纤维素的结晶结构
- 格式:ppt
- 大小:2.84 MB
- 文档页数:48


纤维素的结构引言纤维素是地球上存在的最丰富的可再生有机资源, 在高等植物、细菌、动物、海藻等生物中广泛存在, 每年总量有几百亿吨, 具有巨大的经济开发价值[1]。
五十年代至六十年代,由于合成高分子材料的兴起,纤维素资源的开发研究受到极大的影响。
七十年代初期,由于国际上出现了石油危机,这种曾被忽视的可更新资源又再次被重视起来.能否利用这些丰富的可再生资源是解决未来能源问题的关键因素。
因此,世界各国都很重视纤维素的研究与开发[2]。
纤维素结构是纤维素性能研究及应用的基础,本文就纤维素的化学剂物理结构进行了概述。
1纤维素的化学结构纤维素的元素组成为:C=44.44%,H=6.17%,O=49.39%, 其化学实验式(C 6H 10O 5)n (n 为聚合度,一般高等植物纤维素的聚合度为7000—150000)[3]纤维素大分子的基环是脱水葡萄糖,其分子式为(C 6H 10O 5)。
纤维素的化学结构是由D-吡喃葡萄糖环彼此以β- 1, 4-糖苷键以C1椅式构象联结而成的线形高分子化合物[4],其结构表达式如图1所示。
非还原端 纤维二糖 还原端图1 纤维素链结构除两端的葡萄糖基外,每个葡萄糖基上都有三个游离羟基,分别位于C 2、C 3和C 6位上,所以纤维素的分子可以表示为[[C 6H 7O 2(OH)3]n,其中C 2和C 3位上为仲醇羟基,C 6位上为伯醇羟基,他们的反应能力不同,对纤维素的性质具有重要影响,如纤维素的酯化、醚化、氧化和接枝共聚,以及纤维素之间的分子间氢键作用,纤维素的溶胀与水解都与纤维素的羟基有关。
纤维素大分子两端的葡萄糖末端基,其结构和性质不同,一端的葡萄糖末端基在C4上存在一个苷羟基,此羟基的氢原子易转移,与基环上的氧原子结合,使氧环结构转变为开链式结构,在C1处形成醛基,具有潜在还原性,固有隐形醛基之称。
左端的葡萄糖末端为非还原性的,由于纤维素的每一个分子链一端是还原性,另一端是非还原性,所以纤维素分子具有极性和方向性。





纤维素水解
纤维素水解是一个广泛应用于工业和生物科学领域的过程。
纤维素是一种多糖
类聚合物,主要存在于植物细胞壁中,包括木质素和纤维素。
纤维素水解是将纤维素分解为更简单的单糖,如葡萄糖,以便更好地利用其作为生物质资源。
纤维素的结构
纤维素是由葡萄糖分子通过β-1,4-糖苷键连接而成的线性多糖,具有高度的结
晶性和稳定性。
这种结构赋予了纤维素出色的机械强度和耐久性,同时也增加了其降解的难度。
纤维素水解的方法
纤维素水解通常采用酶解法和酸解法两种主要方法。
酶解法
酶解法是目前应用最为广泛的纤维素水解方法之一。
在酶解过程中,纤维素酶
通过降解纤维素的β-1,4-糖苷键来将纤维素水解为葡萄糖。
常用的纤维素酶包括纤
维素酶、β-葡聚糖酶等。
酶解法具有选择性高、反应条件温和等优点,但同时也存在酶的稳定性、成本等方面的挑战。
酸解法
酸解法是另一种纤维素水解的方法,通过在酸性条件下将纤维素水解成葡萄糖。
常用的酸包括硫酸、盐酸等。
酸解法具有操作简单、反应速度快等优点,但会产生大量的废弃物,并对环境造成污染。
纤维素水解的应用
纤维素水解是生物质能源利用的重要途径之一。
通过将纤维素水解成葡萄糖,
可以进一步转化为乙醇、生物柴油等可再生燃料。
同时,纤维素水解产生的糖类还可以用于生物化学品和生物材料的生产,促进生物经济的发展。
纤维素水解技术的不断发展将为可再生能源和生物资源开发提供更多可能性,
促进绿色和可持续发展的实现。

天然纤维素结晶变体I结晶变体II简谈摘要:纤维素(celluloe)是由葡萄糖组成的大分子多糖。
不溶于水及一般有机溶剂。
是植物细胞壁的主要成分。
在自然。
具有一定构象的纤维素高分子链按一定的秩序堆砌,便成为纤维素的微晶体,微晶体的组成单元称为晶胞。
在纤维素中存在着化学组成相同,而单元晶胞不同的同质多晶体(结晶变体),常见的结晶变体有四种,即纤维素Ⅰ、Ⅱ、Ⅲ、Ⅳ。
本文将这种介绍纤维素Ⅰ、纤维素Ⅱ之间的转化。
关键字:纤维素结晶变体转化结构1、简述纤维素纤维素是由D-吡喃型葡萄糖基(失水葡萄糖)组成。
简单分子式为(C6H10O5)n;化学结构式可用下二式表示:霍沃思式是由许多D-葡萄糖基(1-5结环),藉1-4,β-型联结连接起来的,而且连接在环上碳原子两端的OH和H位置不相同,所以具有不同的性质。
式中n为聚合度。
在天然纤维素中,聚合度可达10000左右;再生纤维素的聚合度通常为200~800。
在一个样品中,各个高分子的聚合度可以不同,具有多分散性。
椅式由于内旋转作用,使分子中原子的几何排列不断发生变化,产生了各种内旋转异构体,称为分子链的构象。
纤维素高分子中,6位上的碳-氧键绕5和6位之间的碳-碳键旋转时,相对于5位上的碳-氧键和5位与4位之间的碳-氧键可以有三种不同的构象。
如以g表示旁式,t表示反式,则三种构象为gt、tg、和gg。
多数人认为,天然纤维素是gt构象,再生纤维素是tg构象。
在纤维素分子链中,存在着氢键。
这种氢键把链中的O6(6位上的氧)与O2'以及O3与O5'连接起来使整个高分子链成为带状,从而使它具有较高的刚性。
在砌入晶格以后,一个高分子链的O6与相邻高分子的O3之间也能生成链间氢键。
2、纤维素结晶变体及其结构纤维素的聚集态结构是研究纤维素分子间的相互排列情况(晶区和非晶区、晶胞大小及形式、分子链在晶胞内的堆砌形式、微晶的大小)、取向结构(分子链和微晶的取向)等。
天然纤维素和再生纤维素纤维都存在结晶的原纤结构,由原先结构及其特性可部分地推知纤维的性质,所以为了解释以纤维素为基质的材料的结构与性能关系,寻找制备纤维素衍生物的更有效方法,则研究纤维素合成的机理、了解纤维素的聚集态结构,在理论研究和实际应用方面都有重要的意义。
互穿聚合物网络(IPN)天然纤维素包括细菌纤维素、海藻和高等植物(如棉花、苎麻、木材等)均属于纤维素I型。
纤维素I分子链在晶胞内是平行堆砌的,纤维素II是纤维素I经由溶液中再生(regeneration)或经丝光处理(mercerization)得到的结晶变体,是工业上使用最多的纤维素形式。
纤维素II 与纤维素I有很大的不同,它是由两条分子链组成的单斜晶胞,属于反平行链的堆砌。
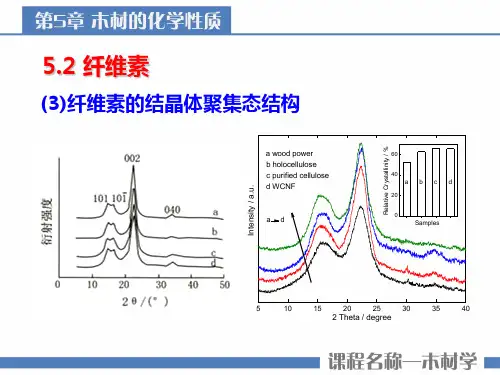
纤维素是一种β-(1-4)-D-糖苷键连接的线型高聚物,由X 射线衍射发现存在四种结晶形态,即纤维素Ⅰ、Ⅱ、Ⅲ和Ⅳ,不同晶型纤维素的C1,C4 和C6 的化学位移具有明显的差别,这种差别可能是因为不同晶型纤维素的链构象转变或晶体堆砌对吡喃葡萄糖单元C4 和C6 的影响差异造成的.基于在非晶区的链段运动显示窄谱线,而晶区的刚性链以及分布在非晶区的刚性链产生宽谱线,因此可以利用宽、窄谱线的峰面积(S b 和S n)求取结晶度χ c纤维素I和纤维素II在C6上的差别就是因为吡喃葡萄糖单元C6位羟基的构象不同,纤维素I为t - g构象,纤维素II和无定型纤维素则为g - t构象。
天然纤维素I也存在两种不同的晶体结构,即纤维素Iα和Iβ。
13C NMR谱指出它们之间最大的差别在C1的化学位移上,Iα 为单峰,Iβ 为双峰。
高分子链构象参数:无扰尺寸(A):A 值愈小,高分子链愈柔顺。
A 值只取决于高分子的近程结构,与高聚物的分子量无关。
空间位阻参数(σ):空间位阻参数是指由于高分子链的内旋转受阻而导致分子尺寸增大程度的量度,其值愈小,高分子链愈柔顺Flory 极限特征比(C∞ ):Flory 极限特征比是指高分子链由于键角限制和空间位阻造成分子链伸展的程度,。
一般来说,C∞ 愈小,链愈柔顺。
合成的柔顺性高聚物的C 值在5~7 范围内,而天然高分子多数高于此范围。
持续长度(q):持续长度q 广义为高分子链在第一个键方向上的投影,它表征分子链的支撑能力。

晶纤维名词解释-概述说明以及解释1.引言1.1 概述晶纤维是一种具有晶体结构的纤维材料,其特点是具有高强度、高模量、低比重和优异的耐热性能。
它由纳米级晶体构成,这些晶体在纤维中具有有序排列的特点,使得晶纤维具有出色的力学性能和优异的热传导性能。
晶纤维的制备方法多样,可以通过溶剂法、湿法纺丝法、干法纺丝法等不同工艺得到。
晶纤维在各个领域都有广泛的应用。
在纺织领域,晶纤维可以用于制作高强度的纺织品,如防弹衣、护甲等。
此外,晶纤维还可以用于制作高性能的绳索和缆绳,被广泛应用于登山、航海等领域。
在航空航天领域,晶纤维则可以用于制作航空器的结构材料,因其轻便和高强度的特点而备受青睐。
同时,晶纤维的耐高温性能也使其成为航空发动机喷嘴等高温部件的理想材料。
可以预见,随着科技的不断进步和新材料的开发,晶纤维在未来将有更广阔的应用前景。
其出色的力学性能和耐热性能使其在新能源、新材料、生物医药等领域的应用成为可能。
与此同时,随着制备工艺的不断改进和成本的降低,晶纤维的商业化生产将更加普及,为各行各业带来更多的机遇和挑战。
综上所述,本文将对晶纤维的定义、制备方法以及应用领域进行详细的介绍和解释。
通过深入了解晶纤维的特点和优势,我们能够更好地掌握和应用这一材料,为各行业的发展和创新做出更大的贡献。
文章结构部分内容的编写如下:1.2 文章结构本文将按照以下结构展开论述晶纤维的相关内容:2.1 晶纤维定义:首先,我们将详细解释晶纤维的定义,并介绍其在纤维材料中的特殊性质和结构。
2.2 晶纤维制备方法:接下来,我们将探讨晶纤维的制备方法,包括熔融法、湿法和干法等不同的制备技术,并对各种方法的优缺点进行比较和分析。
2.3 晶纤维应用领域:然后,我们将介绍晶纤维在各个领域的广泛应用,包括纺织、建筑、医疗等方面,以及其在未来发展中的潜在应用领域。
3.结论3.1 对晶纤维的重要性进行总结:在本部分,我们将总结晶纤维在现代社会中的重要性,强调其在工业、科学研究和生活中的关键作用。
天然纤维素结晶变体I、结晶变体II简谈(作业1)摘要:纤维素(cellulose)是由葡萄糖组成的大分子多糖。
不溶于水及一般有机溶剂。
是植物细胞壁的主要成分。
在自然。
具有一定构象的纤维素高分子链按一定的秩序堆砌,便成为纤维素的微晶体,微晶体的组成单元称为晶胞。
在纤维素中存在着化学组成相同,而单元晶胞不同的同质多晶体(结晶变体),常见的结晶变体有四种,即纤维素Ⅰ、Ⅱ、Ⅲ、Ⅳ。
本文将这种介绍纤维素Ⅰ、纤维素Ⅱ之间的转化。
关键字:纤维素结晶变体转化结构1、简述纤维素纤维素是由D-吡喃型葡萄糖基(失水葡萄糖)组成。
简单分子式为(C6H10O5)n;化学结构式可用下二式表示:霍沃思式是由许多D-葡萄糖基(1-5结环),藉1-4,β-型联结连接起来的,而且连接在环上碳原子两端的OH和H位置不相同,所以具有不同的性质。
式中n为聚合度。
在天然纤维素中,聚合度可达10000左右;再生纤维素的聚合度通常为200~800。
在一个样品中,各个高分子的聚合度可以不同,具有多分散性。
椅式由于内旋转作用,使分子中原子的几何排列不断发生变化,产生了各种内旋转异构体,称为分子链的构象。
纤维素高分子中,6位上的碳-氧键绕5和6位之间的碳-碳键旋转时,相对于5位上的碳-氧键和5位与4位之间的碳-氧键可以有三种不同的构象。
如以g表示旁式,t表示反式,则三种构象为gt、tg、和gg。
多数人认为,天然纤维素是gt构象,再生纤维素是tg构象。
在纤维素分子链中,存在着氢键。
这种氢键把链中的O6(6位上的氧)与O2'以及O3与O5'连接起来使整个高分子链成为带状,从而使它具有较高的刚性。
在砌入晶格以后, 一个高分子链的O6与相邻高分子的O3之间也能生成链间氢键。
2、纤维素结晶变体及其结构纤维素的聚集态结构是研究纤维素分子间的相互排列情况(晶区和非晶区、晶胞大小及形式、分子链在晶胞内的堆砌形式、微晶的大小)、取向结构(分子链和微晶的取向)等。
纤维素的大分子结构纤维素是一种由β-葡萄糖单体组成的天然聚合物大分子。
它是地球上最常见的有机化合物之一,在植物细胞壁中起着关键的结构和功能作用。
纤维素的大分子结构决定了它的物理性质和化学性质,对于理解纤维素的特性和应用至关重要。
纤维素的大分子结构是由若干个β-葡萄糖单体通过β-1,4-连接键连接而成的线性聚合物。
β-葡萄糖单体有两个C1和C4碳原子,它们通过氧原子形成1,4-葡萄糖醚键。
这种键的特殊性决定了纤维素的特殊性质,如生物降解性和高强度。
纤维素的结构中的OH基团没有被化学修饰,因此纤维素是一种天然的、无毒的高分子化合物。
纤维素在自然界中主要存在于植物细胞壁中。
在植物细胞中,纤维素通常以微纤的形式存在,形成了复杂的网状结构。
纤维素的微纤具有一定的直径和长度,纤维素纤维在纳米尺度上呈平行排列,形成了纤维素纤维束和纤维。
纤维素的大分子结构非常有序,这种有序结构使纤维素具有很高的拉伸强度和模量。
纤维素纤维的强度和模量远远超过钢铁,因此纤维素具有很高的生物力学性能。
纤维素还具有超强的吸水能力,纤维素纤维能够吸收大量的水分,使其体积增大,并形成高度结晶的纤维素水胶体。
在纤维素纤维中,纤维素链之间通过氢键和范德华力相互作用。
这种相互作用使纤维素具有相对稳定的二级结构。
纤维素链通常以平行排列的方式组织在一起,形成纤维素纤维束和纤维。
纤维素的线性结构和氢键相互作用决定了纤维素的高度结晶性和热稳定性。
纤维素还具有很高的生物降解性和可再生性。
纤维素是植物细胞壁中的主要组分,它在自然界中被微生物和酶降解。
纤维素的降解产物是水和二氧化碳,没有任何有害的副产物。
这种生物降解性使纤维素成为一个非常重要的可再生材料,可以广泛应用于纺织、造纸、食品、医药等领域。
总结来说,纤维素的大分子结构是由若干个β-葡萄糖单体通过β-1,4-连接键连接而成的线性聚合物。
纤维素以微纤的形式存在于植物细胞壁中,并且形成了复杂的网状结构。
纤维素的有序结构使其具有很高的拉伸强度和模量,而其生物降解性和可再生性使其成为一个重要的可持续发展材料。
纳米纤维素的结晶度1纳米纤维素的简介纳米纤维素是一种从植物中提取的纳米级纤维素,其直径约为10-90纳米。
与传统的微米级纤维素相比,纳米纤维素具有更高的比表面积和更好的可溶性,在材料科学、生物医学、环境保护等领域具有广泛的应用前景。
2纳米纤维素的结晶度概念纳米纤维素的结晶度是指其纤维素晶体的数量和排列程度。
纤维素是构成植物细胞壁的一种主要成分,其晶体结构由许多葡萄糖分子构成。
随着纤维素晶体的增长,其晶体结构会逐渐排列成纤维素纤维,形成杆状结构。
3结晶度的影响因素纳米纤维素的结晶度受到多种因素的影响,包括原料来源、制备方法、溶液浓度、加热温度、离子强度等。
其中,原料来源和制备方法是影响纤维素晶体结构的主要因素。
不同的原料来源和制备方法会产生不同的晶体形态和结晶度,从而影响纳米纤维素的性质和应用。
4结晶度的测定方法目前,常用的纳米纤维素结晶度测定方法主要包括X射线衍射、红外光谱、偏振显微镜和热分析等。
其中,X射线衍射是最常用的方法之一。
X射线衍射可以通过测量衍射峰的位置和强度来确定纤维素晶体的结构和结晶度,但需要很高的仪器精度和样品制备技术。
5结晶度对纳米纤维素性质的影响结晶度是影响纳米纤维素结构和性质的重要参数之一。
纤维素晶体的结晶度越高,纳米纤维素的机械强度、热稳定性和抗水解性等性质就越好。
同时,高结晶度的纳米纤维素还具有更好的光学性能和可逆吸附性能,可用于光学材料和环境治理等领域。
6结晶度的调控随着纳米纤维素在各个领域中的应用越来越广泛,对其结晶度的调控也越来越重要。
目前,影响纳米纤维素结晶度的方法主要包括化学处理、物理处理和结构设计等。
化学处理是通过改变溶液成分和pH 值等来调控纤维素晶体结构和结晶度;物理处理是通过热处理、机械剪切等方式来促进晶体生长和调控结晶度;结构设计是通过构建有序和多级结构来调控纤维素纤维的结晶度和组织结构。
7结论纳米纤维素的结晶度是影响其性质和应用的重要参数之一。
随着技术的不断改进和深入研究,我们相信纳米纤维素的结晶度调控会带来更多的技术创新和应用突破。