Intraspecific responses in crop growth and yield of 20 soybean cultivars to enhanced
- 格式:pdf
- 大小:134.22 KB
- 文档页数:8

Effect of Biocyanol on Growth Performance of Grass CarpZHENG Xuebin 1,TAO Chunwei 2,WANG Anmin 3(1.College of Biotechnology,Hefei Vocational and Technical College,Chaohu 238000,Anhui China;2.Beijing Feedig Feed Sci &Tel Co.,Ltd.,Beijing 100085,China;3.Shanghai Xin Shangrun Biotechnology Co.,Ltd.,Shanghai 201605,China )Abstract:This study adopted the design of single factor experiments,was divided into four groups,each group ofthree repeat,including Ⅰgroup as the control group,the breeding area of 20000m 2,diet don 't add "net force",Ⅱ,Ⅲ,Ⅳgroup as experimental group,each group breeding area of 26667m 2,diet "net force","net force"150,200and 250mg ·kg -1.The average weight of grass carps used in the experiment was about 0.76g.The area of the fishpond used in the experiment was 100000m 2,and the feeding density of the four groups was the same,that is,grass carps with 6·tails ·m 2-1.The experimental results showed that:"the net force"can enhance the survival rate of the grass carp,which Ⅱ,Ⅲ,Ⅳgroup survival rate increased by 15.83%,23.82%and 24.33%respectively,and a significant difference (P <0.05);improve the specific growth rate of grass carp,which Ⅱ,Ⅲ,Ⅳgroup of specific growth rate were increased by 0.13,0.29,0.30%·d -1,and significant difference (P <0.05);reduce the bait coefficient of grass carp,which Ⅱ,Ⅲ,Ⅳgroup 0.09,0.16,0.17,bait coefficient,and a significant difference (P <0.05).The optimal addition level of "net force"in grass carp is 200g ·t -1.Key words:oregano;grass carp;growth performance牛至香酚(净力安)对草鱼生长性能的影响郑学斌1,陶春卫2,王安民3(1.合肥职业技术学院生物工程学院,安徽巢湖238000;2.北京菲迪饲料科技有限责任公司,北京100085;3.上海鑫上润生物科技有限公司,上海201605)收稿日期:2018-11-08基金项目:合肥职业技术学院食品专业综合改革试点项目(2016ZS001);安徽省省级质量工程项目(2016msgzs062)作者简介:郑学斌(1969-),男,安徽黄山人,硕士,副教授,畜牧师,主要从事动物营养、食品的教学和科研工作。



生态学&进化生物学词汇精选abundance多度(对物种个体数目多少的一种估测指标,多用于群落野外调查)abyssal zone深海带bathyl zone半深海带euphotic zone透光带pelagic zone浮游带,远洋带profundal zone深底带,深水层littoral zone沿岸带,滨海带 limnetic zone湖沼带,湖泊透光层intertidal zone潮间带acclimation驯化(指在实验条件下诱发的生理补偿机制,一般需时较短)acclimatisation驯化(指在自然环境条件下所诱发的生理补偿机制,通常需时较长)acquired character获得性状inheritance of acquired characters获得性状遗传adaptation适应(指生物的形态结构和生理机能与其赖以生存的一定环境条件相适应的现象)adaptive peak适应峰adaptive suite适应组合(生物对一组特定环境条件的适应必定会表现出彼此之间的相互关联性的协同适应特性)aerodynamic method空气动力法(用于测定初级生产量)aestivation夏眠 hibernation冬眠torpor 蛰伏(对恒温动物而言)diapause滞育(指某些昆虫的发育停滞) dormancy休眠age distribution年龄分布age pyramid年龄金字塔,年龄锥体age structure年龄结构increasing population增长型种群stable population稳定型种群declining population衰退型种群agglomerative method归并法agonistic behavior争斗行为altruism利他主义、利他行为(指一个个体牺牲自我而使社群整体或其他个体获得利益的行为)communication通讯行为courtship behavior求偶行为ingestive behavior索食行为parental behavior双亲行为social behavior社群行为territorial behavior领域行为atavism返祖现象atavistic regeneration返祖性再生allele等位基因allelopathy他感作用、克生作用(某些植物分泌一些有害物质,阻止别种植物在其周围生长的现象)allelopathic effects异种抑制效应,他感效应(指植物将其次生物质释放到周围环境中用以抑制别种生物的现象)allelopathic substances他感作用物质ammonification氨化作用(分解含氮有机物产生氨的生物学过程)mineralization矿化作用(微生物分解有机物质,释放CO2和无机物的过程)nitrification硝化作用(氨态氮被微生物氧化成亚硝酸,并进一步氧化成硝酸的过程)amphibian两栖动物reptile爬行动物mammal哺乳动物primate灵长类动物rodent啮齿动物arthropod节肢动物analogous organs同功器官homologous organs同源器官ancon sheep(短腿)安康羊Anthropoid类人猿anthropomorphism人神同性论(18~19世纪,即对社会和人类作神学的、形而上学的解释)antibiosis抗生(一种生物产生物质有害于一起生活的另一种生物)aposematic color警戒色(=warning coloration)protective coloration保护色mimicry拟态archeobacteria古细菌eubacteria真细菌Archeo-pteryx始祖鸟Ardipithecus ramidus始祖阿德猿、始祖南猿Australopithecus afarensis 阿法南猿Australopithecus南方古猿Homo habilis能人(“早期猿人”,早期人类第一阶段的代表)Homo erectus直立人(“晚期猿人或猿人”,早期人类第二阶段的代表)Homo sapiens智人(“化石智人”,早期人类第三阶段的代表,与现代人属同一个种)[人类起源发展的几个阶段]artificial ecosystem人工生态系统natural ecosystem自然生态系统technosphere工艺圈(指城市工业生态系统)agnosphere农艺圈(指农业生态系统)noosphere理智圈(Vernadsky,1945 定义为“人类按其意志、兴趣和利益而重新塑造的生物圈,即人类影响和控制的特殊生态系统”)association coefficient关联系数(表达种对之间是否关联。

植物逆境响应的分子机制植物作为一种生命体,在生长过程中难免会遭受各种不利条件的影响,比如土壤中的盐碱化、干旱、高温等环境因素。
对于植物本身而言,逆境的来临无异于是一场危机,即便是那些早已适应了恶劣环境的野生植物也无法完全摆脱逆境所带来的压力。
不过,植物也会在遭受逆境的时候出现以适应为目标的分子级反应,俗称逆境响应过程。
植物逆境响应的分子机制是什么呢?1.热休克蛋白热休克蛋白(Heat shock proteins,HSPs)是一种酶或者酶辅因子,在逆境环境下高表达,它们能够帮助植物维持正确的蛋白折叠状态、抑制错配和解聚。
逆境环境下,热休克蛋白对植物细胞的保护作用十分显著。
2.质膜脂质调节植物逆境响应过程中,受逆境条件刺激的质膜脂质会发生改变,进而导致细胞质壁的导电性能改变,或者是需要依赖于脂膜的酶等调节层级也会受到影响。
植物紧接着会启动各种反应,保证细胞膜的稳定性和完整性。
3.脱落酸脱落酸(Abscisic acid,ABA)是一种植物激素,起主要作用是抑制细胞分裂以及促进干旱等逆境环境下的植物适应。
在植物受到逆境刺激时,ABA的生产量会增加,依次来提高植物逆境抵抗能力。
4.氧化还原状态氧化还原状态同样是植物逆境响应过程的重要组成部分。
在植物细胞中,因应的代谢过程需要一定的氧化还原状态来进行调节。
在逆境环境下,产生大量有害的活性氧,会对植物细胞的氧化还原状态造成严重的影响。
植物逆境响应机制是复杂的,涉及到多种细胞层面的调节。
每一层面都有多个环节可以预测到的逆境,以便可能依次打击。
植物在反应逆境的过程中,有着复杂精细而高明的调控系统,最终保证了其在逆境环境中的生长。

处理小麦中多糖的工具—NIRS
Natasja Smeets;Filip Nuyens;王晶晶
【期刊名称】《国外畜牧学-猪与禽》
【年(卷),期】2013(033)003
【摘要】在欧洲,小麦因其淀粉和蛋白含量高而成为家禽日粮中一种主要的饲料成分。
然而,众所周知,小麦中的非淀粉多糖(Non-Starch Polysaccharides,NSP)对家禽有抗营养作用,此外,其浓度非常不定,而目前的饲料配方没有考虑该变异性。
本文将讨论NSP在肉鸡营养中的重要性,以及近红外光谱法(Near Infrared Reflectance Spectroscopy,NIRS)监测其浓度的可能性。
【总页数】2页(P27-28)
【作者】Natasja Smeets;Filip Nuyens;王晶晶
【作者单位】比利时Kemin EuropaNV公司;比利时Kemin EuropaNV公司【正文语种】中文
【中图分类】S816
【相关文献】
1.近红外技术(NIR)在小麦商品粮收购中的应用研究 [J], 胡新中;郭波莉;魏益民;张国权;欧阳韶晖
2.小波图像修补技术在小麦NIR图像处理中的应用 [J], 周光华;朱大洲;王成
3.酒精发酵处理对小麦非淀粉多糖的影响及酒糟在馒头中的应用 [J], 曹宇锋;赵仁勇
4.基于乙醇胺的深度共熔溶剂预处理小麦秸秆提高其多糖酶解效率 [J], 董艳梅;洪
元芳;马阳阳;安艳霞
5.一种从黑钨矿-白钨矿-锡石混合精矿中回收钨和锡的综合处理工艺:Sreeniras T.,et al.,Mineral Processiong &Extractive Metall.Rev [J], 陈隆玉
因版权原因,仅展示原文概要,查看原文内容请购买。

人类活动产生的废弃物中也含有大量的纤维素,如农业废物( 稻草、稻壳、麦杆、花生壳、玉米芯、棉籽壳、甘蔗渣等)、食品加工废物(果皮、果渣等)、木材废物(木屑、树皮)以及城市废弃物(40%~60% 固体废物自然界中细菌、真菌、某些无脊椎动物,直至高等植物中都有纤维素酶的存在
具有高比活力的动物性纤维素酶的研究随着对纤维素酶研究的不断深入,“动物自身不含纤维素酶”这一传统理论被推翻,动物纤维素酶成为纤维素酶研究的热点。
1998年,Smant等用分子生物学的方法,从两种不同种属的植物寄生的线虫中得到了4个内切B一1,4一葡聚糖酶(EG)的cDNA;证明了动物体内确实存在内源性的纤维素酶。
中国科学院上海生命科学研究院生物化学与细胞生物学研究所从福寿螺(Ampullafia crossean)的胃液中纯化到了一种分子量约41.5 kD具有葡聚糖内切酶、葡聚糖外切酶以及木聚糖酶等多种酶活力的多功能纤维素酶并命名为EGX.从福寿螺卵巢中克隆
由9个外显子和8个内含子组成的该基因组片段,这被认为是迄今为止在动物中发现的第一个具有潜在应用价值的多功能纤维素酶¨。
找到适合于工业生产的、高活性的纤维素酶是目前将纤维素物质转化为再生能源的关键点,动物性纤维素酶的研究为我们增添了一个解决问题的方向。
有报道指日本一家实验室从甲虫中得到一种葡聚糖内切酶水解羧甲基纤维素(CMC.Na)的比活力可高达150 IU/mg 。

植物逆境胁迫下的分子响应机制Introduction植物的长期生存离不开逆境环境中的谨慎应对,其中逆境胁迫是最常遇到的典型案例。
植物感知范围内的逆境胁迫,并启动分子层面的响应机制以抵抗挑战和尽可能地维持生长发育。
这篇文章将重点介绍植物逆境胁迫下的分子响应机制,其中包括植物模式识别受体、激素调控网络、次生代谢物品类、基因表达和蛋白质翻译调节等方面。
植物模式识别受体植物模式识别受体(PRRs)是植物防御响应的第一道屏障,它们可针对微生物致病分子通路中的共生分子(例如Flagellin,Chitin等)而产生响应,启动植物免疫机能。
由此可见,PRRs是植物维持自身稳态和适应不良环境的关键分子。
在逆境胁迫的背景下,PRRs可促进细胞外和细胞内的免疫反应。
例如,在病原体致病后,PRR调控局部酶催化,从而启动微生物降解机制。
在盐胁迫环境中,PRRs还可以介导盐胁迫适应过程中的肾上腺素和ATP信号转导,促进水分利用率和植物水分代谢能力的提高。
因此,PRRs是植物适应性的重要分子基础。
激素调控网络植物逆境胁迫中的激素调控网络是植物应对逆境的重要分子机制之一。
植物体内的激素调控网络包括乙烯(ethylene)、赤霉素(gibberellin)、脱落酸(abscisic acid)、生长素(auxin)和脯氨酸(proline)等,这些激素在不同的逆境环境中发挥着不同的生物学功能。
其中,乙烯和赤霉素能调节植物生长和发育,并在逆境胁迫后发挥重要作用。
例如,在盐胁迫环境中,乙烯可以促进盐胁迫适应的前期响应,从而增加植物的生物质生产和水分利用效率。
相反,在水胁迫环境中,赤霉素抑制植物根系的伸长,降低水分利用和土壤探测能力,并在逆境胁迫长期作用下,使植物处于萎蔫状态,失去活力。
次生代谢物品类植物紫外线、水胁迫和盐胁迫等逆境胁迫条件下,可激发植物次生代谢物品类的生产。
次生代谢物品类是由酚、醛、酮、生物碱、三萜类、酸类、内酯类等不同类型的化合物组成的,这些化合物在植物逆境响应中起到重要的生物学功能。
![拟南芥磷脂酶C基因及其在提高农作物抗热性中的应用[发明专利]](https://uimg.taocdn.com/f179d0117c1cfad6185fa77c.webp)
专利名称:拟南芥磷脂酶C基因及其在提高农作物抗热性中的应用
专利类型:发明专利
发明人:郑术芝,刘玉良,李冰,周人纲
申请号:CN201010581637.6
申请日:20101210
公开号:CN102121022A
公开日:
20110713
专利内容由知识产权出版社提供
摘要:本发明公开了拟南芥一种磷脂酶C基因及其应用,其特别是利用此基因提高植物的抗热性的方法。
本发明所提供的拟南芥的一种磷脂酶C蛋白基因,其核苷酸序列是序列表中序列1中的DNA序列;其所编码的氨基酸序列是序列表中序列2氨基酸残基序列。
实验证明,将上述基因在拟南芥中过量表达,可以提高植物在热激条件下的存活率和叶绿素含量,从而提高植株的耐热性。
本发明在利用分子育种提高农作物抗热上有重要作用。
申请人:河北师范大学
地址:050016 河北省石家庄市裕华东路113号
国籍:CN
代理机构:石家庄新世纪专利商标事务所有限公司
代理人:董金国
更多信息请下载全文后查看。
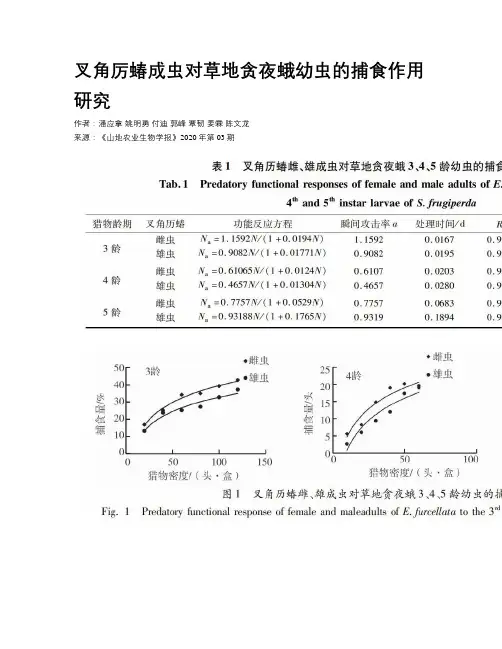
叉角厉蝽成虫对草地贪夜蛾幼虫的捕食作用研究作者:潘应拿姚明勇付迪郭峰覃韧姜霖陈文龙来源:《山地农业生物学报》2020年第03期摘要:叉角厉蝽是一种多食性的优势天敌,为系统评估叉角厉蝽对草地贪夜蛾的捕食能力,在室内条件下测试了叉角厉蝽雌、雄成虫对草地贪夜蛾3、4和5龄幼虫的捕食功能反应和种内干扰反应。
结果表明:叉角厉蝽雌、雄成虫对草地贪夜蛾3、4和5龄的捕食功能反应均符合HollingⅡ方程,雌成虫对草地贪夜蛾3、4和5龄捕食功能反应方程分别为Na=1.1592N/(1+0.0194N),Na=0.61065N/(1+0.0124N),Na=0.7757N/(1+0.0529N);雄成虫对草地贪夜蛾3、4和5龄捕食功能反应方程分别为Na=0.9082N/(1+0.01771N),Na=0.4657N/(1+0.01304N),Na=0.93188N/(1+0.1765N)。
叉角厉蝽雌、雄成虫对草地贪夜蛾龄幼虫的寻找效应随着猎物密度的增加而下降。
叉角厉蝽雌成虫对草地贪夜蛾的捕食作用率随着自身密度的增加而降低,其关系为E=0.3982P.-0.3409。
关键词:叉角历蝽;草地贪夜蛾;捕食作用;搜寻效应中图分类号:S433.4文献标识码:A文章编号:1008-0457(2020)03-0023-05國际DOI编码:10.15958/ki.sdnyswxb.2020.03.004Predation Effects of Eocanthecona furcellata Adults Against Spodoptera frugiperda LarvaePAN Yingna, YAO Mingyong, FU Di, GUO Feng, QIN Ren, JIANG Lin, CHEN Wenlong.*(Institute of Entomology, Provincial Key Laboratory for Agricultural Pest Management of Mountainous Region, Guizhou University, Guiyang, Guizhou 550025, China)Abstract:Eocanthecona furcellata is an omnivorous natural enemy. In order to systematically evaluate the predation ability of E. furcellata against Spodoptera frugiperda, predatory functional responses and intraspecific interference responses of female and male adults of E. furcellata on 3rd,4th and 5.th instar larvae of S. frugiperda were tested in the laboratory. The result showed that the response of female and male adults of E.furcellata to the predatory function of the 3.rd, 4.th and 5.th instars of S. frugiperda were fitted with HollingⅡequation. The predatory functional response equations of female E. furcellata to the predatory function of the 3.rd, 4.th and 5.th instars of S. frugiperda were Na=1.1592n/(1+0.0194n), Na=0.61065n/(1+0.0124n), Na=0.7757n/(1+0.0529n). The predatory functional response equations of male E. furcellata to the predatory function of the 3.rd, 4.th and 5.th instars of S. frugiperda were Na=0.9082N/(1+0.01771N),Na=0.4657N/(1+0.01304N),Na=0.93188N/(1+0.1765N). The search effect of female and male adults of E. furcellata on the larva of S. frugiperda decreased with the increasing of prey density. The predation rate of female adults of E. furcellata on the larva of S. frugiperda decreased with the increasing of density of S. frugiperda, and the relationship was E=0.3982P.-0.3409.Keywords:Eocanthecona furcellata; Spodoptera frugiperda;predation; searching efficiency草地贪夜蛾Spodoptera frugiperda,又称秋粘虫、行军虫,起源于美洲热带和亚热带地区,它是一种多食性的害虫,具有远距离迁飞能力,被联合国粮农组织称为“全球跨国界迁飞性的重大农业害虫”[1-2]。

刘满强,1975年8月生,山东人,教授,博士生导师。
1998年本科毕业于南京农业大学资环学院土壤与农业化学专业,2001年和2005年先后获得硕士和博士学位,毕业后留校。
近年来主要从事环境变化条件下土壤生物多样性、群落结构与生态功能关系的研究。
研究内容包括:1)干扰条件下土壤生物群落与功能的稳定性;2)陆地生态系统地上部和地下部亚系统的相互作用;3)人为措施对土壤食物网结构、生物相互作用和土壤生态系统服务功能的影响;4)不同时空尺度及高活性微域(界面)内土壤生物群落分布和碳氮转化;5)利用土壤生物资源促进土壤生物肥力及改善作物品质的研究。
2003年赴德国李比希大学动物生态研究所短期访问,参加DFG项目“Soil as sink and sources of CO2”的有机碳研究小组。
2005年底至2006年初赴英国苏格兰作物研究所(现The James Hutton Institute),进行土壤生物群落和功能稳定性方面的研究。
2009年初赴英国The James Hutton Institute进行植物地上部和地下部多营养级交互作用的学术交流。
2009至2010年在丹麦哥本哈根大学生物系陆地生态学研究所进行外来生物入侵及土壤食物网结构与植物生长关系的博士后研究。
2011年赴爱尔兰环境研究中心进行有关土壤动物-微生物相互作用对于土壤功能稳定性影响的研究。
主讲本科生《生态学》、《土壤生物与生态学》及研究生《土壤生态学》、《生态学研究进展》和《生态学研究方法》课程。
现为中国土壤学会会员,中国生态学会会员,国际土壤生态学会会员,江苏省生态学会副秘书长。
发表的主要论文有:[1]Song, X., Liu, M., Wu, D., Griffiths, B., Jiao, J., Li, H., Hu, F., 2015. Interaction matters: synergy betweenvermicompost and PGPR agents improves soil quality, crop quality and crop yield in the field. Applied Soil Ecology 89, 25–34(通讯作者)[2]Ma, C., Liu, M., Wang, H., Chen, C., Fan, W., Griffiths, B., Li, H., 2015. Resource utilization capability ofbacteria predicts their invasion potential in soil. Soil Biology and Biochemistry 81, 287–290(通讯作者)[3]Huang, J., Liu, M., Chen, X., Chen, J., Li, H., Hu, F., 2015. Effects of intraspecific variation in rice resistanceto aboveground herbivore, brown planthopper, and rice root nematodes on plant yield, labile pools of plant and rhizosphere soil. Biology and Fertility of Soils 51, 417-425(通讯作者)[4]Wu, D., Liu, M., Song, X., Jiao, J., Li, H., Hu, F., 2015. Earthworm ecosystem service and dis-service in anN-enriched agroecosystem: increase of plant production leads to no effects on yield-scaled N2O emissions.Soil Biology and Biochemistry 82, 1–8(通讯作者)[5]戚琳, 刘满强,蒋林惠, 张楗峤, 李修强, 陈法军, 胡锋. 2015. 基于根际与凋落物际评价转Bt水稻对土壤线虫群落的影响. 生态学报, 35 (5): 1434-1444(通讯作者)[6]王慧, 桂娟, 刘满强, 卢焱焱, 帕提古丽·亚生, 陈小云, 胡锋. 2015. 稻草和三叶草分解对微型土壤动物群落的影响. 土壤学报, 52(5): 1124-1134.(通讯作者)[7]郑加为, 陈法军, 刘满强, 赵宗潮, 范珍珍, 周诗竹, 胡锋. 2014. 转植酸酶玉米大田种植对根际土壤磷含量及组成的影响. 土壤学报, 51(5): 1110-1119.(通讯作者)[8]张腾昊, 王楠, 刘满强, 李方卉, 祝康利, 李辉信, 胡锋. 2014.秸秆、氮肥和食细菌线虫交互作用对土壤活性碳氮和温室气体排放的影响. 应用生态学报, 25 (11): 3307-3315.(通讯作者)[9]张微, 刘满强,何园球, 樊剑波, 陈晏. 2014. 长期施用不同无机肥对旱地红壤线虫群落的影响. 应用生态学报, 25 (8): 2361-2368.(通讯作者)[10]陈婧, 陈法军, 刘满强, 冯运, 党志浩, 李辉信, 胡锋. 2014. 温度和CO2浓度升高下转Bt水稻种植对土壤活性碳氮和线虫群落的短期影响. 生态学报, 34(6): 1481-1489.(通讯作者)[11]Song, X., Liu, M., Wu, D., Qi, L., Ye, C., Jiao, J., Hu, F., 2014. Heavy metal and nutrient changes duringvermicomposting animal manure spiked with mushroom residues. Waste Management 34, 1977-1983. (通讯作者)[12]Wu, Y., Jiang, Y., Jiao, J., Liu, M., Hu, F., Griffiths, B.S., Li, H., 2014. Adsorption of Trametes versicolorlaccase to soil iron and aluminum minerals: Enzyme activity, kinetics and stability studies. Colloids and Surfaces B-Biointerfaces 114, 342-348.[13]刘满强, 陈小云, 秦江涛, 黄欠如, 余喜初, 李辉信, 胡锋. 2013. 土壤团聚结构上水溶性有机物的性质及其对有机肥的响应. 中国农业科学, 46 (5), 961–969.[14]刘雨迪, 陈小云, 刘满强, 秦江涛, 李辉信, 胡锋. 2013. 不同稻作年限下土壤微生物学性质和线虫群落特征的变化. 生物多样性, 21 (3), 334–342.(通讯作者)[15]戚琳, 陈法军, 刘满强, 陈小云, 祝向钰, 李辉信, 胡锋. 2013. 三种转Bt水稻短期种植对土壤微生物生物量和线虫群落的影响. 生态学杂志, 32(4), 975–980.(通讯作者)[16]Huang, J., Liu, M., Chen, X., Chen, J., Chen, F., Li, H., Hu, F., 2013. Intermediate herbivory intensity of anaboveground pest promotes soil labile resources and microbial biomass via modifying rice growth. Plant and Soil, 367(1-2), 437–447.(通讯作者)[17]Yang, J., Li, X., Xu, L., Hu, F., Li, H., Liu, M., 2013. Influence of the nitrification inhibitor DMPP on thecommunity composition of ammonia-oxidizing bacteria at microsites with increasing distance from the fertilizer zone. Biology and Fertility of Soils, 49(1), 23–30 (通讯作者)[18]Huang, J., Liu, M., Chen, F., Griffiths, B., Chen, X., Johnson, S., Hu, F., 2012. Crop resistance traits modifythe effects of an above-ground herbivore, brown planthopper, on soil microbial biomass and nematode community via changes to plant performance. Soil Biology and Biochemistry 49, 157–166 (通讯作者). [19]Liu, M., Chen, X., Griffiths, B.S., Huang, Q., Li, H., Hu, F., 2012. Dynamics of nematode assemblages andsoil function in adjacent restored and degraded soils following disturbance. European Journal of Soil Biology 49, 37–46.[20]Bjørnlund, L., Liu, M., Rønn, R., Christensen, S., Ekelund, F., 2012. Nematodes and protozoa affect plantsdifferently, depending on soil nutrient status. European Journal of Soil Biology 50, 28–31.[21]Liu, M., Bjørnlund, L., Rønn, R., Christensen, S., Ekelund, F., 2012. Disturbance promotes non-indigenousbacterial invasion in soil microcosms: analysis of the roles of resource availability and community structure.PLoS ONE 7, e45306.[22]李修强, 陈法军, 刘满强, 胡锋. 2012. 转Bt水稻对土壤可溶性有机碳氮及微生物学性质的影响. 应用生态学报 23, 96–102(通讯作者).[23]Liu, M., Chen, X., Chen, S., Li, H., Hu, F., 2011. Resource, biological community and soil functional stabilitydynamics at the soil-litter interface. Acta Ecologica Sinica 31, 347–352.[24]陈小云, 郭菊花, 刘满强, 焦加国, 黄欠如, 赖涛, 李辉信, 胡锋. 2011. 施肥对红壤性水稻土有机碳活性和难降解性组分的影响. 土壤学报48, 125–131(通讯作者).[25]Li, D., Liu, M., Cheng, Y., Wang, D., Qin, J., Jiao, J., Li, H., Hu, F., 2011. Methane emissions fromdouble-rice cropping system under conventional and no tillage in southeast China. Soil & Tillage Research 113, 77–81.[26]汤英,刘满强,王峰,陈法军,邵波,苏昱,葛成,黄菁华,李辉信,胡锋. 2010. 褐飞虱对水稻苗期生长及地下部土壤活性碳氮的影响. 生态学报30, 2890–2898(通讯作者)[27]刘满强,黄菁华,陈小云,王峰,葛成,苏昱,邵波,汤英,李辉信,胡锋. 2009. 地上部植食者褐飞虱对不同水稻品种土壤线虫群落的影响. 生物多样性17, 431–439.[28]Liu, M., Hu, F., Chen, X., Huang, Q., Jiao, J., Zhang, B., Li, H. 2009. Organic amendments with reducedchemical fertilizer promote soil microbial development and nutrient availability in a subtropical paddy field: the influence of quantity, type and application time of organic amendments. Applied Soil Ecology 42, 166–175.[29]陈石,陈小云,李辉信,胡锋,刘满强. 2009. 食真菌线虫对热或铜胁迫下土壤生态功能稳定性的影响.应用生态学报20, 435-440(通讯作者).[30]Tao, J., Chen, X., Liu, M., Hu, F., Griffiths, B., Li, H. 2009. Earthworms change the abundance andcommunity structure of nematodes and protozoa in a maize residue amended rice-wheat rotation agro-ecosystem. Soil Biology and Biochemistry 41, 898–904.[31]Liu, M., Chen, X., Qin, J., Wang, D., Griffiths, B., Hu, F. 2008. A sequential extraction procedure reveals thatwater management affects soil nematode communities in paddy fields. Applied Soil Ecology 40, 250–259. [32]Mao, X., Hu, F., Griffiths, B., Chen, X., Liu, M., Li, H., 2007. Do bacterial-feeding nematodes stimulate rootproliferation through hormonal effects? Soil Biology & Biochemistry 39, 1816-1819.[33]刘满强, 陈小云, 郭菊花, 李辉信, 胡锋. 2007. 土壤生物对土壤有机碳稳定性的影响. 地球科学进展22, 152–158.[34]刘满强, 胡锋, 陈小云. 2007. 土壤有机碳稳定机制研究进展. 生态学报27, 2642–2650.[35]Ekschmitt, K., Liu, M.,Vetter, S., Fox, O., Wolters, V. 2005. Strategies used by soil biota to overcome soilorganic matter stability-Why is dead organic matter left over in the soil? Geoderma 128, 167–176.[36]刘满强, 胡锋, 陈小云, 何圆球. 2004. 退化红壤不同植被恢复方式对蚯蚓种群的影响. 应用生态学报15, 2152–2156.[37]刘满强, 胡锋, 何圆球, 李辉信. 2003. 退化红壤不同植被恢复下土壤微生物量季节动态及其指示意义.土壤学报 40, 937–944.[38]刘满强, 胡锋, 李辉信, 陈小云, 何圆球. 2002. 退化红壤不同植被恢复下土壤节肢动物群落特征. 生态学报 22, 54–61.参加或主持的主要科研项目:[1]国家自然科学基金面上项目“蚯蚓对农田土壤碳氮转化、平衡及作物生产力的影响”(2004-2006);[2]国家自然科学基金青年基金“水稻土有机碳的生物稳定机制及影响因素”(2006-2008);[3]教育部博士点专项基金“蚯蚓产生的可溶性有机物对重金属污染土壤植物修复效率的影响”(2008-2010);[4]国家科技支撑计划“退化红壤肥力重建及生态功能定向培育技术研究”(2009-2011);[5]转基因生物新品种培育科技重大专项“基于土壤动物及其生物标志物的转基因作物检测与安全性监测新技术”子课题(2009-2011);[6]国家重点基础研究发展计划(973)子专题“新型硝化抑制剂/脲酶抑制剂对肥际微域氮素转化和损失的影响”(2007-2011);[7]国家自然科学基金重点项目“稻田土壤有机碳固定与稳定化过程及机制:土壤-作物-微生物相互作用研究”子专题(2009-2012);[8]农业部公益性行业科研专项“农业清洁生产与农村废弃物循环利用集成配套技术体系研究与示范”(2010-2013);[9]国家自然科学基金“水稻地上和地下部植食者的相互关系、机制及调控研究”(2012-2015)。
20/881环球农化 富美实近日推出了新款产品Presence,是巴西市场上首个获准用于种子的微生物杀线虫剂。
Presence 是溶解于悬浮液的水分散性粉剂,该剂型不会增加药液体积。
产品具有绝佳的稳定性,是市场上浓度最高的产品,并且能与目前所有用作种子处理剂的化学产品兼容。
与化学产品相比,这一新款微生物杀线虫剂在用作种子处理剂时重点防治植物寄生线虫。
该产品已获得登记,而通常在市场上见到标签注明为杀线虫剂的产品并没有获得此类用途的登记。
该产品含有枯草芽孢杆菌和地衣芽孢杆菌两种细菌,其独有的特点在于这些细菌的浓度,但比浓度更重要的是每克产品与植物共生的数量。
Presence 中活性成分的浓度为2×1011,比市场上的同类产品高200倍。
富美实推出微生物杀线虫剂 Presence 具备多重作用机制,例如通过形成物理障碍保护作物。
在种子开始萌芽时,产品中将要繁殖并寄生于植物根系的两类细菌会形成一层起屏障作用的生物膜。
这两种寄生于根部的细菌会与植物产生协同作用,在消耗根部产生的分泌物后,它们会发挥抵御根部寄生线虫的作用机制。
在这些根部分泌物浓度较低时,线虫将会难以发现根部。
同时,这些细菌还会分泌几丁质酶,该物质可杀死线虫的幼虫。
Presence 的保质期长达24个月,产品登记用于防治南方根结线虫和最短尾短体线虫。
由于这是一款生物制品,产品因其用于靶标线虫的功能而受到监管,而非其针对作物的功效。
Presence 可用于所有受到这些线虫侵害的作物,尤其是棉花、玉米和豆类作物。
巴西农业、畜牧业和食品供应部(Mapa)将很快宣布批准登记含氟吡菌酰胺的新产品。
氟吡菌酰胺是一种高效化合物,已获得欧洲、美国和澳大利亚监管机构的使用批准,可防治灰霉病、白粉病、斑点落叶病、菌核病和念珠菌病等病害。
在巴西进行的研究还表明,它能有效防治根腐线虫(Pratylenchus brachyurus)。
农业部农药及相关产品总协调员Carlos Venâncio 介绍说:“线虫通常不易看到,但对农业造成的损害却相当大,而该产品是控制线虫的新选择,且与市场上的在售产品相比该产品的毒性更小。
急性胰腺炎患者肠道菌群变化及益生菌的应用吕彦青,李嘉杰,刘坤禹,郭汝华,季慧范吉林大学白求恩第一医院肝胆胰内科,长春 130021通信作者:季慧范,************.cn(ORCID: 0000-0001-8816-1906)摘要:肠道菌群是人体中最复杂和最重要的微生态系统,肠道菌群失调与急性胰腺炎的发生、发展密切相关,近年来靶向调控肠道微生态辅助治疗急性胰腺炎逐步被人们重视。
本文详细叙述了急性胰腺炎患者肠道菌群变化情况及机制,总结了益生菌应用的研究现状,指出益生菌辅助治疗方案的研究方向,并提出了预测急性胰腺炎患者优势菌群的新方法,以期为急性胰腺炎的治疗带来新思路。
关键词:胰腺炎,急性坏死性;胃肠道微生物组;鼠李糖乳杆菌基金项目:吉林市人口与健康发展研究(371170143428)Changes in intestinal microbiota and application of probiotics in patients with acute pancreatitisLYU Yanqing, LI Jiajie, LIU Kunyu, GUO Ruhua, JI Huifan.(Department of Hepatobiliary and Pancreatic Medicine, The First Hospital of Jilin University, Changchun 130021, China)Corresponding author: JI Huifan,************.cn(ORCID: 0000-0001-8816-1906)Abstract:Intestinal microbiota is the most complex and important microecosystem in the human body, and gut microbiota dysbiosis is closely associated with the development and progression of acute pancreatitis. Targeted regulation of intestinal microecology in assisting the treatment of acute pancreatitis has attracted more attention in recent years. This article describes the changes in intestinal microbiota and related mechanisms in patients with acute pancreatitis,summarizes the current research status of the use of probiotics, points out the research direction of probiotics as the adjuvant treatment regime, and proposes a new method for predicting the dominant flora in patients with acute pancreatitis, in order to bring new ideas for the treatment of acute pancreatitis.Key words:Pancreatitis, Acute Necrotizing; Gastrointestinal Microbiome; Lactobacillus RhamnosusResearch funding:Research on Population and Health Development of Jilin City (371170143428)急性胰腺炎(acute pancreatitis,AP)是多种病因导致胰腺组织自身消化所致的胰腺水肿、出血及坏死等炎症性损伤,组织学上以腺泡细胞破坏为特征。
植物富含半胱氨酸的类受体激酶的研究进展郑超;李登高;白薇【摘要】Receptor-like kinases(RLKs)are protein kinases existing in plants,and play an essential role in many plant signal transduction pathways. As receptors localized on plasma membrane,RLKs perceive environmental stimuli and are involved in intercellular signal transductions through phosphorylation. Cysteine-rich receptor-like kinases(CRKs),also called domain of unknown function 26(DUF26) receptor-like kinases,are a large subgroup of RLKs. Recently,CRKs are found to be involved in plant disease resistant defenses. In this paper,we reviewed the structural characters of CRKs and summarized biological functions in abiotic and biotic stresses and their roles in plant growth anddevelopment,moreover,we prospect the future research,aim to follow-up stuclies.%类受体激酶(receptor-like kinase,RLK)是植物体内普遍存在的一类蛋白激酶,是许多信号识别传递途径中的关键组分,作为识别信号的质膜受体,能够感受外界刺激,通过磷酸化作用参与胞内信号传递。
Green food,a term that has become increasingly popular in recent years,refers to food products that are produced in accordance with specific standards and are free from harmful substances such as chemical pesticides,fertilizers,and growth hormones.These products are considered to be beneficial for health and the environment.Here are some key points to consider when discussing green food in an English essay:1.Definition and Importance:Begin by defining what green food is and why it is important.Mention that it is food produced without the use of harmful chemicals,which makes it safer and healthier for consumption.2.Health Benefits:Discuss the health benefits of consuming green food.This could include reduced exposure to harmful chemicals,better nutrient content,and the potential for improved overall health.3.Environmental Impact:Highlight the positive environmental effects of green food production.This includes less pollution from chemical runoff,healthier soil,and support for biodiversity.4.Production Standards:Explain the standards that green food must meet.This could involve organic farming practices,sustainable agriculture,and the avoidance of genetically modified organisms GMOs.5.Certification and Labeling:Talk about the certification process that green food undergoes to ensure its quality and authenticity.Discuss the significance of labels that consumers can look for to identify green food products.6.Challenges and Solutions:Address the challenges faced in the production and distribution of green food,such as higher costs and limited availability.Suggest solutions like government support,consumer education,and technological advancements to improve efficiency.7.Consumer Behavior:Discuss how consumer behavior can influence the demand for green food.Mention the role of awareness and the willingness to pay a premium for healthier and environmentally friendly products.8.Future Outlook:Conclude with an optimistic view of the future of green food.Discuss the potential for increased adoption of green food practices and the benefits this could bring to both individuals and the planet.9.Personal Perspective:Add a personal touch by sharing your own experiences oropinions on green food.This could include your reasons for choosing green food or any changes youve noticed since incorporating it into your diet.10.Call to Action:End the essay with a call to action,encouraging readers to consider the benefits of green food and to make informed choices when shopping for groceries.Remember to use clear and concise language,provide examples where appropriate,and maintain a logical flow throughout your essay.This will help to engage your readers and effectively communicate the importance of green food.。
The future of agriculture is poised for significant changes that will revolutionize the way we grow,harvest,and distribute food.As the global population continues to grow, the need for sustainable and efficient agricultural practices becomes increasingly important.Here are some key areas where we can expect to see major transformations in the coming years:1.Technological Advancements:The integration of technology in agriculture is set to increase exponentially.This includes the use of drones for crop monitoring,precision farming tools that optimize the use of water and fertilizers,and data analytics to predict crop yields and manage resources more effectively.2.Sustainable Practices:With climate change being a pressing issue,sustainable farming practices will become more prevalent.This includes organic farming,regenerative agriculture,and the use of renewable energy sources to power agricultural operations.3.Vertical Farming:Urbanization is leading to a decrease in available farmland.Vertical farming,which involves growing crops in vertically stacked layers,will allow for the efficient use of space and reduce the need for arable land.4.Genetic Modification:Advances in genetic engineering will enable the development of crops that are more resistant to pests,diseases,and changing climate conditions.This could lead to higher yields and more resilient food sources.5.Agricultural Robotics:Robots will play a larger role in agriculture,performing tasks such as planting,harvesting,and weeding.This will not only increase efficiency but also reduce the need for manual labor.6.Aquaculture and Insect Farming:As traditional livestock farming faces challenges related to environmental impact and resource use,alternative protein sources like aquaculture fish farming and insect farming will gain popularity.7.Precision Livestock Farming:Similar to precision farming for crops,precision livestock farming will use technology to monitor the health and wellbeing of animals, ensuring optimal growth conditions and reducing the need for antibiotics.8.Food Waste Reduction:Innovations in storage,transportation,and packaging will help to minimize food waste.This includes the development of new materials that prolong the shelf life of produce and the use of smart systems that track and manage inventory.9.Blockchain in Agriculture:The use of blockchain technology will improve traceabilityand transparency in the food supply chain,allowing consumers to know the origin of their food and ensuring quality and safety standards are met.10.ClimateSmart Agriculture:Farmers will adopt practices that help to mitigate the effects of climate change,such as carbon sequestration in soils,water conservation techniques,and the use of cover crops to improve soil health.cation and Training:There will be a greater emphasis on educating farmers about new technologies and practices.This will involve partnerships between educational institutions,government agencies,and the agricultural industry.12.Policy and Regulation:Governments will play a crucial role in shaping the future of agriculture through policies that encourage innovation,support sustainable practices,and ensure food security.In conclusion,the future of agriculture is one that will be characterized by innovation, sustainability,and a focus on meeting the needs of a growing global population.By embracing these changes,we can work towards a more efficient and environmentally friendly food system.。
Intraspecific responses in crop growth and yield of 20soybean cultivars to enhanced ultraviolet-B radiation under field conditionsLi Yuan a,b,*,Zu Yanqun b ,Chen Jianjun b ,Chen Haiyan baThe Center for Agricultural Biodiversity Research and Training of Yunnan Province,Yunnan Agricultural University,Kunming 6502011,PR ChinabEco-environment Research Institute,College of Resources and Environment,Yunnan Agricultural University,Kunming 6502011,PR ChinaReceived 22October 2001;received in revised form 15April 2002;accepted 18April 2002AbstractField studies were conducted to determine the potential for intraspecific responses in crop growth and grain yield of 20soybean cultivars to enhanced ultraviolet-B (UV-B,280–315nm)radiation.The supplemental UV-B radiation was 5.00kJ m À2,simulating a depletion of 20%stratospheric ozone at Kunming (258N,1950m).Out of the 20soybean cultivars tested,17and 15showed significant change in plant height at 80DAP (days after planting)and ripening stages,respectively.Sensitivity in plant height was greater at 80DAP than at ripening.The plant height of 3cultivars increased,and that of 17cultivars decreased.Under UV-B radiation,LAI (leaf area index),biomass and grain yield decreased,respectively.The greatest percent decrease was 95.7,93.9and 92.8,respectively.RI (response index)was the sum of percent change in plant height at ripening,LAI,biomass and grain yield.The results showed that all 20soybean cultivars had a negative RI,indicating inhibition by UV-B radiation on soybean growth.The RI of 6tolerant cultivars was higher than À163.1and 5out of 6originated from south China (low latitude).The RI of the most tolerant cultivars,Yunnan 97801,was À72.4.Meanwhile,the RI of 5sensitive cultivars was lower than À256.9and 4out of the 5originated from north China (high latitude).The RI of the most sensitive cultivar,Huanxianhuangdou,was À295.7.These UV-B tolerant cultivars identified in this study might be useful in breeding programs.#2002Elsevier Science B.V .All rights reserved.Keywords:Soybean;Stratospheric ozone depletion;UV-B radiation;Response index;Crop growth;Grain yield1.IntroductionThe rapid decline in stratospheric ozone concentra-tions has been confirmed by satellite measurements (Molina and Molina,1992).The most pronounced thinning of the ozone layer has been measured overthe Antarctic continent with up to 71%depletion during the Antarctic spring (Kerr,1993).Recent mathematical models predict a further increase in solar ultraviolet-B (UV-B)irradiation in future years (Madronich et al.,1995).UV-B effects on plants have been the subject of considerable research;approxi-mately 600papers have been published (Caldwell et al.,1998).An examination of more than 200plant species reveals that roughly 20%are sensitive,50%are mildly sensitive or tolerant and 30%are comple-tely insensitive to UV-B radiation (Teramura,1983).Field Crops Research 78(2002)1–8*Corresponding author.Present address:College of Resources and Environment,Yunnan Agricultural University,Kunming 6502011,PR China.0378-4290/02/$–see front matter #2002Elsevier Science B.V .All rights reserved.PII:S 0378-4290(02)00084-9Whilst the impact of enhanced UV-B radiation on plant physiology,morphology,growth,nutrition and biomass has been investigated extensively(Li et al., 1998,1999;Yue et al.,1988),little is known about intraspecific responses of plants to enhanced UV-B. Intraspecific responses in maize and wheat(Biggs and Kossuth,1978;Li et al.,2000a,b),broad bean (Bennett,1981),cucumber(Murali and Teramura, 1986),soybean(Teramura and Murali,1986;Sullivan and Teramura,1990;Teramura et al.,1990a;D’surney et al.,1993),rice(Teramura et al.,1991;Barnes et al., 1993;Dai et al.,1994)have been reported.Plant species and even genotypes within species can differ greatly in their responses to UV-B.Reasons for this are not clear (Caldwell and Flint,1994).Due to our lack of under-standing of the basis of intraspecific responses to UV-B radiation,further studies on its importance would be worthwhile.Most of the UV-B research in the past two decades has been conducted as short-term experiments in growth chambers and greenhouses,where an unna-tural spectral balance of radiation may have substan-tially changed plant sensitivity to UV-B.It is important in experiments to maintain a realistic bal-ance between various spectral regions,since both UV-A(315–400nm)and visible(400–700nm)radiation can have ameliorating effects on responses of plants to UV-B(Caldwell et al.,1995).In growth chamber and greenhouse experiments,the visible and UV-A radiation is usually much less than in sunlight;thus, even if realistic levels of UV-B are used in simulating ozone reduction,the plant response may be exagger-ated relative tofield conditions.Unfortunately,only 15%of the studies have been conducted underfield conditions.While the laboratory and glasshouse stu-dies provide information on mechanisms and pro-cesses of UV-B action,onlyfield studies can provide realistic assessments of what will happen as the stratospheric ozone layer thins(Caldwell et al., 1995).Vegetation communities function differently to iso-lated plants and the behavior of the former is not easily predicted from the latter.Besides,the responses of soybean crops to UV-B are more complicated than those of plants isolated(Teramura and Murali,1986; Sullivan and Teramura,1990;Teramura et al., 1990a,b;D’surney et al.,1993).Community-level field experimentation is needed to evaluate realistic consequences of increased solar UV-B resulting from ozone reduction.Soybean is one of the major world food crops.The effects of enhanced UV-B radiation on photosynthe-tic characteristics,growth,morphology,intraspecific responses,total biomass and yield have already been studied(Sullivan and Teramura,1990;Teramura, 1980,1983;Teramura and Murali,1986;Teramura et al.,1990a,b;Lydon et al.,1980;D’surney et al., 1993).Unfortunately,only few studies have been conducted underfield conditions(Teramura et al., 1990a).In this study we grew20soybean cultivars in thefield under ambient and supplemental levels of UV-B radiation for a season with the objective of evaluating intraspecific responses in crop growth and yield.We hypothesized that enhanced UV-B radiation would affect crop growth and yield and result in intraspecific responses underfield condi-tions.2.Materials and methods2.1.Plant materials and growth conditionsThefield experiment was conducted at Yunnan Agricultural University,Kunming,China.No fertili-zation was needed during the season.Seeds of20 soybean(Glycine max)cultivars were obtained from Yunnan Academic of Agricultural Sciences,Gansu Academic of Agricultural Sciences and Henan Aca-demic of Agricultural Sciences.Seeds were sown in rows spaced0.4m apart at a density of50seeds mÀ2in 120plots of2:0Â1:0m2each on10April1999.Five border rows were sown round each plot in order to minimize heterogeneity in microclimate.The overall experimental design was a randomized complete block with two UV-B treatments and three replica-tions.At30DAP(days after planting),plants were thinned to40mÀ2for uniformity in growth.This planting density is within commonly used sowing practices for the Kunming region.2.2.UV-B radiationSupplemental UV-B radiation was provided by filtered Gucun brand30W sunlamps(Gucun Instru-ment Factory,Shanghai,China)following the2L.Yuan et al./Field Crops Research78(2002)1–8procedure outlined in Lydon et al.(1986).Lamps were suspended above and perpendicular to the planted rows(rows oriented in an east–west direction to minimize shading)andfiltered with either0.13mm thick cellulose diacetate(transmission down to 290nm)for supplemental UV-B radiation or 0.13mm polyester plasticfilms(absorbs all radiation below320nm)as a control(Sullivan and Teramura, 1990).Cellulose diacetatefilters were presolarized for 8h and changed weekly to ensure uniformity of UV-B transmission.The spectral irradiance from the lamps was determined with an Optronics Model742(Optro-nics Laboratories,Orlando,FL,USA)spectroradi-ometer.The spectral irradiance was weighted with the generalized plant response action spectrum(Cald-well,1971)and normalized at300nm to obtain UV-B BE.Six lamps were installed above each plot.Plants were irradiated for7h daily from30DAP to ripening stage,centered around solar noon.Plants under polye-ster-filtered lamps received only ambient levels of UV-B radiation(10.00kJ mÀ2UV-B BE during clear sky conditions on the summer solstice).Plants beneath the cellulose diacetatefilters received ambient plus sup-plemental levels of UV-B.The lamp height above the plants was adjusted weekly to maintain a distance of 0.45m between the lamps and the top of the canopy, and provided supplemental irradiances of5.00effec-tive kJ mÀ2UV-B BE.This supplemental level was similar to that which would be experienced at Kunm-ing(258N,1950m)with a20%stratospheric ozone reduction during a clear day on the summer solstice (10.00kJ mÀ2UV-B BE)according to a mathematical model of Madronich et al.(1995).Total daily photo-synthetic photonfluence(PPF between400and 700nm)under lampfixtures was90%of that above the lamps.2.3.Measurements and statistical analysesFifteen plants per plot were used to observe plant height measured as the distance from the soil surface to shoot tip at80DAP and ripening.Plants in two subplots of0:5Â0:5m2each were harvested from each plot to determine leaf area index (LAI)at55DAP,total biomass(including roots)per subplot and grain yield per subplot at ripening.Total leaves and a subsample of15leaves per subplot were collected at55DAP,and LA(area per leaf)of this subsample was measured with a Li-Cor3100(Li-Cor, Lincoln,NE,USA)area meter.All leaves were then oven-dried at688C for68h and weighed.A regres-sion relationship was developed between leaf weight and leaf areaðP<0:01Þ.This linear regression was used to determine total LA per subplot,then LAI was I is leaf area per unit ground area. Total biomass and grain were oven-dried at688C for 68h and weighed.The response index(RI)according to Dai et al. (1994),Teramura and Murali(1986)and Li et al. (2000a)used to evaluate the overall response of soybean to enhanced UV-B radiation which was cal-culated as follows:RI¼PHtÀPH cPH cþLAI tÀLAI cLAI cþTB tÀTB cTB cþGY tÀGY cGY cÂ100%where RI is the response index,PH the plant height at ripening stage,LAI the leaf area index,TB the total biomass and GY the grain yield under t(UV-B radia-tion)and c(control).Statistical differences between means of control and UV-B radiation treatment of any measured parameter were determined by t-test at the P<0:05or0.01level. Correlation between the percent change of different parameters were determined at the P<0:05or0.01 level.3.Results3.1.Plant heightUV-B radiation had obvious effects on the plant height of20soybean cultivars at80DAP and ripening underfield conditions(Table1).At80DAP,UV-B radiation had a positive effect on Yunnan97929ðP<0:05Þ,and a negative effect on16cultivars (P<0:05or0.01).At ripening,UV-B radiation increased the height of Lanyin20ðP<0:05Þ,and significantly decreased the height of14cultivars (P<0:05or0.01).Out of20cultivars,17and15 showed significant differences(P<0:01or0.05)at 80DAP and at ripening,respectively.Sensitivity in plant height of20soybean cultivars to enhanced UV-BL.Yuan et al./Field Crops Research78(2002)1–83radiation varied with time,being greater at80DAP than at ripening stage.3.2.Leaf area indexUnderfield conditions at55DAP,LAI of20 soybean cultivars decreased significantly by UV-B radiation(Table2).Of20cultivars,Yunnan97801 and Yunnan97501showed significance at P<0:05 while another18cultivars were significant at P<0:01.UV-B radiation did not have positive effects on LAI for any of the cultivars.3.3.Total biomassUV-B radiation significantly decreased total bio-mass of20soybean cultivars(Table2).Of the20 cultivars,Yunnan97801,Yunnan97929and Yudou10 were significant at P<0:05,while another17culti-vars were significant at P<0:01.UV-B radiation did not have positive effects on total biomass for any of the cultivars.3.4.Grain yieldThe effect of UV-B radiation on grain yield of20 soybean cultivars underfield conditions is presented in Table2.Of the20cultivars,UV-B radiation had no significant effect on Lanyin20,Yudou8,Lvgundou, Heidadou and Longdou1,and had a negative effect on the other15cultivars(P<0:05or0.01).3.5.Response indexThe RI is an integration of the effect on plant height, LAI,total biomass and grain yield,which could reflect the overall sensitivity of soybean cultivars to enhanced UV-B radiation.In this case,all20cultivars had a negative RI,indicating an overall inhibition of UV-B radiation on soybean growth(Table3).Huanxian-huangdou was most adversely affected(RI,À295.7),while Yunnan97801was least affected (RI,À72.4).Across all cultivars tested in the present study,five cultivars had negative RI less thanÀ256.9, the most sensitive cultivars were Huanxianhuangdou,Table1Intraspecific sensitivity to UV-B radiation based on plant height(cm)of20soybean cultivars underfield conditions aCultivar name80DAP Ripening stageControlþUV-B%Change ControlþUV-B%Change Yunnan9780141.237.7À8.546.846.3À1.1 Lanyin2021.522.3 3.718.723.324.6* Yunnan9792943.349.514.3*57.058.8 3.2 Yudou1046.441.1À11.4*58.646.4À20.8** Yudou1854.246.4À14.4**61.948.1À22.3** Yudou849.540.3À18.6**58.342.2À27.6** Lvgundou34.934.8À0.349.949.8À0.2 Heidadou85.265.8À22.8**88.866.3À25.3* Yunnan9750162.348.1À22.8**68.247.6À30.2** Pitiaohuangdou64.250.9À18.4**60.451.9À14.1* Yunnan9750645.236.8À18.6**46.939.1À16.6** Yunnan9770153.843.6À19.0**49.043.4À11.4 Longdou176.646.4À39.4**79.852.4À34.3** E013857.140.4À29.3**68.452.3À23.5** Yunnan9651052.538.2À27.2**53.039.4À25.7** Df-159.445.7À23.1**72.144.9À37.7** Lingtaihuangdou39.527.2À31.1**39.732.4À18.4** Kanglehuangdou33.929.6À12.7*34.336.7 6.7 Tuhuangdou166.449.5À25.5**73.640.5À45.0** Huanxianhuangdou37.326.8À28.2**37.431.0À17.1**a Measurements were made on15plants per plot.*Significant differences between control and UV-B radiation at P<0:05according to t-test.**Significant differences between control and UV-B radiation at P<0:01according to t-test.4L.Yuan et al./Field Crops Research78(2002)1–8L.Yuan et al./Field Crops Research78(2002)1–85 Table2Intraspecific sensitivity to UV-B radiation based on LAI,total biomass and grain yield of20wheat cultivars underfield conditions a Cultivar name LAI Total biomass(g mÀ2)Grain yield(g mÀ2)ControlþUV-B%Change ControlþUV-B%Change ControlþUV-B%Change Yunnan97801 1.2870.976À24.2*396.8312.2À21.3*98.973.4À25.8* Lanyin20 1.7090.285À83.3**201.2105.0À47.8**68.556.5À17.5 Yunnan97929 3.455 1.078À68.8**617.5357.1À42.2*133.0105.5À20.7* Yudou10 2.982 1.057À64.5**446.2282.2À36.8*42.233.0À21.8* Yudou18 1.8500.735À60.3**412.4193.8À53.0**36.626.8À26.8* Yudou8 3.290 1.181À68.9**474.2237.6À49.9**45.537.9À16.7 Lvgundou 3.8240.390À89.8**698.5255.5À63.4**82.368.9À16.3 Heidadou 2.2040.551À75.0**615.3272.5À55.7**130.7109.4À16.3 Yunnan97501 1.655 1.069À35.4*833.3185.5À77.7**36.322.5À38.0* Pitiaohuangdou0.5430.269À50.5**283.569.7À75.4**33.817.0À49.7** Yunnan97506 1.8790.621À67.0**593.3294.4À50.4**161.564.3À60.2** Yunnan97701 1.9320.734À62.0**416.9122.1À70.7**39.116.9À56.8** Longdou1 5.4780.238À95.7**1042.4277.4À73.4**142.6125.9À11.7E0138 1.9510.330À83.1**674.4155.0À77.0**50.830.6À39.8* Yunnan96510 2.0820.753À63.8**392.259.6À84.8**10.4 5.2À50.0** Df-1 2.7880.413À85.2**971.6254.8À73.8**195.577.8À60.2** Lingtaihuangdou 2.8730.156À94.6**387.445.3À88.3**42.516.0À62.4** Kanglehuangdou 1.3510.106À92.2**343.446.9À86.3**92.8 6.7À92.8** Tuhuangdou1 1.4550.254À82.5**326.166.3À79.7**105.317.5À83.4** Huanxianhuangdou 2.4170.189À92.2**845.151.7À93.9**292.022.0À92.5**a LAI was determined on three plots at55DAP.Total biomass and grain yield are expressed on a ground area basis.Measurements were made on three subplots per plot.Shoot biomass and grain were determined at ripening.*Significant differences between control and UV-B radiation at P<0:05according to t-test.**Significant differences between control and UV-B radiation at P<0:01according to t-test.Table3RI of20wheat cultivars under enhanced UV-B radiation,together with their origin and habitat aRank Cultivar name Origin Habitat RI1Yunnan97801South China High elevation,uplandÀ72.4 2Lanyin20North China Middle elevationÀ124.0 3Yunnan97929South China High elevation,uplandÀ128.5 4Yudou10South China Low elevation,lowlandÀ143.9 5Yudou18South China Low elevation,lowlandÀ162.4 6Yudou8South China Low elevation,lowlandÀ163.1 7Lvgundou North China Middle elevation,uplandÀ169.7 8Heidadou North China Middle elevation,uplandÀ172.3 9Yunnan97501South China High elevation,uplandÀ181.3 10Pitiaohuangdou North China Middle elevationÀ189.7 11Yunnan97506South China High elevation,uplandÀ194.2 12Yunnan97701South China High elevation,uplandÀ200.9 13Longdou1North China Middle elevation,uplandÀ215.1 14E0138South China High elevation,uplandÀ223.4 15Yunnan96510South China High elevation,uplandÀ224.3 16Df-1South China High elevation,uplandÀ256.9 17Lingtaihuangdou North China Middle elevation,uplandÀ263.7 18Kanglehuangdou North China Middle elevation,uplandÀ264.6 19Tuhuangdou1North China Middle elevation,uplandÀ290.8 20Huanxianhuangdou North China Middle elevation,uplandÀ295.7a Ranking1–20is in the order of increasing sensitivity to UV-B radiation.Tuhuangdou1,Kanglehuangdou,Lingtaihuangdou and Df-1.The6most tolerant cultivars were Yunnan 97801,Lanyin20,Yunnan97929,Yudou10,Yudou 18,Yudou8,with response indices higher than À163.1.4.DiscussionThis is thefirst report identifying intraspecific responses in crop growth and yield of20soybean cultivars to enhanced UV-B radiation underfield conditions.This is supported by earlierfindings of intraspecific responses in soybean(Teramura and Murali,1986;Sullivan and Teramura,1990;Teramura et al.,1990a;D’surney et al.,1993),maize and wheat (Biggs and Kossuth,1978;Li et al.,2000a,b),horse-bean(Bennett,1981),cucumber(Murali and Tera-mura,1986),rice(Teramura et al.,1991;Barnes et al., 1993;Dai et al.,1994).Reduction in plant height has often been used as an index to assess the degree of UV-B radiation sensi-tivity(Biggs and Kossuth,1978).In thisfield study, UV-B radiation had significant effects on the height of 20soybean cultivars at80DAP and ripening(Table1). Sensitivity in height of20soybean cultivars to enhanced UV-B radiation was greater at80DAP than at the ripening.The correlation between height at80 DAP and height at ripening was significant(r¼0:754, P<0:01).Height at ripening was used in the RI.In a greenhouse experiment,UV-B radiation decreased plant height of soybean(Teramura,1980).UV-B radiation significantly dwarfed soybean,primarily due to shorter internodes rather than smaller node number(Teramura,1980).A similar result was observed in wheat(Li et al.,1998).This could be due to photo-oxidative destruction of the phytohor-mone indole acetic acid followed by reduced cell wall extensibility as demonstrated in sunflower seedlings (Ros and Tevini,1995).The levels of ethylene,which promote radial growth and reduce elongation,are increased after irradiation with UV-B(Caldwell et al.,1995).However,the mechanism for UV-B radiation to increase plant height is still not clear. Although UV-B radiation had no significant effects on leaf area and total leaf number of soybean in an early study(Teramura,1980),in this study,LAI of20 soybean cultivars responded significantly to UV-B radiation(Table2).Under UV-B radiation,sensitivity in LAI was significant.So,LAI may be used as a parameter for selection of UV-B tolerance.UV-B radiation may directly affect cell division and some intrinsic growth characteristics(Beggs et al.,1985).It may be associated with many of the changes observed following UV-B exposure,such as the changes in leaf area I was contributed by area per leaf and leaf number per subplot.In this study,leaf number per subplot and area per leaf decreased under UV-B radiation,respectively.Similarly,UV-B radiation decreased LA of soybean(Sullivan and Teramura, 1990)and this was the reason why soybean LAI decreased.Total biomass is a good indictor of the effects of UV-B radiation on growth(Teramura,1983).Total biomass represents a long-term integration of all biochemical,physiological,and growth parameters. Therefore,even subtle UV-B induced effects on phy-siological processes could accumulate and result in significant effects on biomass.Other studies indicated UV-B radiation decreased total biomass of soybean (Teramura,1983;Teramura et al.,1990b;Sullivan and Teramura,1990).Decrease in total biomass of soy-bean in this study are similar to those found in both greenhouse(Teramura,1980,1983)andfield(Sullivan and Teramura,1990)studies.No greenhouse experi-ments have reported increases or no changes in bio-mass of soybean with enhanced UV-B radiation (Teramura,1983).The effect of UV-B radiation on total biomass may be dependent on the concentration of UV-absorbing compounds that attenuate incoming radiation and affectively limit damage to cellular components including the genetic material(D’surney et al.,1993).Early experiments on effects of UV-B radiation on crops were conducted in growth chambers and green-houses.Owing to space limitations,it is impractical to grow plants to reproductive maturity in such facilities. Thus,very little is known about UV-B effects on grain yield of soybean(Teramura,1983).No changes were reported in a greenhouse study(Teramura et al., 1990b)and afield study(Sullivan and Teramura, 1990),grain yield of soybean cultivars Essex and Williams decreased when combined over the6-year field study(Teramura et al.,1990a).Reduced grain yield has important economic implications;conse-quently,using grain yield as a parameter in a RI is6L.Yuan et al./Field Crops Research78(2002)1–8necessary.In addition,comparisons between green-house andfield observations revealed that UV-B sen-sitivity based on seed yield of soybean differs markedly,indicating thatfield validations are ulti-mately necessary for assessing the potential impacts of increased solar UV-B radiation(Teramura and Murali,1986).The RI was a good indicator to assess plant sensi-tivity to enhanced UV-B radiation(Dai et al.,1994).In previous studies,parameters used in response indices were different.Only in the soybeanfield study of Teramura and Murali(1986),was grain yield included as a parameter,and response sensitivity of soybean to UV-B radiation differed markedly when based on seed yield.Additionally,these parameters were only observed with isolated plants of cucumber and rice during short-term experiments in the greenhouse (Murali and Teramura,1986;Teramura et al.,1991; Barnes et al.,1993;Dai et al.,1994).Because plants respond differently to UV-B in different environments (Teramura and Murali,1986)and inherent genetic differences in different cultivars also contribute,varia-bility between experiments could also be due to differences in growth conditions,length of UV-B radiation,stage of growth and the ratio of incident PPF to UV-B radiation,all of which have been demon-strated to greatly modify UV-B responsiveness(Ter-amura et al.,1991).Besides,the responses of soybean crops to UV-B were more complicated than those of isolated plants(Teramura and Murali,1986;Sullivan and Teramura,1990;Teramura et al.,1990a,b; D’surney et al.,1993).Intraspecific differences of soybean to UV-B radia-tion were observed in the greenhouse and thefield (Teramura and Murali,1986;D’surney et al.,1993).In this study,the parameters of the RI included plant height,LAI,total biomass and grain yield.The corre-lations between LAI and total biomass(r¼0:500, P<0:05),total biomass and grain yield(r¼0:672, P<0:01)were significant,respectively.And the parameters were measured in crops underfield con-ditions.Then the results of this study might provide realistic assessments of intraspecific responses among 20soybean cultivars to enhanced UV-B radiation. As ambient UV-B radiation level is greater at lower latitudes than that at higher latitudes,it is generally assumed that crop cultivars originating near the equa-tor are more tolerant to UV-B radiation.In this study,out of the6tolerant cultivars,5cultivars originated from south China(low latitude).The most sensitive cultivars,i.e.Huanxianhuangdou(RI-295.7),Tuhuang-dou1(RI-290.8),Kanglehuangdou(RI-264.6)and Lingtaihuangdou(RI-263.7),originated in north China(high latitude).Similar results were observed in wheat in afield study(Li et al.,2000a).However, rice cultivars originating from regions with high ambi-ent UV-B were not necessarily more tolerant to enhanced UV-B radiation(Barnes et al.,1993;Dai et al.,1994).The UV-B tolerant cultivars identified in our study might be used as possible donors in breeding programs;however,the genetic basis for these differ-ences must be further examined.In conclusion,enhanced UV-B radiation had sig-nificant effect on plant height,LAI,total biomass and grain yield of soybean crops underfield conditions. The RI presented here showed that soybean is poten-tially a UV-B sensitive species.This result was similar to the previous study(Biggs and Kossuth,1978).UV-B sensitivity might be associated with UV-B radiation fluences and long-term accumulation(Li et al.,1998). Soybean is an economically important crop and its intraspecific response to UV-B radiation underfield conditions is still not clearly understood.Effects of UV-B radiation on plants are related to other environ-mental factors,including PPF,CO2,drought,phos-phorus nutrition,temperature,ozone fumigation and heavy metal(Teramura et al.,1990b;Caldwell et al., 1995).Furtherfield studies are needed to improve the understanding of the relationship between UV-B radiation effectiveness and other environmental vari-ables.This would greatly enhance our ability to more realistically assess intraspecific responses of soybean to increased levels of UV-B radiation. AcknowledgementsThis work was supported by the National Natural Science Foundation of China,the National Research Foundation for Post-Doctorate Studies in China, the Applied Base Research Foundation of Yunnan Province,China,and the Research Foundation for Academic Leaders in Yunnan Province,China.We wish to thank Tan Lingling of the Department of Biology,Lanzhou University,for supporting the experimental work.L.Yuan et al./Field Crops Research78(2002)1–87ReferencesBarnes,P.W.,Maggard,S.,Holman,S.R.,Vergara, B.,1993.Intraspecific variation in sensitivity to UV-B radiation in rice.Crop Sci.33,1041–1046.Beggs, C.J.,Schneider-Zeibert,R.,Wellman, E.,1985.UV-B radiation and adaptive mechanisms in plants.In:Worrest,R.C.(Ed.),Stratospheric Ozone Reduction,Solar Ultraviolet Radia-tion and Plant Life.Springer,Berlin,pp.235–250. Bennett,J.H.,1981.Photosynthesis and gas diffusion in leaves of selected crop plants exposed to ultraviolet-B radiation.J.Environ.Qual.10,271–275.Biggs,R.H.,Kossuth,S.V.,1978.Impact of solar UV-B radiation on crop productivity.Final report of UV-B biological and climate effects research.Terr.FY77,11–77.Caldwell,M.M.,1971.Solar UV-B irradiation and the growth and development of higher plants.In:Giese,A.C.(Ed.),Photo-physiology,V ol.6.Academic Press,New York,NY,pp.131–171. Caldwell,M.M.,Flint,S.D.,1994.Stratospheric ozone reduction, solar UV-B radiation and terrestrial ecosystem.Climate Change 27,375–394.Caldwell,M.M.,Teramura,A.H.,Tevini,M.,Kulandaivelu,G., 1995.Effects of increased solar ultraviolet radiation on terrestrial plants.Ambio24,166–173.Caldwell,M.M.,Bjorn,L.O.,Bornman,J.F.et al.,1998.Effects of increased solar ultraviolet radiation on terrestrial ecosystem.J.Photochem.Photobiol.Biol.46,40–52.D’surney,S.J.,Tschaplinski,T.J.,Edwards,N.T.,Shugart,L.R., 1993.Biological responses of two soybean cultivars exposed to enhanced UV-B radiation.Environ.Exp.Bot.33,347–356. Dai,Q.J.,Peng,S.B.,Chavez, A.Q.,Vergara, B.S.,1994.Intraspecific responses of188rice cultivars to enhanced UV-B radiation.Environ.Exp.Bot.34,422–433.Kerr,R.A.,1993.The ozone hole reaches a new low.Science262, 501.Li,Y.,Yue,M.,Wang,X.L.,1998.Effects of enhanced ultraviolet-B radiation on crop structure,growth and yield components ofspring wheat underfield conditions.Field Crops Res.57,253–263.Li,Y.,Yue,M.,Wang,X.L.,Hu,Z.D.,petition and sensitivity of wheat and wild oat exposed to enhanced UV-B radiation at different densities underfield conditions.Environ.Exp.Bot.41,47–55.Li,Y.,Zu,Y.Q.,Chen,H.Y.,Chen,J.J.,Yang,J.L.,Hu,Z.D., 2000a.Intraspecific responses in crop growth and yield of20 wheat cultivars to enhanced ultraviolet-B radiation underfield conditions.Field Crops Res.67,25–33.Li,Y.,Zu,Y.Q.,Chen,J.J.,Chen,H.Y.,Yang,J.L.,Hu,Z.D., 2000b.Intraspecific differences in physiological responses of 20wheat cultivars to enhanced ultraviolet-B radiation under field conditions.Environ.Exp.Bot.44(2),95–103. Lydon,J.,Teramure, A.H.,Summers, E.G.,1986.Effects of ultraviolet-B radiation on growth and productivity offield-grown soybean.In:Worrest,R.C.(Ed.),Stratospheric Ozone Reduction,Solar Ultraviolet Radiation and Plant Life.Springer, Berlin,pp.313–325.Madronich,S.,Mckenzie,R.L.,Caldwell,M.M.,Bjorn,L.O.,1995.Changes in ultraviolet radiation reaching the earth’s surface.Ambio24,143–153.Molina,M.J.,Molina,L.T.,1992.Stratospheric ozone.ACS Symp.Ser.483,24–35.Murali,N.S.,Teramura,A.H.,1986.Intraspecific differences in Cucumis sativus sensitivity to ultraviolet-B radiation.Physiol.Plant.68,673–677.Ros,J.,Tevini,M.,1995.Interaction of UV-radiation and IAA during growth of seedling and hypocotyl segments of sun-flower.J.Plant Physiol.146,295–302.Sullivan,J.H.,Teramura,A.H.,1990.Field study of the interaction between solar ultraviolet-B radiation and drought on photo-synthesis and growth in soybean.Plant Physiol.92,141–146. Teramura, A.H.,1980.Effects of ultraviolet-B irradiances on soybean.I.Importance of photosynthetically active radiation in evaluating ultraviolet-B irradiance effects on soybean and wheat growth.Physiol.Plant.48,333–339.Teramura,A.H.,1983.Effects of ultraviolet-B radiation on the growth and yield of crop plants.Physiol.Plant.58,415–427. Teramura,A.H.,Murali,N.S.,1986.Intraspecific differences in growth and yield of soybean exposed to ultraviolet-B radiation under greenhouse andfield conditions.Environ.Exp.Bot.26, 89–95.Teramura,A.H.,Sullivan,J.H.,Lydon,J.,1990a.Effects of UV-B radiation on soybean yield and seed quality:a6-yearfield study.Physiol.Plant.80,5–11.Teramura,A.H.,Sullivan,J.H.,Ziska,L.H.,1990b.Interaction of elevated ultraviolet-B radiation and CO2on productivity and photosynthetic characteristics in wheat.Plant Physiol.94,470–475.Teramura, A.H.,Ziska,L.H.,Sztein, A.E.,1991.Changes in growth and photosynthetic capacity of rice with increased UV-B radiation.Physiol.Plant.83,373–380.Yue,M.,Li,Y.,Wang,X.L.,1988.Effects of enhanced ultraviolet-B radiation on plant nutrients and decomposition of spring wheat underfield conditions.Environ.Exp.Bot.40, 187–196.8L.Yuan et al./Field Crops Research78(2002)1–8。