细胞超微结构(1)
- 格式:ppt
- 大小:5.27 MB
- 文档页数:88







一)超微结构:(电镜下)1)叶绿体由叶绿体膜(2层单位膜)、类囊体、基质(stroma)三部分构成2)结构特点;三种不同膜(外膜、内膜和类囊体膜)三种彼此分隔空隙(膜间隙、基质、类囊体腔)3)叶绿体膜(Chloroplast membrane) 或称外被(outer velope)两层单位膜组成,6~8nm/层(膜厚度)①外膜:通透性大,能通透:无机盐、核苷、蔗糖等多种物质;细胞质和膜间隙的功能屏障;②内膜:选择性通透,上分布有专门的转运载体,负责转运小分子;隔开膜间隙和基质其。
③膜间隙(intermembrane space):10~20nm4)类囊体(thylakoid)①叶绿体基质中由单位膜封闭形成的扁平小囊—类囊体;(基粒类囊体和基质类囊体)②叶绿体内部组织的基本结构和功能单位(其上分布许多光合作用色素,是光合作用的光反应场所);③类囊体结构特点:一般沿叶绿体长轴平行排列,某些部位园盘状的基粒类囊体堆积成柱形颗粒—基粒(grana);连接相邻基粒—基质类囊体;基质类囊体使叶绿体内类囊体成为一完整、连续的封闭的膜囊。
④基粒类囊体(granum thylakoid)直径0.25~0.8μm,厚0.01μm5~30个基粒类囊体/基粒40~80个基粒/叶绿体中基质类囊体(stroma thylakoid);A)贯穿在两个或以上没有发生重叠的类囊体,B)由基粒类囊体延伸出来的网状或片状结构;类囊体膜特点—光合膜:A)膜结构不对称;B)膜上分布不同的参与光和作用的色素复合体(如叶绿素蛋白复合体),酶和电子传递载体蛋白及细胞色素。
5)叶绿体基质(stroma)①存在位置:内膜与类囊体之间无定形物质,为叶绿体基质;②基质的主要成分:A)可溶性蛋白,RuBP羧化酶(1,5—二磷酸核酮糖羧化酶)占可溶性蛋白的60%;B)DNA和RNA(rRNA)。
6)类囊体腔——储氢池(H+—池)二)叶绿体功能—光合作用(photosynthesis):1)光合作用意义——能量源泉自然界将光能转化为化学能主要途径,地球上植物每年捕获太阳能,至少可转化产生4.2×1017焦耳自由能。

细胞超微结构细胞超微结构概述Virchow在19世纪中期所奠定的细胞病理学说,通过近代对细胞及其病变的超微结构以及结构与功能相结合的研究,已经获得了新的更广更深的基础,扩大和加深了对疾病的理解. 细胞是一个由细胞膜封闭的基本生命单元,内含一系列明确无误的互相分隔的反应腔室,这就是以细胞膜为界限的各种细胞器,是细胞代谢和细胞活力的形态支柱. 细胞内的这种严格分隔保证各种细胞器分别进行着无数的生化反应,行使各自的独特功能,维持细胞和机体的生命活动.细胞器的改变是各种病变的基本组成部分.一、细胞核细胞核(nucleus)是遗传信息的载体,细胞的调节中心,其形态随细胞所处的周期阶段而异,通常以间期核为准. 细胞核外被核膜.核膜由内外二层各厚约3nm的单位膜构成,中间为2~5nm宽的间隙(核周隙);核膜上有直径约50nm的微孔,作为核浆与胞浆间交通的孔道,其数目因细胞类型和功能而异,多者可占全核表面积的25%;在肝细胞核据估算约有2000个核孔. 核浆主由染色质构成,其主要成分为DNA,并以与蛋白质相结合的形式存在,后者由组蛋白与非组蛋白组成.染色质的DNA现在已可用多种方法加以鉴定和定量测定. 核内较粗大浓缩的,碱性染料深染的团块状染色质为异染色质,呈细颗粒状弥散分布的,用普通染色法几乎不着色的染色质则为常染色质.一部分异染色质也可以上述两种状态存在.从生化角度看,异染色质不具遗传活性,相反,常染色质则大部分具遗传活性. 间期核的染色质模式还反映细胞的功能状态.一般而言,大而淡染的核(浓缩染色质少)提示细胞活性(如蛋白质和酶的合成)较高;小而深染的核(浓缩染色质较多)则提示细胞活性有限或降低. (一)细胞损伤时核的改变1,核大小的改变核的大小通常反映着核的功能活性状态,功能旺盛时核增大,核浆淡染,核仁也相应增大和(或)增多.如果这种状态持续较久,则可出现多倍体核或形成多核巨细胞.多倍体核在正常情况下亦可见于某些功能旺盛的细胞,如肝细胞中可见约20%为多倍体核.在病理状态下,如晚期肝炎及实验性肝癌前期等均可见多倍体的肝细胞明显增多. 核的增大除见于功能旺盛外,也可见于细胞受损时,最常见的情况为细胞水肿.这主要是细胞能量匮乏或毒性损伤所致,是核膜钠泵衰竭导致水和电解质运输障碍的结果.这种核肿大又称为变性性核肿大. 相反,当细胞功能下降或细胞受损时,核的体积则变小,染色质变致密,如见于器官萎缩时.与此同时核仁也缩小. 2.核形的改变光学显微镜下,各种细胞大多具有各自形状独特的核,可为圆形,椭圆形,梭形,杆形,肾形,印戒形,空洞形以及奇形怪状的不规则形等.在电镜下由于切片极薄,切面可以多种多样,但均非核的全貌.核的多形性和深染特别多见于恶性肿瘤细胞,称为核的异型性(atypia). 3.核结构的改变细胞在衰亡及损伤过程中的重要表征之一是核的改变,主要表现为核膜和染色质的改变. 核浓缩(karyopyknosis):染色质在核浆内聚集成致密浓染的大小不等的团块状,继而整个细胞核收缩变小,最后仅留下一致密的团块,是为核浓缩.这种浓缩的核最后还可再崩解为若干碎片(继发性核碎裂)而逐渐消失. 核碎裂(karyorrhexis):染色质逐渐边集于核膜内层,形成较大的高电子密度的染色质团块.核膜起初尚保持完整,以后乃在多处发生断裂,核逐渐变小,最后裂解为若干致密浓染的碎片. 核溶解(karyolysis):变致密的结成块状的染色质最后完全溶解消失,即核溶解.核溶解也可不经过核浓缩或核碎裂而一开始即独立进行.在这种情况下,受损的核很早就消失. 上述染色质边集(即光学显微镜下所谓的核膜浓染),核浓缩,核碎裂,核溶解等核的结构改变为核和细胞不可复性损伤的标志,提示活体内细胞死亡(坏死). 4.核内包含物(intranuclear inclusions) 在某些细胞损伤时可见核内出现各种不同的包含物,可为胞浆成分(线粒体,内质网断片,溶酶体,糖原颗粒,脂滴等),亦可为非细胞本身的异物,但最常见的还是前者. 这种胞浆性包含物可在两种情况下出现: ①胞浆成分隔着核膜向核内膨突,以致在一定的切面上看来,似乎胞浆成分已进入核内,但实际上大多仍可见其周围有核膜包绕,其中的胞浆成分常呈变性性改变(如髓鞘样结构,膜碎裂等).这种包含物称为胞浆性假包含物; ②在有丝分裂末期,某些胞浆结构被封入形成中的子细胞核内,以后出现于子细胞核中,称为真性胞浆性包含物. 非胞浆性(异物性)核内包含物的种类繁多,性质各异.在真性糖尿病时,肝细胞核内可有较多糖原沉积.在常规切片制作过程中,糖原被溶解,核内出现或大或小的空洞(糖尿病性空洞核). 在铅,铋,金等重金属中毒时,核内亦可出现丝状或颗粒状真性包含物,其中有时含有相应的重金属(如铅中毒时). 此外,在某些病毒性疾病如DNA病毒感染时,可在电镜下检见核内病毒颗粒,如聚积成较大集团(如巨细胞包涵体病),则亦可在光学显微镜下检见,表现为较大的核内包含物. 5.核仁的改变核仁(nucleolus)为核蛋白体RNA转录和转化的所在.除含蛋白的均质性基质外,电镜下核仁主由线团状或网状电子致密的核仁丝(nucleolonema)和网孔中无结构的低电子密度的无定形部(pars amorpha)组成.核仁无界膜,直接患浮于核浆内. 形态学和生物学上核仁由3种不同的成分构成:①原纤维状成分,内含蛋白质及与其相结合的45S-rRNA;②细颗粒状成分,主要由12S-rRNA构成,为核仁的嗜碱性成分;③细丝状成分,仅由来自胞浆的蛋白质构成,穿插于整个核仁内. 3种核仁成分的空间排列状态可反映细胞的蛋白合成活性,例如:壳状核仁:原纤维状成分集中位于核仁中央,细颗状成分呈壳状包绕于外层.这种细胞的合成活性甚低. 海绵状核仁:这种核仁的原纤维状与细颗状成分呈海绵状(或线团状)排列.这种细胞的合成活性升高.大多数所谓的"工作核"具有这种核仁. 高颗粒性核仁:由海绵状核仁转化而成,原纤维状成分几乎消失,核仁主要由颗粒状成分构成,故组织学上呈强嗜碱性,细胞的合成活性旺盛.这种核仁常见于炎症和肿瘤细胞. 低颗粒性核仁:与上述高颗粒性核仁相反,这种核仁的细颗粒状成分锐减,故电镜下原纤维状成分显得突出,电子密度较低.这种核仁常见于再生时,因此时细颗粒成分(rRNA)过多地被胞浆所利用. 分离性核仁:超微结构上3种核仁成分清楚地互相分离,原纤维状和细颗粒状成分减少.这种核仁变小,无活性,常见于核仁转录过程被抗生素,细胞抑制剂,缺氧和蝇菌素中毒等所完全阻断时. 由此可见,核仁的大小和(或)数目的多少常反映细胞的功能活性状态:大和(或)多的核仁是细胞功能活性高的表现,反之则细胞功能活性低.二、细胞膜细胞膜是包于细胞表面,将细胞与周围环境隔开的弹性薄膜,厚约8~10nm,由脂质和蛋白质构成,故为脂蛋白膜,对于细胞的生命活动和功能具有十分重要的意义. 细胞膜在许多特定场合可向外形成大量的纤细突起(微绒毛,纤毛),或向内形成各种形式的内褶,以利于其功能活动.侧面则特化为细胞连接. 相邻细胞的细胞膜之间连接包括闭锁小带(紧密连接),附着小带(中间连接),桥粒和缝隙(管)连接等各种特化结构,以保持细胞间的联系. 此外,新近还发现,在相邻细胞膜上有"粘附分子(如cadherin)",对细胞正常结构和联系以及细胞极性的维持和细胞的分化等,均具有重要作用. 细胞膜除作为细胞的机械性和化学性屏障外,还具有一系列重要的功能诸如细胞内外的物质交换,细胞运动,细胞识别以及细胞的生长调控,免疫决定和各种表面受体形成等. 细胞的物质交换:细胞内外的物质交换主要以两种方式进行,一为渗透,一为出入胞过程. 渗透乃指低分子物质(主要为水和电解质)通过细胞膜进出细胞,又可分两种情况:一种是按该物质在细胞内外环境中的浓度差,由浓高的一侧弥散底到低的一侧(被动运输); 另一种则逆浓度差进行,即由浓度低的一侧向浓度高的一侧输送(主动运输),其经典的例子即Na+和K+的运输(在细胞内Na+和K+的比例为1:20,而在细胞外隙则为30:1),即依靠所谓"钠泵"的作用将Na+ 移向细胞外隙,而使K+移向细胞内. 这种主动运输是一个耗能的过程,并由Na+和K+激活细胞膜上的ATP酶分解ATP而提供所需的能量.因此,如ATP酶受到某些毒物的抑制,则这种主动运输过程也同样受到阻抑.除Na+和K+外,其他一些有机物质如葡萄糖,氨基酸以及一些低分子代谢产物也是借这样的过程运输的. 第二种物质运输方式为出入胞过程.较大的分子和颗粒不能借渗透过程通过细胞膜,乃借出,入胞过程将细胞内物质运送到细胞外和将细胞外物质移入细胞内. 前者称为出胞(exocytosis),后者称为入胞(endocytosis).进入细胞的如为液态物质则称之为胞饮或吞饮(pinocytosis),如为固体颗粒(如细菌,尘粒等异物)则称之为吞噬(phagocytosis). 在吞饮过程中,被吞饮的物质先接触并附着于细胞膜上,然后该处细胞膜连同该物质内凹,继而从细胞膜上断离下来,在胞浆内形成有膜包绕的小泡(吞饮小泡); 吞噬过程与吞饮相似,稍不同的是,被吞噬物附着于细胞膜上后,细胞膜乃形成伪足样突起,将该物质环抱,最后封闭成有膜包绕的泡状结构,从而将该物质移入胞浆内(吞噬泡或吞噬体). 细胞自身的成分如蛋白质分子,糖原颗粒,衰变的或受损而待处理的细胞器等,亦可被膜包绕而形成自噬泡(autophagic vacuoles)或自噬体(autophagosome). 胞饮泡或吞噬泡一般在胞浆内与溶酶体相结合,并被溶酶体酶所降解消化.但胞饮泡也可不经处理而穿过胞浆,最后从细胞的另一极重新移出细胞外. 细胞膜上还有特殊的识别区,结合在糖萼上,借此,细胞可相互识别,从而相互接近形成一定的细胞组合,或相互排斥而分离. 同样,通过识别区,增生中的细胞在互相接触时就会停止分裂(接触抑制),而癌细胞则已失去这种表面功能,故可不受限制地增生. 此外,细胞膜上还有一种膜抗原可以识别"自我"和"非我"(组织相容性抗原).这种膜抗原在器官移植中具有重要意义,因为它可致敏受体,从而引起对移植物的排斥反应. 细胞上还有一些特异性区域带着特殊的化学簇,可以接受相应的化学信号,称为膜受体或表面受体.但从形态学上不能辨认. 这种膜受体具有十分重要的意义,因为已知许多物质如激素,免疫球蛋白,药物,毒素以及感染因子等都是作为外来信号被受体接受后才转化为细胞内效应而发挥其作用.如封闭其受体,则亦同时消除其作用. 细胞膜的病变 1.细胞膜形态结构的改变机械力的作用或细胞强烈变形,可引起红细胞膜的破损,如人工心瓣膜可引起细胞膜的破裂;某些脂溶性阴离子物质,溶蛋白和溶脂性酶以及毒素等也能破坏细胞膜的完整性.细胞膜结构的损伤可导致细胞内容物的外溢或水分进入细胞使细胞肿胀. 2.细胞膜通透性的改变能量代谢不足(如缺氧时)或毒物的直接损害等所致各种不同的细胞损伤时,均可造成细胞主动运输的障碍,从而导致细胞内Na+的潴留和K+的排出,但Na+的潴留多于K+的排出,使细胞内渗透压升高,水分因而进入细胞,引起细胞水肿. 这种单纯的通透性障碍时并不见细胞膜的形态学改变,只有借细胞化学方法才可在电镜下检见细胞膜上某些酶(如ATP酶,碱性磷酸酶,核苷酸酶等)活性的改变. 当然,如损伤或水肿严重,则亦可发生继发性形态改变如出现胞浆膨出,微绒毛变短甚至消失,细胞膜基底变平乃至细胞膜破裂等.在某些较严重的损伤时还可出现细胞膜的螺旋状或同心圆层状卷曲,形成典型的髓鞘样结构(myelin figure). 3.细胞内和细胞间腺管结构的形成细胞质l各种细胞器l包涵物l基质三、内质网除红细胞外,内质网或多或少地见于所有各种细胞.内质网为由生物膜构成的互相通连的片层隙状或小管状系统,膜片间的隙状空间称为池,通常与细胞外隙和细胞浆基质之间不直接相通. 这种细胞内的膜性管道系统一方面构成细胞内物质运输的通路,另一方面为细胞内各种各样的酶反应提供广阔的反应面积.内质网与高尔基体及核膜相连续. (一)粗面内质网在病理状态下,粗面内质网(RER)可发生量和形态的改变.在蛋白质合成及分泌活性高的细胞(如浆细胞,胰腺腺泡细胞,肝细胞等)以及细胞再生和病毒感染时,粗面内质网增多. 粗面内质网的含量高低也常反映肿瘤细胞的蛋白质合成功能的状态,并在一定程度上反映了肿瘤细胞的分化程度.如恶性程度较高的骨肉瘤细胞中,粗面内质网十分发达. 当细胞受损时,粗面内质网上的核蛋白体往往脱落于胞浆内,粗面内质网的蛋白合成乃下降或消失;当损伤恢复时,其蛋白合成也随之恢复. 在由各种原因引起的细胞变性和坏死过程中,粗面内质网的池一般出现扩张,较轻的和局限性的扩张只有在电镜下才能窥见,重度扩张时则在光学显微镜下可表现为空泡形成,电镜下有时可见其中含有中等电子密度的絮状物. 在较强的扩张时,粗面内质网同时互相离散,膜上的颗粒呈不同程度的脱失.进而内质网本身可断裂成大小不等的片段和大小泡.这些改变大多见于细胞水肿时,故病变不仅见于内质网,也同时累及Golgi器,线粒体和胞浆基质,有时甚至还累及溶酶体. (二)滑(光)面内质网光面内质网的功能多种多样,即参与糖原的合成,又能合成磷脂,糖脂以及糖蛋白中的糖成分,此外,还在类固醇化合物的合成中起重要的作用,故在合成类固醇激素的细胞中特别丰富. 光面内质网含有脱甲基酶,脱羧酶,脱氨酶,葡糖醛酸酶以及混合功能氧化酶等,因而光面内质网能分解甾体,能灭活药物和毒物并使其能被排除(如肝细胞).肠上皮细胞的光面内质网参与脂肪的运输,心肌细胞的光面内质网(肌浆网)则参与心肌的刺激传导. 在生理状态下,随着细胞功能的升降,光面内质网(SER)的数量也呈现相应改变.但亦可出现完全相反的情况,例如在某些疾病(如淤胆)时,从形态结构上看,肝细胞光面内质网显著增生,但其混合功能氧化酶的活性反而下降,这实际上是细胞衰竭的表现. 肝细胞的光面内质网具有生物转化作用,能对一些低分子物质如药物,毒品,毒物等,进行转化解毒,并将间接胆红素转化为直接胆红素. 许多成瘾药物和嗜好品如巴比妥类,吸毒,嗜酒等,可导致肝细胞光面内质网的增生,长期服用口服避孕药,安眠药,抗糖尿病药等也能导致同样后果. 某些肿瘤细胞也可见增生扩张的滑面内质网,如在垂体前叶嗜酸细胞腺瘤的瘤细胞内,就可见大量扩张的滑面内质网. 在HBsAg阳性肝炎时,肝细胞内光面内质网明显增生,在其管道内形成HBsAg.由于光面内质网的大量增生,这种肝细胞在光学显微镜下呈毛玻璃外观,故有毛玻璃细胞之称,并可为地衣红(orcein)着染. 在细胞损伤时光面内质网也可出现小管裂解为小泡或扩大为大泡状.在药物及某些芳香族化合物(主为致癌剂)的影响下,光面内质网有时可在胞浆内形成葱皮样层状结构,即"副核",可为细胞的适应性反应(结构较松)或为变性性改变(结构致密).四、线粒体线粒体(mitochondrion)是细胞内主要的能量形成所在,故不论在生理上或病理上都具有十分重要的意义. 线粒体为线状,长杆状,卵圆形或圆形小体,外被双层界膜.外界膜平滑,内界膜则折成长短不等的嵴并附有基粒.内外界膜之间为线粒体的外室,与嵴内隙相连,内界膜内侧为内室(基质室). 在合成甾类激素的内分泌细胞(如肾上腺皮质细胞,卵巢滤泡细胞,睾丸的Leydig细胞等),线粒体嵴呈小管状. 内外界膜的通透性不同,外界膜的通透性高,可容许多种物质通过,而内界膜则构成明显的通透屏障,使一些物质如蔗糖和NADH全然不能通过,而其他物质如Na+ 和Ca 2+等也只有借助于主动运输才能通过. 线粒体的基质含有电子致密的无结构颗粒(基质颗粒),与二价阳离子如Ca2+及Mg2+ 具有高度亲和力.基质中进行着β氧化,氧化脱羧,枸橼酸循环以及尿素循环等过程. 在线粒体的外界膜内含有单胺氧化酶以及糖和脂质代谢的各种转移酶;在内界膜上则为呼吸链和氧化磷酸化的酶类. 线粒体是对各种损伤最为敏感的细胞器之一.在细胞损伤时最常见的病理改变可概括为线粒体数量,大小和结构的改变: 1.数量的改变线粒体的平均寿命约为10天.衰亡的线粒体可通过保留的线粒体直接分裂为二予以补充. 在病理状态下,线粒体的增生实际上是对慢性非特异性细胞损伤的适应性反应或细胞功能升高的表现.例如心瓣膜病时的心肌线粒体,周围血液循环障碍伴间歇性跛行时的骨骼肌线粒体的呈增生现象. 线粒体的增生也可见于某些肿瘤组织,如甲状腺,肾,脑垂体等处发生的嗜酸性腺瘤(oncocytoma).瘤细胞内可见大量线粒体. 线粒体数量减少则见于急性细胞损伤时线粒体崩解或自溶的情况下,持续约15分钟.慢性损伤时由于线粒体逐渐增生,故一般不见线粒体减少(甚至反而增多).此外,线粒体的减少也是细胞未成熟和(或)去分化的表现. 2.大小改变细胞损伤时最常见的改变为线粒体肿大.根据线粒体的受累部位可分为基质型肿胀和嵴型肿胀二种类型,而以前者为常见. 基质型肿胀时线粒体变大变圆,基质变浅,嵴变短变少甚至消失.在极度肿胀时,线粒体可转化为小空泡状结构.此型肿胀为细胞水肿的部分改变.光学显微镜下所谓的浊肿细胞中所见的细颗粒即肿大的线粒体. 嵴型肿胀较少见,此时的肿胀局限于嵴内隙,使扁平的嵴变成烧瓶状乃至空泡状,而基质则更显得致密.嵴型肿胀一般为可复性,但当膜的损伤加重时,可经过混合型而过渡为基质型. 线粒体为对损伤极为敏感的细胞器,其肿胀可由多种损伤因子引起,其中最常见的为缺氧;此外,微生物毒素,各种毒物,射线以及渗透压改变等亦可引起.但轻度肿大有时可能为其功能升高的表现,较明显的肿胀则恒为细胞受损的表现.但只要损伤不过重,损伤因子的作用不过长,肿胀仍可恢复. 线粒体的增大有时是器官功能负荷增加引起的适应性肥大,此时线粒体的数量也常增多,例如见于器官肥大时.反之,器官萎缩时,线粒体则缩小,变少.在某些病理条件下,其体积也可增大. 3.结构的改变线粒体嵴是能量代谢的明显指征,但嵴的增多未必均伴有呼吸链酶的增加.嵴的膜和酶平行增多反映细胞的功能负荷加重,为一种适应状态的表现;反之,如嵴的膜和酶的增多不相平行,则是胞浆适应功能障碍的表现,此时细胞功能并不升高. 在急性细胞损伤时(大多为中毒或缺氧),线粒体的嵴被破坏;慢性亚致死性细胞损伤或营养缺乏时,线粒体的蛋白合成受障,以致线粒体几乎不再能形成新的嵴. 根据细胞损伤的种类和性质,可在线粒体基质或嵴内形成病理性包含物. 这些包含物有的呈晶形或副晶形(可能由蛋白构成),如在线粒体性肌病或进行性肌营养不良时所见;有的呈无定形的电子致密物,常见于细胞趋于坏死时,乃线粒体成分崩解的产物(脂质和蛋白质),被视为线粒体不可复性损伤的表现. 线粒体损伤的另一种常见改变为髓鞘样层状结构的形成,这是线粒体膜损伤的结果. 衰亡或受损的线粒体,最终由细胞的自噬过程加以处理并最后被溶酶体酶所降解消化.五、高尔基体高尔基体(Golgi apparatus)见于一切有核细胞,来自核膜外层,由数列弯曲成蹄铁状的扁平囊组成,在横切面上表现为光面双膜,其末端膨大成烧瓶状.高尔基体面向核的一面称为形成面,由许多与粗面内质网池相连的小泡构成.另一面称为成熟面,由此断下一些较大的泡,内含分泌物. 由粗面内质网合成的蛋白质输送到此,经加工装配形成分泌颗粒,分泌到细胞外,例如肝细胞合成的白蛋白和脂蛋白即按此方式形成和输出. 此外,细胞本身的酶蛋白如溶酶体的水解酶类也是这样,但却不装配成分泌颗粒和排出细胞外,而是以高尔基小泡的形式(初级溶酶体,前溶酶体)输送到各种吞噬体中.高尔基体在形成含糖蛋白的分泌物中,在构成细胞膜及糖萼中,以及在形成结缔组织基质中也均起着重要的作用. 高尔基体的病变 1.高尔基体肥大见于细胞的分泌物和酶的产生旺盛时.巨噬细胞在吞噬活动旺盛时,可形成许多吞噬体,高尔基体增多,并从其上断下许多高尔基小泡. 2.高尔基体萎缩在各种细胞萎缩时,可见高尔基体变小和部分消失.其中扁平囊可看不到. 3.高尔基体扩张高尔基体损伤时,大多出现扁平囊的扩张以及扁平囊,大泡和小泡的崩解.。
细胞超微结构范文细胞是构成生物体的最基本单位,能够进行代谢、生长、分裂和传递遗传信息。
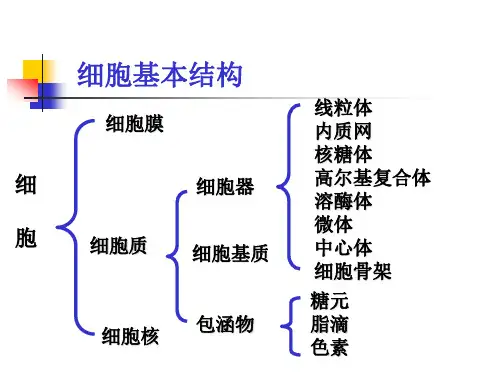
细胞超微结构指的是细胞内部的细节和组织结构,包括细胞膜、细胞质、细胞器和细胞核等。
以下将详细介绍细胞超微结构的各个组成部分。
细胞膜是细胞的外包层,是细胞与外界环境之间的分界线。
细胞膜由磷脂双分子层构成,其中嵌入着蛋白质和其他生物分子。
细胞膜具有选择性通透性,可以控制物质的进出。
此外,细胞膜还参与信号传导和细胞黏附等重要生理过程。
细胞质是细胞膜内的液体,包含有各种溶质、细胞器以及细胞骨架等。
细胞质内有许多胞浆小滴,其中含有许多溶解物质并参与代谢活动。
细胞质还包含有细胞骨架,细胞骨架由微丝、中间丝和微管组成,赋予细胞形态支持和运动功能。
细胞器是细胞内的各种结构,具有不同的功能。
其中,线粒体是细胞内的主要能量生产器,通过细胞呼吸产生ATP分子。
高尔基体参与蛋白质的合成、修饰和包装,细胞内的内分泌物质也由高尔基体合成。
溶酶体是细胞的“垃圾处理厂”,负责分解陈旧或损坏的细胞组分。
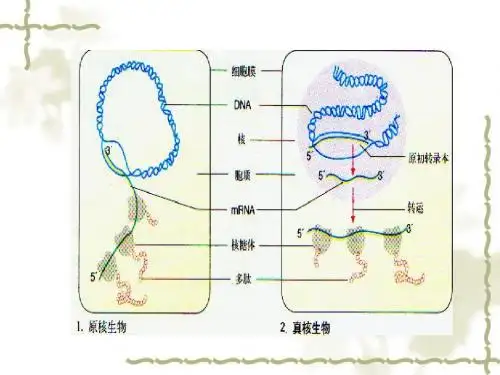
内质网是一种连通的膜系统,参与蛋白质的合成和转运。
核糖体则是合成蛋白质的场所,通过转录和翻译过程,将基因中的遗传信息转化为蛋白质。
细胞核是细胞中最重要的结构之一,存储着遗传信息。
细胞核由核膜、染色质和核仁组成。
核膜是一个由两层磷脂双分子层构成的膜,分别包围着外核膜和内核膜之间的核间隙。
核膜上有许多核孔,允许物质的进出。
染色质则是细胞核内的DNA分子和相关蛋白质的复合体,包含了细胞的遗传信息。
核仁则是细胞中的核小体组成的区域,参与蛋白质的合成和组装过程。
总之,细胞超微结构是细胞内部各个组成部分的组织和结构。
细胞膜、细胞质、细胞器和细胞核等都起着重要的功能作用,共同维持细胞的生命活动和遗传信息的传递。
对于深入了解生物体机制和细胞功能的研究非常关键。
实验八1细胞超微结构观察与透射电子显微镜的使细胞超微结构观察与透射电子显微镜的使用引言:细胞是构成生物体的基本单位,是人们研究生命现象和生理特点的重要对象。
为了更好地了解细胞的组成和结构,科学家们发展了不同的显微镜技术,其中透射电子显微镜(Transmission Electron Microscope, TEM)成为观察细胞超微结构的最高分辨率显微镜。
本实验旨在通过使用透射电子显微镜,观察和研究细胞超微结构,以加深对细胞组成的理解。
一、实验目的通过透射电子显微镜观察和研究细胞超微结构,加深对细胞组成的理解。
二、实验步骤1.将样品制备成细切片。
2.将细切片固定在铜网上。
3.将样品放入透射电子显微镜中。
4.调整透射电子显微镜的参数,如加速电压和对比度。
5.通过透射电子显微镜观察细胞超微结构,并进行拍照记录。
6.分析和解读观察到的细胞超微结构。
三、实验结果和讨论在透射电子显微镜下观察到的细胞超微结构有以下几个重要组成部分。
1. 细胞核(Nucleus):细胞核是细胞的控制中心,包含DNA分子和相关蛋白质。
在透射电子显微镜下,细胞核可见为一个圆形或椭圆形的结构,外部由双层核膜包围,内部含有染色质和核仁。
2. 内质网(Endoplasmic Reticulum, ER):内质网是一种细胞内膜系统,参与蛋白质合成和运输。
在透射电子显微镜下,内质网可见为一系列连接的膜囊泡、膜管或平片结构。
3. 线粒体(Mitochondria):线粒体是细胞内的能量发生器,参与细胞的呼吸作用。
在透射电子显微镜下,线粒体可见为一个椭圆形的结构,由内外两层膜组成,内膜上有许多呼吸酶。
4. 高尔基体(Golgi Apparatus):高尔基体是细胞内的分泌系统,参与蛋白质的修饰和分泌。
在透射电子显微镜下,高尔基体可见为一系列扁平的膜囊泡。
细胞超微结构的观察和研究有助于我们深入了解细胞的功能和活动。
透射电子显微镜具有高分辨率和高放大倍数的特点,能够观察到细胞内部微观结构的细节,如蛋白质的精确位置和细胞器之间的相互作用。