第十二章细胞周期和细胞分裂
- 格式:doc
- 大小:29.00 KB
- 文档页数:6

细胞生物学中的细胞周期和细胞分裂细胞生物学是研究细胞的结构、功能和生命过程的科学。
细胞周期和细胞分裂是细胞生物学中非常重要的概念,它们直接关系到细胞的增殖和遗传信息的传递。
本文将从细胞周期和细胞分裂的定义、细胞周期的阶段以及细胞分裂的过程进行详细阐述。
一、细胞周期和细胞分裂的定义细胞周期是指从一次细胞分裂开始,到下一次细胞分裂开始的整个过程。
细胞周期可以分为四个连续的阶段:G1期(细胞生长期)、S 期(DNA合成期)、G2期(前期)和M期(有丝分裂期)。
其中,G1、S、G2三个阶段合称为间期。
细胞分裂是指细胞通过复制染色体并均等分配到两个新的细胞中,从而使一个细胞分裂成为两个细胞的过程。
细胞分裂主要分为两种类型:有丝分裂和无丝分裂。
有丝分裂是大多数真核细胞的分裂方式,而无丝分裂主要发生在原核生物和有些真核生物的有特殊要求的细胞中。
二、细胞周期的阶段1. G1期(细胞生长期)G1期是细胞周期中最长的一个阶段,它通常占据整个周期的一半甚至更长的时间。
在G1期,细胞会进行各种生化代谢活动,例如合成蛋白质和增加细胞器的数量。
在这个阶段,细胞还会接受外界信号,判断是否具备进行DNA复制和细胞分裂的条件。
2. S期(DNA合成期)在S期,细胞会进行DNA的复制,这是细胞周期中至关重要的一个阶段。
DNA的复制过程是通过酶的作用,在细胞核内顺次复制每一个染色体。
这样,每个染色体会变成由两条完全相同的复制体组成的染色体。
3. G2期(前期)G2期是DNA复制完成后距离细胞分裂的前期。
在这一阶段,细胞会进行所必需的准备工作,例如合成蛋白质和其他细胞器的增殖。
细胞会通过检查自身是否具备正常状态来保证细胞分裂的成功进行。
4. M期(有丝分裂期)M期是细胞周期中用于有丝分裂的阶段。
有丝分裂是细胞分裂的一种重要方式,它包括核分裂(核分裂前期、核分裂中期和核分裂后期)和细胞质分裂。
在核分裂前期,细胞核会逐渐发育成具有两个核仁的核。

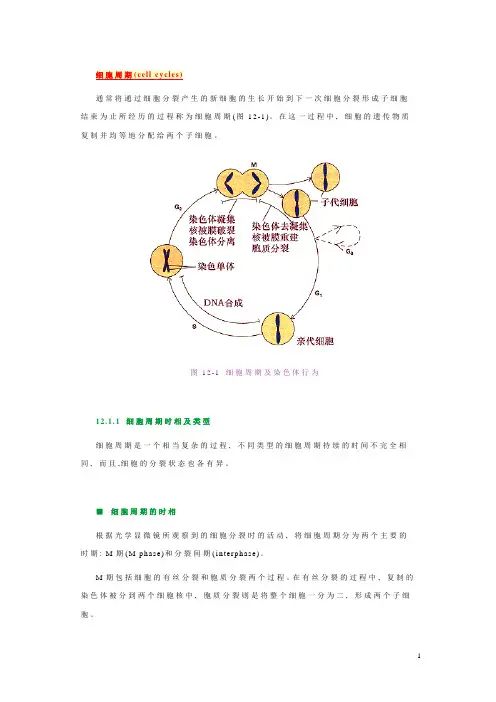
通常将通过细胞分裂产生的新细胞的生长开始到下一次细胞分裂形成子细胞结束为止所经历的过程称为细胞周期(图12-1)。
在这一过程中,细胞的遗传物质复制并均等地分配给两个子细胞。
图12-1细胞周期及染色体行为12.1.1细胞周期时相及类型细胞周期是一个相当复杂的过程,不同类型的细胞周期持续的时间不完全相同,而且,细胞的分裂状态也各有异。
■细胞周期的时相根据光学显微镜所观察到的细胞分裂时的活动,将细胞周期分为两个主要的时期:M期(M p h a s e)和分裂间期(i n t e r p h a s e)。
M期包括细胞的有丝分裂和胞质分裂两个过程。
在有丝分裂的过程中,复制的染色体被分到两个细胞核中,胞质分裂则是将整个细胞一分为二,形成两个子细胞。
分裂间期实际上是新细胞的生长期,根据新细胞从开始生长起到分裂前止的分裂间隔期中的生理和生化变化,可分为: G1期(Ga p1 p h a s e),即从M期结束到S 期开始前的一段间歇期;S期,即D N A合成期(D N A s yn t h e t i c p h a s e);G2期(G2p h a s e),即D N A合成后(S期)到有丝分裂前的一个间歇期。
是否所有生物的细胞周期持续的时间都相同?主要差别在哪里?(答案)答:不同生物的细胞周期时间不同,同一系统中不同细胞,其细胞周期的时间也有很大的差异。
细胞周期所持续的时间一般为12~32小时,M期所持续的时间较短,一般为30~60分钟,分裂间期的时间跨度较长,根据细胞的类型和所处的生理条件不同而不同,有几小时、几天、几周或更长。
如人的细胞周期约为24小时∶丝裂期30分钟,G1期9小时,S期10小时,G2期 4.5小时。
一般说来,S+G2+M的时间变化较小,主要差别在G1期的长短。
如消化系统,小鼠食管和十二指肠上皮细胞,它们的细胞周期时间,分别为115小时和15小时,食管上皮细胞的G1期长达103)不同生物的细胞周期时间不同,同一系统中不同细胞,其细胞周期的时间也有很大的差异。





第十三章细胞分裂与细胞周期一、名词解释1、细胞周期2、细胞周期检验点3、有丝分裂4、减数分裂5、有丝分裂器6、(细胞促分裂因子)7、细胞周期蛋白8、终端分化细胞二、填空题1、在细胞有丝分裂中, 微管的作用是、;微丝的作用是。
2、中心粒是由构成的,每个中心体各含有一对互相的中心粒,在细胞周期的期进行复制。
3、动物细胞的有丝分裂器有、、和四种类型的微管;植物细胞中没有。
4、细胞分裂的方式有、和。
5、细胞周期可分为四个时期即、、、和。
6、按照细胞增殖能力不同,可将细胞分为三类、和。
7、在细胞周期调控中,调控细胞越过G1期限制点的与周期蛋白的复合物称为。
8、由两个亚单位组成,即和。
当两者结合后表现出蛋白激酶活性,其中为催化亚单位,为调节亚单位。
9、肝细胞和肌细胞属于不同细胞周期类型,肝细胞在受到损伤情况下能进行分裂,而肌细胞却不行,由此可判断肝细胞属于,而肌细胞属于。
10、细胞周期中重要的检验点包括、、和。
11、在减数分裂的前期发生同源染色体的和等位基因的;在有丝分裂后期中,是发生分离,而在减数分裂后期I中则是发生分离。
三、选择题1、若在显微镜下观察到的某细胞具有核仁, 并且核物质与细胞质的界限清晰, 则可判定此细胞处于细胞的( )。
A、间期B、前期C、中期D、后期2、在细胞分裂中期与纺锤体的动粒微管相连,保证染色体平均分配到两个子细胞中的结构是()。
A、复制源B、着丝粒C、端粒D、动粒3、关于细胞周期限制点的表述,错误的是()。
A、限制点对正常细胞周期运转并不是必需的B、它的作用是细胞遇到环境压力或受到损伤时使细胞周期停止的"刹车"作用,对细胞进入下一期之前进行“检查”。
C、细胞周期有四个限制点:G1、2、G2和G1限制点D、最重要的是G1限制点4、的分子组成是()。
A、2和B、1和C、4和D、2和5、细胞周期正确的顺序是()。
A、G12B、G12C、G12D、G126、在减数分裂过程中,同源染色体进行交叉和互换的这个时期称为()。

第十三章细胞分裂与细胞周期一、名词解释1、细胞周期2、细胞周期检验点3、有丝分裂4、减数分裂5、有丝分裂器6、MPF(细胞促分裂因子)7、细胞周期蛋白8、终端分化细胞二、填空题1、在细胞有丝分裂中, 微管的作用是、;微丝的作用是。
2、中心粒是由_________构成的,每个中心体各含有一对互相__________的中心粒,在细胞周期的______________期进行复制。
3、动物细胞的有丝分裂器有、、和四种类型的微管;植物细胞中没有。
4、细胞分裂的方式有、和。
5、细胞周期可分为四个时期即、、、和。
6、按照细胞增殖能力不同,可将细胞分为三类、和。
7、在细胞周期调控中,调控细胞越过G1/S期限制点的CDK与周期蛋白的复合物称为。
8、MPF由两个亚单位组成,即和。
当两者结合后表现出蛋白激酶活性,其中为催化亚单位,为调节亚单位。
9、肝细胞和肌细胞属于不同细胞周期类型,肝细胞在受到损伤情况下能进行分裂,而肌细胞却不行,由此可判断肝细胞属于,而肌细胞属于。
10、细胞周期中重要的检验点包括、、和。
11、在减数分裂的前期发生同源染色体的和等位基因的;在有丝分裂后期中,是发生分离,而在减数分裂后期I中则是发生分离。
三、选择题1、若在显微镜下观察到的某细胞具有核仁, 并且核物质与细胞质的界限清晰, 则可判定此细胞处于细胞的( )。
A、间期B、前期C、中期D、后期2、在细胞分裂中期与纺锤体的动粒微管相连,保证染色体平均分配到两个子细胞中的结构是()。
A、复制源B、着丝粒C、端粒D、动粒3、关于细胞周期限制点的表述,错误的是()。
A、限制点对正常细胞周期运转并不是必需的B、它的作用是细胞遇到环境压力或DNA受到损伤时使细胞周期停止的"刹车"作用,对细胞进入下一期之前进行“检查”。
C、细胞周期有四个限制点:G1/S、S/G2、G2/M和M/ G1限制点D、最重要的是G1/S限制点4、MPF 的分子组成是()。
细胞周期与细胞分裂细胞是生命的基本单位,无论是单细胞生物还是多细胞生物,其生命周期都与细胞周期密切相关。
细胞周期是指从细胞分裂开始到再次分裂结束的一系列连续发展过程,它分为四个阶段:G1期(第一生长期)、S期(DNA复制期)、G2期(第二生长期)和M期(有丝分裂期)。
在G1期,细胞进行生长和代谢活动,准备进行DNA复制。
G1期是细胞周期中最长的一个阶段,其时间长短因细胞类型不同而不同。
在生长因子的刺激下,细胞会进一步进入S期。
S期是细胞周期的重要阶段,也是DNA复制的阶段。
在这个阶段,细胞将其染色体的DNA复制为两倍,并保留在细胞核中。
这一过程确保每个新细胞都包含与母细胞完全相同的遗传信息。
G2期是G1期和M期之间的一个重要过渡阶段,它标志着DNA复制的完成,细胞开始准备进行有丝分裂。
在这个阶段,细胞核中的染色体变得更加紧密,并准备分离为两个子细胞。
M期是细胞周期中最为重要的阶段,也是细胞分裂的阶段。
M期可以分为核分裂和细胞质分裂两个步骤。
核分裂又分为促进期、分裂期和结束期。
在核分裂的过程中,细胞核中的染色体分别缩短并形成以两个相同的染色体为一对的染色体。
同时,纺锤体在细胞的两端形成,它们的纤维束将染色体吸引过来分离。
细胞质分裂发生在核分裂之后,它是细胞周期中的最后一个阶段。
在细胞质分裂的过程中,细胞中的胞质分离为两个子细胞。
子细胞分离后,细胞周期又重新开始,进入新的G1期。
细胞周期的调控对于维持细胞正常的生长和遗传稳定性至关重要。
细胞周期调控通过一系列复杂的信号传导通路和调节蛋白分子来实现。
其中,细胞周期素和Cyclin便是两个关键调节蛋白。
细胞周期素与Cyclin通过结合形成活化复合体,从而促进细胞向下一个阶段的转变。
除了细胞周期的正常进行,细胞分裂也是维持生物体生长和发育的重要过程。
细胞分裂分为两种类型:有丝分裂和无丝分裂。
有丝分裂是较为常见和典型的细胞分裂方式,包括前面提到的M期的步骤。
第十二章细胞周期和细胞分裂1. 是否所有生物的细胞周期持续的时间都相同?主要差别在哪里?答: 不同生物的细胞周期时间不同, 同一系统中不同细胞,其细胞周期的时间也有很大的差异。
细胞周期所持续的时间一般为12~32小时, M期所持续的时间较短, 一般为30~60分钟, 分裂间期的时间跨度较长, 根据细胞的类型和所处的生理条件不同而不同, 有几小时、几天、几周或更长。
如人的细胞周期约为24小时∶丝裂期30分钟,G1期9小时,S期10小时,G2期4.5小时。
一般说来, S+G2+M的时间变化较小, 主要差别在G1期的长短。
如消化系统, 小鼠食管和十二指肠上皮细胞, 它们的细胞周期时间, 分别为115小时和15小时, 食管上皮细胞的G1期长达103小时, 而十二指肠上皮细胞的G1期为6小时。
2. 根据细胞分裂行为,可将细胞分为几种类型?各有什么特点?答: 根据细胞的分裂行为, 可将真核生物细胞分为三类:①持续分裂细胞,又称周期性细胞, 即在细胞周期中连续运转的细胞。
机体内某些组织需要不断的更新,组成这些组织的细胞就必须通过不断分裂产生新细胞。
此类细胞的分裂周期非常正常, 有丝分裂的活性很高。
如性细胞(包括卵母细胞和精原细胞),它们要不断地产生配子; 造血干细胞需要不断地产生红细胞和白细胞;上皮基底层细胞需要通过分裂不断补充表面老化死亡的细胞; 植物的根茎尖端细胞需要通过分裂进行生长等都是具有正常周期的持续分裂细胞。
②终端分化细胞, 即永久性失去了分裂能力的细胞,它们不可逆地脱离了细胞周期, 但保持了生理活性机能。
这些细胞都是高度特化的细胞, 如哺乳动物的红细胞、神经细胞、多形性白细胞、肌细胞等, 这些细胞一旦分化,就永远保持这种不分裂状态直到死亡。
③G0细胞,又称休眠细胞,暂时脱离细胞周期,不进行DNA复制和分裂, 也称静止细胞群。
但这些细胞可在某些条件的诱导下重新开始DNA合成, 进行细胞分裂。
如肝细胞, 外科手术切除部分肝组成后可以诱导进入细胞分裂。
淋巴细胞可通过与抗原的相互作用诱导增殖。
在胚胎发育早期(卵裂期),所有的细胞均为周期性细胞, 以后随着发育成熟, 某些细胞进入了GO期, 某些细胞分化后丧失分裂能力。
到成体时,只有少数细胞处于增殖状态, 它们的增殖仅作为补充丢失的细胞, 或对外界刺激的反应。
3. 根据细胞周期各时相的生化活动,推测细胞的表面形态和内部结构各有哪些变化?答: 由于细胞周期的各时相的生化活动不同,引起不同的表面和内部结构的变化:细胞形态的变化:如处于S期的细胞呈扁平状, 紧贴在培养瓶壁上, 细胞表面的微绒毛和小泡很少。
细胞进入G2期, 特别是G2期的中后期, 细胞渐渐从贴壁摊平的状态鼓起来, 而细胞表面的微绒毛增多, 此时摇动培养瓶, 细胞很容易与瓶壁脱离。
进入M期的细胞, 变成球形。
细胞内部结构的变化内部结构的最大变化是染色质结构的变化。
在S期, 染色体处于极松散的状态, DNA半保留复制和核小体八聚体组蛋白全保留方式偶联。
到G2期已形成两条染色质纤维; 到M期, 染色单体形成。
与染色体复制周期相关联的是核仁的变化。
从细胞分裂前期到M期中期, 核仁消失, 核膜解体;分裂后期重新形成核膜。
在间期-分裂期过渡中, 有两点明显的变化: 一是形成纺锤体, 需要大量的微管蛋白,另一是细胞表面微绒毛的形成,这与细胞骨架的肌动蛋白纤维相关。
4. 美国科学家利兰?哈特韦尔和英国科学家蒂莫西?亨特、保罗?纳西分享了2001年的生理学会医学诺贝尔奖,他们各自的贡献是什么?答: 利兰?哈特韦尔发现了“START”基因;保罗?纳西的贡献是发现了CDK。
蒂莫西?亨特的贡献是发现了调节CDK的功能物质CYCLIN。
5. 遍在蛋白如何介导周期蛋白的降解?答:遍在蛋白加到周期蛋白上需要三种不同的酶介导。
首先遍在蛋白在它的羧基端通过与遍在蛋白激活酶E1的半胱氨酸残基形成硫酯键而激活。
然后遍在蛋白从E1转移到E2的半胱氨酸残基, E2称作遍在蛋白结合酶(ubiquitin)。
E2和第三种酶, 遍在蛋白连接酶(ubiquitinligase, E3)一起将遍在蛋白转移到底物蛋白的赖氨酸残基(共价结合), 在那里进行遍在蛋白的聚合化,最后作为蛋白酶体的降解底物, 被快速降解。
E3通常是一种复合体,由多亚基组成。
例如从非洲爪蟾卵细胞中分离的周期蛋白B的E3至少含有8个不同的亚基。
触发有丝分裂周期蛋白遍在蛋白聚合化的E3又称为后期促进复合物(anaphase-promoting complex,APC)。
APC激发E2-遍在蛋白复合物同有丝分裂周期蛋白破坏框结合, 然后激发遍在蛋白同破坏框C-末端的赖氨酸残基结合,此过程不断循环使遍在蛋白聚合化。
通过基因操作构建了不含破坏框的周期蛋白, 这些蛋白不会被降解。
6. APC的活性调节及在周期蛋白B降解中的作用如何?答: 当MPF的活性在有丝分裂中期达到最高峰时, 它将APC磷酸化并将其激活。
被激活的APC与E2结合, 最后结合到周期蛋白B的破坏框中, 促使周期蛋白B的进行遍在蛋白多聚化, 其结果导致周期蛋白B降解。
由于周期蛋白B是MPF的一个必需亚基, 它的降解势必导致MPF活性降低甚至失活, 触发细胞进入有丝分裂末期。
胞质分裂之后, 周期蛋白B在子细胞的间期合成, APC的活性保持到G1期的后期, 被G1 Cdk失活。
周期蛋白B的浓度不断升高,同时提高MPF的活性, 以便进入下一个有丝分裂期。
7. 真核细胞周期调控模型的主要特点和机制是什么?答: 特点表现在三类周期蛋白-Cdk复合物和三个关键的过渡和对细胞周期的控制。
细胞周期中三个关键的过渡:细胞周期中三个关键的过渡,即G1期→S期、中期→后期、后期→末期及胞质分裂期是细胞周期中三个关键过渡。
这三个过渡分别被Cdc34途径和APC途径控制。
三类周期蛋白-Cdk复合物:真核细胞主要通过三类周期蛋白-Cdk复合物的作用,控制细胞周期, 这三种复合物分别是:G1期、S期和有丝分裂Cdk复合物。
这些复合物都是由周期蛋白依赖性的蛋白激酶和周期蛋白两个亚基组成的复合物。
在Cdc34途径和APC途径中,蛋白复合物都是通过遍在蛋白的蛋白酶体将一些特殊的底物, 包括S期抑制物、后期抑制物、以及有丝分裂周期蛋白降解进行周期调节。
细胞周期中三个关键的过渡:细胞周期中三个关键的过渡,即G1期→S期、中期→后期、后期→末期及胞质分裂期的过渡。
这些过渡都是通过触发蛋白质的降解进行的, 所以都是不可逆转的, 这样迫使细胞周期只能沿一个方向进行。
三个过渡分别是通过Cdc34和APC途径的降解作用完成的。
Cdc34途径促使细胞从G1→S期过渡:在G1期的早、中期,在DNA的复制起点就装配了复制起始复合物,并且开始了S期的CdkC组份的转录;但是,S期CdkC抑制物被G1 CdkC磷酸化而激活,激活的S期CdkC抑制物抑制了S期CdkC的活性,从而使细胞停留在G1期。
在G1期的后期, Cdc34诱导S期Cdk抑制物的降解, 释放出有活性的S期Cdk复合物,这种复合物能够激发细胞进入S期。
一旦S期Cdk的降解作用被激活, S-期Cdk复合物将与DNA形成预复制复合物中的蛋白质的调节位点磷酸化,预复制起始复合物是G1期在DNA复制起点上装配的复合物。
这些被S-期Cdk复合物磷酸化的蛋白质不仅能够激活DNA复制起始, 还能够阻止新的预复制复合物的装配。
由于这种抑制作用, 每条染色体在细胞周期中只复制一次, 保证了合适的分配到子细胞中的染色体数。
有丝分裂Cdk复合物是在S期和G2期合成的, 但是它们的活性一直受到抑制直到DNA合成完毕。
一旦被激活, 有丝分裂Cdk复合物就会诱导染色体凝聚、核膜解体、有丝分裂器的装配以及凝聚的染色体在中期赤道板上排列。
在所有凝聚的染色体都与适当的纺锤体微管结合之后,有丝分裂Cdk复合物激活后期启动复合物(anaphase promoting complex, APC)。
这种多蛋白的复合物指导后期抑制物通过遍在蛋白介导的蛋白酶解作用进行降解, 导致在中期将姐妹染色单体结合在一起的蛋白复合物失活。
因此这些抑制物的降解作用, 允许有丝分裂进入到后期。
在后期末, APC也可指导有丝分裂周期蛋白的蛋白酶体的降解。
有丝分裂Cdk活性的降低, 使得分离的姐妹染色单体去凝聚、核膜重新形成、在胞质分裂中细胞质的分裂, 最后形成子细胞。
8. 裂殖酵母的MPF的化学本质是什么?是如何发现的?答: 裂殖酵母油MPF是一种复合物,由Cdc2和Cdc13两种蛋白组成,其中Cdc2是一种蛋白激酶,Cdc13是周期蛋白。
主要是通过对裂殖酵母温度敏感突变的研究发现了编码这两种蛋白的基因。
裂殖酵母中几个温度敏感的cdc基因的隐性突变使得裂殖酵母在周期中不能进入M期,由于生长没有停止, 所以比正常的酵母长很多。
这些基因中的一个显性突变, 命名为cdc2,使得酵母出现wee表型(小而不分裂)。
一般来说, 隐形表型是因为缺少野生型的功能蛋白所致, 而显性表型是因为增加了一些蛋白功能,导致调节失常。
对分离的突变体的研究发现, 没有Cdc2的活性,细胞不能进入有丝分裂。
这表明Cdc2是裂殖酵母进入有丝分裂的一个关键调节因子。
通过基因工程克隆到cdc2-基因, 序列分析表明该基因编码一个分子量为34kDa的蛋白, 与真核生物的蛋白激酶同源, 该蛋白又称为p34cdc2 蛋白。
后来从裂殖酵母中分离了另一个cdc基因, 命名为cdc13+,该基因的产物也是裂殖酵母进入有丝分裂必需的;序列分析表明该基因编码的蛋白质与非洲爪蟾和海胆的周期蛋白B同源。
进一步研究发现Cdc13和Cdc2蛋白能够形成异质二聚体, 并且具有蛋白激酶的活性。
另外还发现这种蛋白激酶的活性随裂殖酵母的周期变化而变化, Cdc13的浓度也随Cdc2的活性波动而波动。
这些结构表明, 裂殖酵母的Cdc2-Cdc13相当于非洲爪蟾的MPF。
9. 裂殖酵母中MPF活性的活性是如何调节的?答: 裂殖酵母MPF的活性调节是相当复杂的, 涉及多种蛋白激酶以及Cdc2亚基上两个位点的磷酸化与去磷酸化。
p34cdc2蛋白单亚基上有两个磷酸化的位点, 一个是激活型的磷酸化位点, 另一个是抑制型的磷酸化位点。
独立存在的p34cdc2 蛋白激酶是无活性的, 同周期蛋白Cdc13结合后,仍然没有活性, 但此时的复合物成为两种蛋白激酶的底物, 一种是Weel激酶, 它使p34cdc2亚基上的抑制位点Tyr-15位残基磷酸化, 抑制MPF的活性。
第二种蛋白激酶是Cdc2激活蛋白激酶(Cdc2-activating kinase, CAK), 可以使Cdc2亚基中激活型的位点Thr-161(T-161)残基磷酸化, 这种磷酸化最大限度地激活了MPF的活性, 但是, 只要Tyr-15(Y-15)位残基是磷酸化的,Cdc2-周期蛋白复合物就没有活性。