养5天的生化需氧量。
COD ccr是指用重铬酸钾强氧化剂使被测水中的有机污染物进行化学氧化时所消耗的氧量。
TOC系指废水中所有有机物的含碳量。
4、简述水污染控制的途径?①改革工艺,减少废水产生,也就是优先采用无污染或少污染的新工艺、新设备,使生产中不产生或少产生有害物质。
②综合利用,从废水中回收有用物质,废水中的污染物,往往是宝贵的工业原料或产品、半成品流失物,若予回收利用,便可以变废为宝,化害为利。
③加强管理,控制水污染必须有完善的管理措施,包括立法管理、资源管理和规划管理。
④无害化处理,达标排放,对外排的废水应妥善处理,做到无害化排放,这是防止环境污染的重要措施。
5、废水处理的方法:废水处理的方法很多,按其作用原理可分为物理法、化学法、物理化学法、生物法四种。
第一章物理处理法6、调节池的作用有哪些?①减少或防止冲击负荷对处理设备的不利影响。
②使酸碱废水中和、PH值保持稳定。
③调节水温④当设备发生故障时起临时储水作用。
7、简述沉淀的作用和沉淀的类型?是使水中的悬浮物在重力作用下下沉,从而与水分离,使水质得到澄清。
沉淀共分四个类型,即自由沉淀、絮凝沉淀,拥培沉淀和压缩沉淀。
8、沉砂池的功能和主要类型?沉砂池的功能是从废水中分离较大的无机颗粒,如砂、炉灰渣等。
它一般设在泵站、沉淀池之前,用于保护机件和管到免受磨损。
沉沙池的类型,按池内水流方向的不同,可分为平流式和竖流式。
常用的有平流式沉砂池曝气沉沙池。
9、沉淀池有哪几种主要类型?平流式沉淀池、辐流式沉淀池、竖流式沉淀池和斜板(管)沉淀池。
第七章废水生物化学处理基础本章重点:如何建立单个细菌以及生物膜或生物絮体的数学模型。
1947年,首次出现了“生物化学工程”( Biochemical engineering)一词。
1965年Aiba等人的专著《物化学工程》(Biochemical Engineering)出版,标志着这一学科的正式出现。
1971年Coulson及Richardson等著述的化学工程标准教材新添了第三卷,其中包括了一章生物化学反应工程,标志着生物化学工程已成为化学工程的—个新的组成部分。
此后出版的生物化学工程专著有Atkinson的《生物化学反应器》(Biochemical Reactors,1974年),Bailey及ollis 的《生物化学工程基础》(Biochemical Engineering Fundamentals.1977年)等书。
生物化学工程中应用的发酵器有两种基本类型,一种是利用微生物絮体的作用,这与废水处理中的活性污泥法相类似;另一种是利用微生物膜的作用,这与废水处理中的生物滤池法相类似。
以生物化学工程的方法来研究废水的生物处理,提高了它的理论深度,应该是发展的方向。
把废水的生化处理看成是生物化学工程的一个重要分支,在学科体系上可能更合适—些。
§7.1 单个细菌的模型从细菌结构及代谢途径来看,如果要按实际情况建立一个数学模型,几乎无法着手。
所以目前一般采用一个远为简化的模型,而这个模型也起到了对营养物传入细菌内的整个过程,给出明确概念的作用。
底物一般是通过细胞的粘液层、细胞壁与细胞膜进入细胞内部的,而代谢作用只发生在细胞内部的细胞质区。
发生代谢作用后,底物也就消失了。
这里,我们假设:①不考虑复杂的代谢过程;②把底物的消失引用流体力学中“汇”的概念来解释;③粘液层、细胞壁、细胞膜等作为底物传递的边界。
这样就得到一个细菌的简化模型,如图7-1所示。
扩散区指细胞壁外粘液层的部分,其表面积为a d cm 2,,底物通过扩散区时服从Fick 的第一扩散定律,即底物的通量为:Nd = -Dγρd d (7-1) 式中,下标d 表示扩散区,γρd d 表示晏半径γ方向的浓度梯度,D 仍然表示分子扩散系数。
扩散区的内面为透酶区。
这一区指细胞膜的透酶所起的运输作用。
透酶是细脑膜内的一类立体专一性载体分子,这类分子也是一种蛋白质,取名透酶以示区别于代谢酶。
透酶区的通量可用下列公式来表示:'P 'p P K a N ρ+ρ= (7-2)式中的下标p 表示透酶区,a p 及Kp 为两个常数,ρ’为透酶区外的底物浓度。
通量Np 只与透酶区外的底物浓度ρ’有关,而与代谢区中的底物浓度ρ’’无关。
当ρ’> ρ’ 时,称为被动运输;ρ’< ρ’时,称为主动运输。
代谢区指细胞膜内的区域。
这一区域内虽然产生了许多极复杂的代谢途径,但组成代谢途径的每一个反应都是由酶控制的,因而服从于Michaelis —Menten 方程。
代谢区内底物消耗速率可以表示为:''m ''m ''K a dt d ρ+ρ=ρ (7-3) 式中,ρ’’表示代谢区中底物的浓度,a m 及K m 为Michaelis-Menten 方程的常数。
当代谢区消耗底物的速率恰好和底物通过两个运输区的速率相等时,便得到一个稳定的状态,这时存在下列关系:⎪⎪⎭⎫ ⎝⎛ρ+ρ=⎪⎪⎭⎫ ⎝⎛ρ+ρ=⎪⎪⎭⎫ ⎝⎛-γρ''m ''m m 'p 'p p r d K a V K a a d d D a d (7-4) 式中,a d 为扩散区的外表面积,下标r d 指浓度d ρ/d γ计值的扩散外径,a p 为透酶区的外表面积,V m 为代谢区的容积。
当底物不需透酶区的运输时,式(7-4)简化为:m ''p m ''p m ''m ''m m r d a )K (a a V K a V d d D a d ⎪⎪⎪⎭⎫ ⎝⎛ρ+ρ=⎪⎪⎭⎫ ⎝⎛ρ+ρ=⎪⎪⎭⎫ ⎝⎛-γρ (7-5) 当包含透酶区时,由式(7-4)看出底物的消耗速率完全由运输过程来控制,即由下列关系控制:⎪⎪⎭⎫ ⎝⎛ρ+ρ=⎪⎪⎭⎫ ⎝⎛-γρ'p 'p p r d K a a d d D a d (7-6) 表达不需要透酶运输的式(7-5)和需要透酶运输的式(7-6)可以共用下列公式来表示:⎪⎪⎭⎫ ⎝⎛ρ+αρ=⎪⎪⎭⎫ ⎝⎛-γρK a d d D a d r d (7-7) 式中,a 代表底物所通过的表面积;α及K 为常数,α代表式(7-5)的V m αm /αm 或式(7-6)的αp ;ρ为相应的浓度ρ’或ρ’’。
§7-2 细菌的连续增殖连续培养器有多种形式,有的结构很复杂,但概括起来只分两类,一类叫恒化器(chemostat),另一类叫恒浊器(turbidostat)。
恒化器控制培养液中某一限制营养物的浓度为恒定值,从而控制了细菌的增殖率,是一种间接的控制。
恒浊器靠控制培养物溢流的浊度(代表细菌浓度)为恒定值来控制细菌的增殖率,足一种直接的控制。
简单的恒化器见图7—3,是一个工作容积可以小至100mL 的容器。
进入恒化器的灭菌培养液的流量为f mL/h ,恒化器的溢流流量也是f rnL/h ,恒化器内液体容积为V .并不断供给灭菌空气,以保证细菌的需氧过程。
培养液处在不断搅拌过程巾,以保证培养液的成分均匀。
就整个体系而言,当达到每秒钟增加的细菌个数与每秒钟排掉的细菌个数相等时,恒化器即处于稳定状态。
图7—3所示的恒化器实际可看作是一个CSTR 。
每小时通过溢流量f 所排掉的细菌质量为:f × 1mL 中的细菌质量 = f × Vx = Dx 式中,x/V 代表恒化器1mL 液体中所含细菌的质量,也是1mL 溢流流体中所含细菌的质量;D 代表f/V ,为新鲜培养液在容积V 中的稀释率,量纲为时间-1。
由于细菌的增殖率可表示为dx/dt=µx ,所以当恒化器处于稳定条件下时得:Dx x dtdx =μ= 在恒化器中,Monod 方程可写为:ρ+ρμ==μs max K D (7-9)由图7-5可知,当生物处理设备的进水有机物浓度在一定范围内波动时不会引起微生物特性很大的变化,因而系统的运行能处于稳定状态。
Monod 方程中µmax 和K s 值取决于所采用的细菌和营养物类别。
根据Monod 方程,即式(7-9),可以求得恒化器稳态条件的营养物浓度ρ为:DD K max s -μ=ρ (7-10) §7-3 细菌增殖速率与底物消耗速率关系式把底物的消耗速率分成两部分.一部分是由于细菌生长新的细胞物质而产生的,以生长⎪⎭⎫ ⎝⎛ρdt d 表示,另一部分是为维持细菌处于活的状态所需的能量而产生的,以维持⎪⎪⎭⎫ ⎝⎛ρdt d 表示,这就得:总⎪⎭⎫ ⎝⎛ρdt d =生长⎪⎭⎫ ⎝⎛ρdt d + 维持⎪⎪⎭⎫ ⎝⎛ρdt d (7-13) 推导:由式(6-22)可得:dt dx Y 1dt d -=⎪⎭⎫ ⎝⎛ρ总 dt dx Y 1dt d c-=⎪⎭⎫ ⎝⎛ρ生长 式中:Y c 称为真产率因数。
维持能量所需的消耗速率维持⎪⎭⎫ ⎝⎛ρdt d 应该与细菌的质量x 成正比,可以表示为:mx dt d -=⎪⎭⎫ ⎝⎛ρ维持式中,m 称为维持系数,量纲为时间-1。
这样(7-13)可以写成:CY 1m Y 1+μ= (7-14) §7-4 BOD 与TbOD1. 生化需氧量(BOD )与BOD 试验水中有机物通过微生物的氧化变成简单无机化合物的过程中,对水中溶解氧的消耗速率,称为它的生化需氧量。
这里的微生物主要指细菌。
细菌以有机物为食物而生长,在生长过程中,一部分有机构转化成为新的细菌细胞,同时产生二氧化碳和水等。
当水中食物不足时,细菌又从本身物质中吸取能量以维持生命.这一现象称为内源代谢(endogenous metabalism)或内源呼吸(endogenous respiration)。
细菌死后.又以有机物的形式作为细菌的食物而重复上述过程。
另外,活的细菌与死的细菌又是原生动物和其它较高级微生物的食物,原生动物这类微生物因此称为捕食微生物。
图7—7给出了新鲜生活废水的生化需氧量历时曲线形式和温度对历时曲线的影响。
第一阶段:由于含碳有机物的分解所需要的生化需氧量,也称碳质BOD (carbonaceous BOD );第二阶段:(硝化阶段)代表含氮有机物硝化过程的需氧量,称为氮质BOD(nitrogenousBOD)。
当典型的碳源物质葡萄糖完全氧化时可以写成:O H 6CO 6O 6O H C 2226126+→+ (7-17)则可认为生化需氧量等于2.67×有机物碳原子的质量浓度。
细菌细胞的合成可以写成:O H 110CO 59N O H C 17NH 17O 59O H C 2422275326126++=++ (7-18) 由此可计算,每合成1g 干细菌,约需单体氧0.985g 。
细菌的氧化分解可以写成:3222275NH O H 2CO 5O 5NO H C ++=+ (7-19)按这一反应计算,每克干细菌的完全氧化约需单体氧1.42g 。
含碳有机物完全氧化成二氧化碳及水的生化需氧量称为总生化需氧量,以BOD L 或BOD u 表示。
硝化BOD 的反应可表示为:O H 2H 2NO 2O 3NH 22223++→++- (7-20)+-+-+→++H 2NO 2H 2O NO 2322 (7-21)按这两个反应,可得:硝化BOD = 4.57(有机氮 + 氨氮) mg/L + 1.14 (NO 2- 氮)mg/L (7-22)这里特别指出:上述生化需氧量概念为其原始涵义,与生化需氧量试验所测得的生化需氧量完全不是一个同一概念。
这可以从图7-8中看出,接种细菌的生长过程中有一个滞后期,在这一段时间内.细菌的浓度没有变化,接种的细菌在滞后期中虽然也要摄取一定食物及溶解氧.但是量甚少,所以有机物浓度可以视作无变化。
BOD值应为零,只是当细菌开始增殖后,有机物浓度才开始下降,当细菌浓度达最大值时,有机物浓度也降为零。
在有机物浓度为零以后.细菌靠内源呼吸以及死的细菌以取得营养物。
在这一阶段,由于有足够的细菌为食料,原生动物也开始增殖起来。
细茵的内源呼吸以及原生动物的生长代谢都同时摄人氧。
包括在这一阶段的BOD值中。
图7-8反映了BOD试验所存在的两方面的问题:①BOD5与总BOD值不会具有一定的数量关系。